
Protein Changes in Shade and Sun Haberlea rhodopensis Leaves during Dehydration at Optimal and Low Temperatures
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
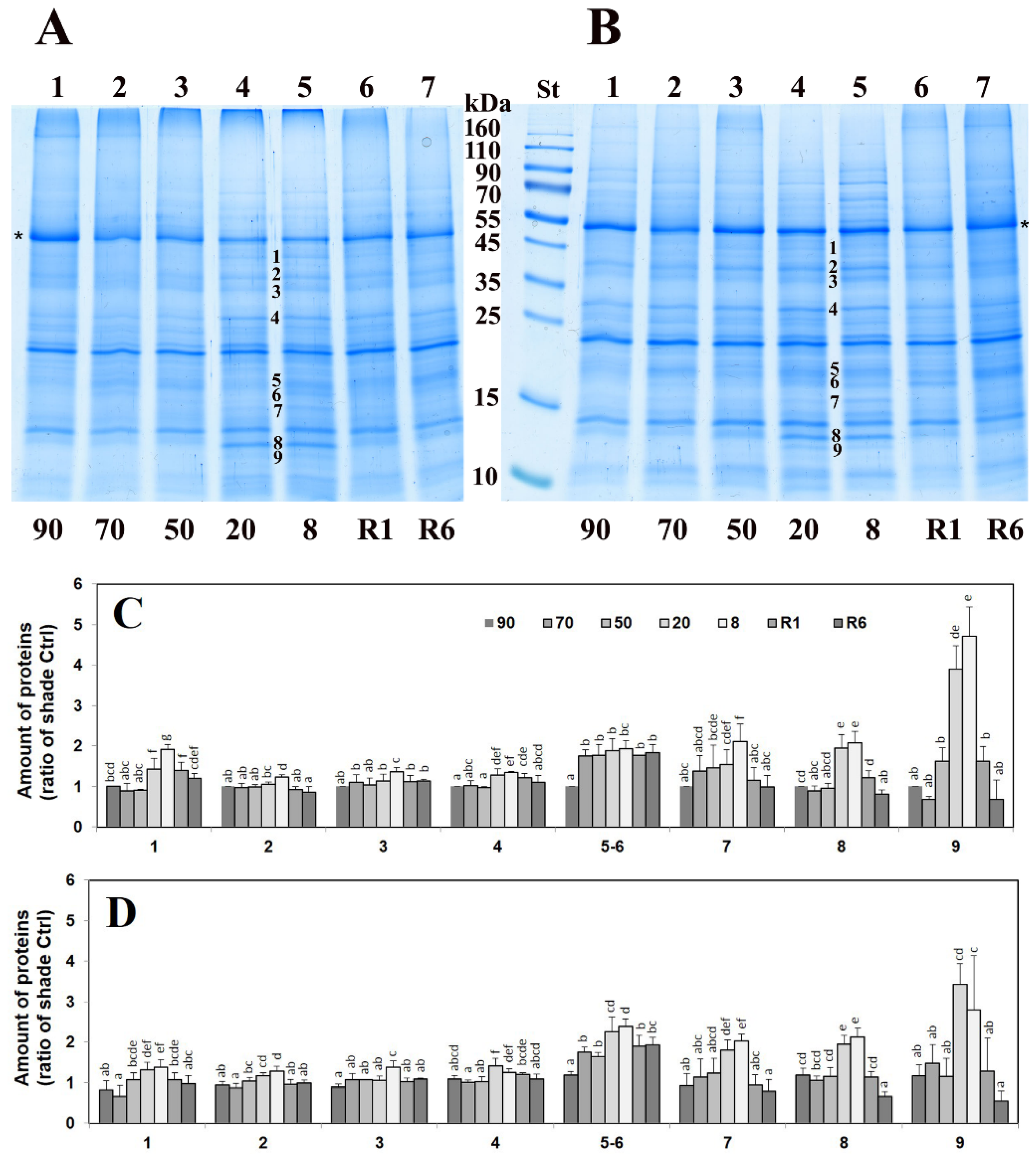
2.1. Stress-Induced Changes in the Leaf Proteome
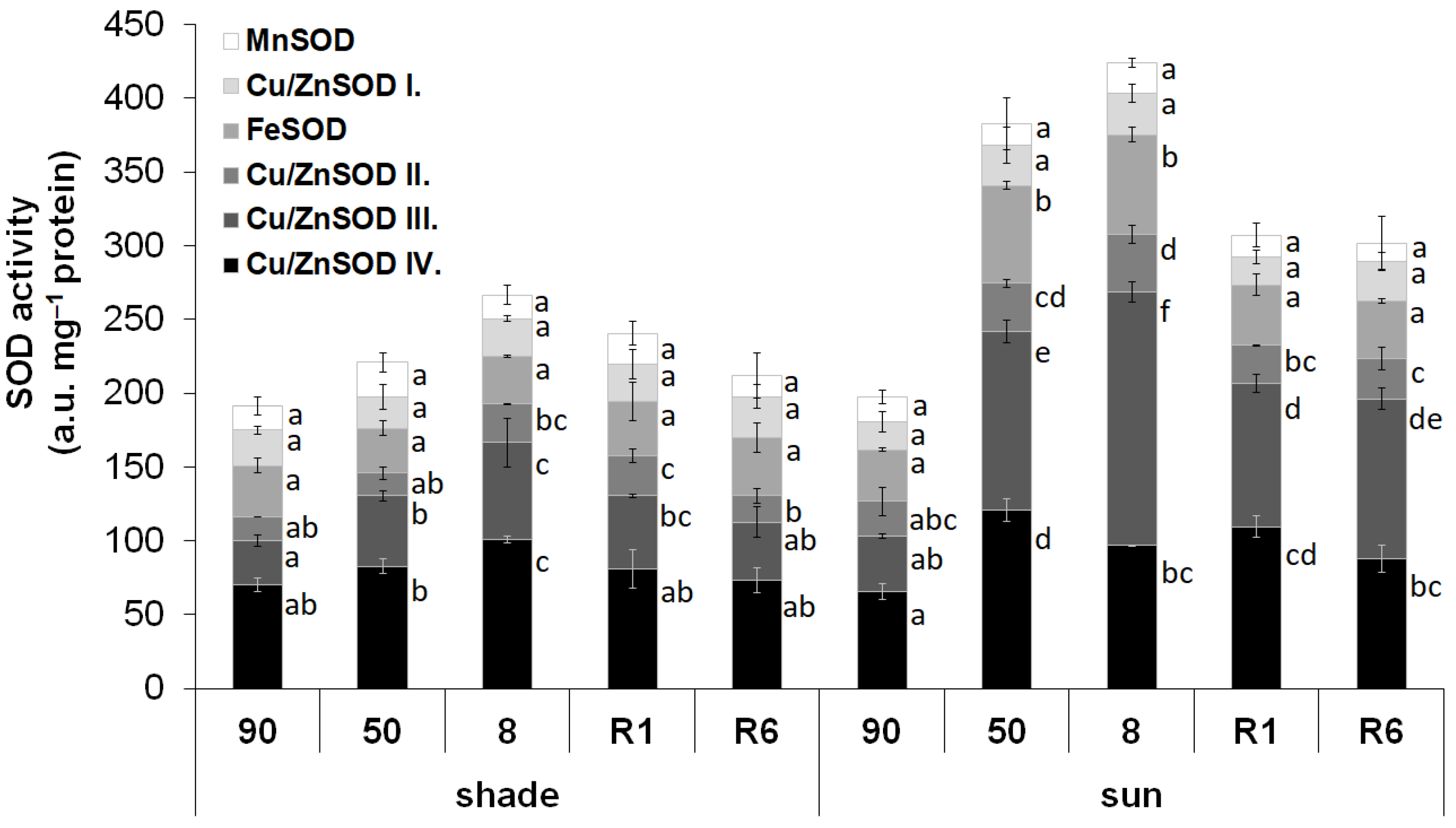
2.2. Activity of SOD Isoenzymes
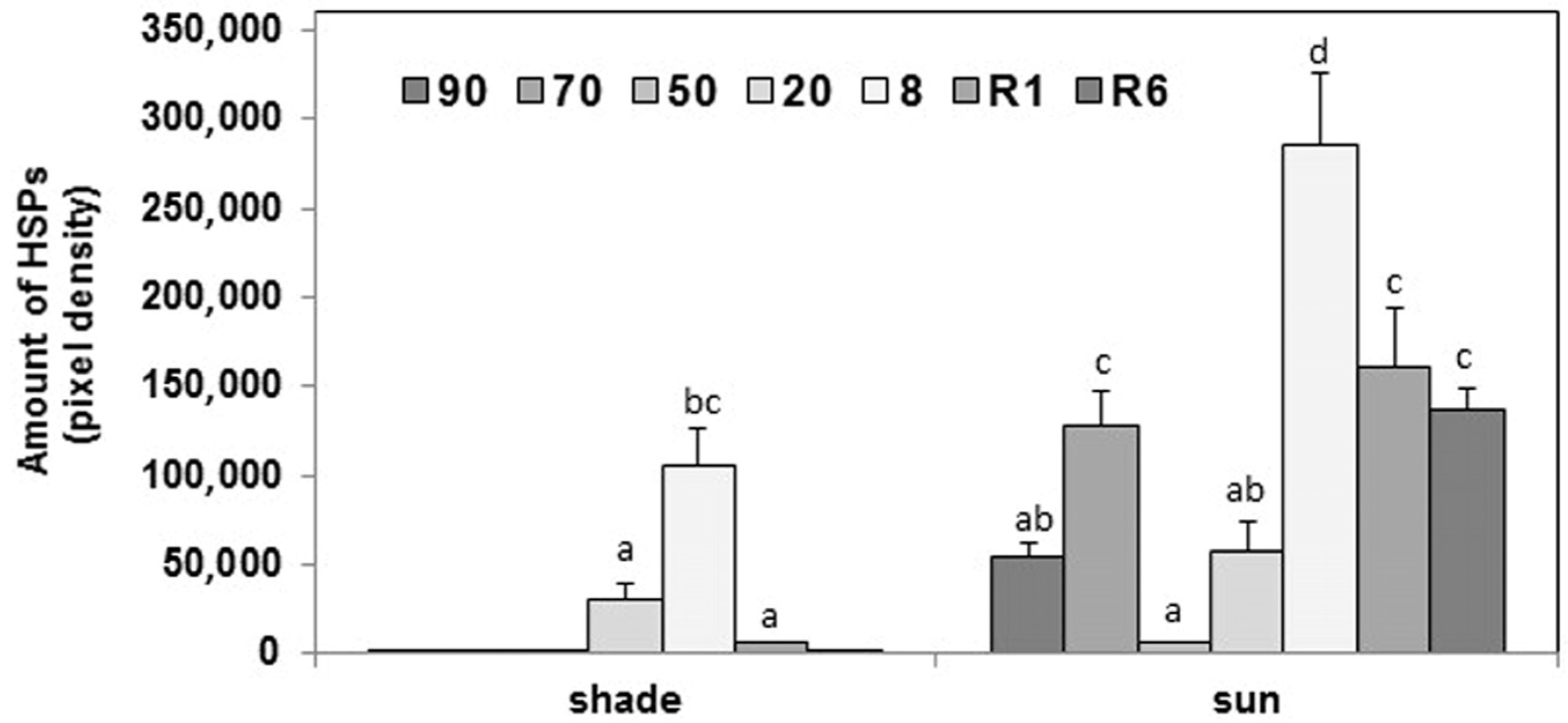
2.3. Protective Proteins
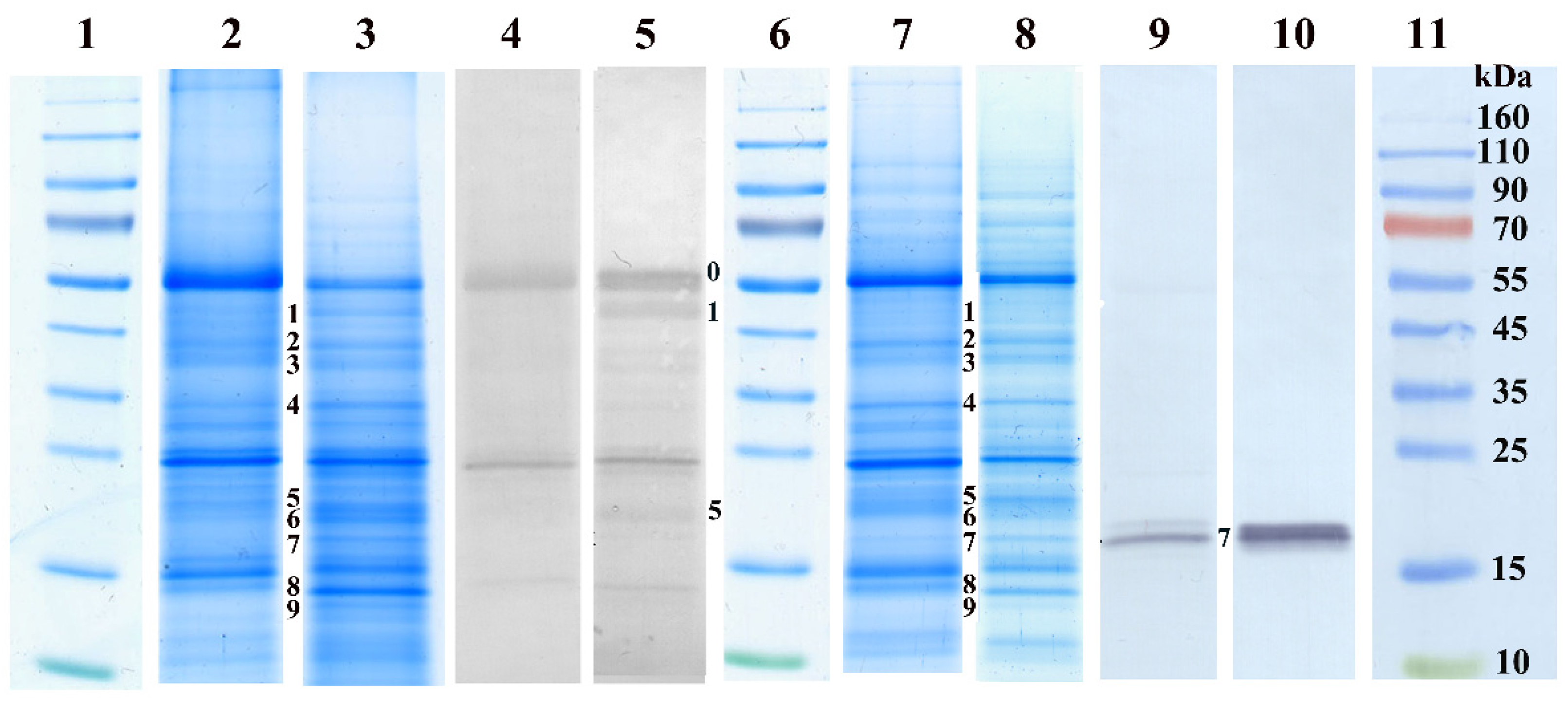
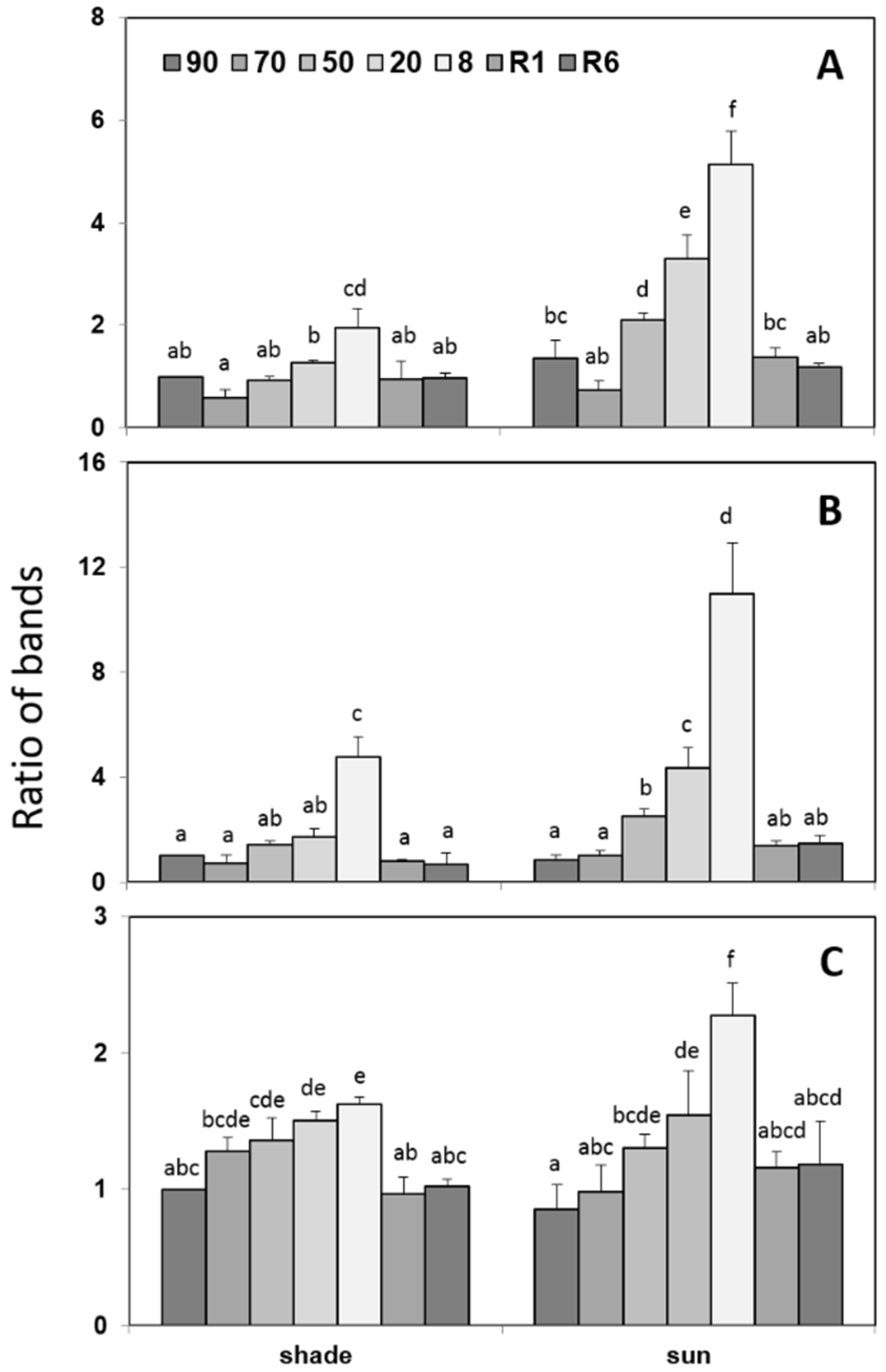
2.4. Untargeted Detection of Changes in the Proteome
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Determination of Relative Water Content (RWC)
4.3. Extraction and SDS PAGE Separation of Leaf Proteins
4.4. Protein Blotting and Western Blot Analysis
4.5. Activity Measurement of Superoxide Dismutase (SOD) Isoforms
4.6. Protein Identification by Mass Spectrometry
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farrant, J.M.; Brandt, W.; Lindsey, G. An overview of mechanisms of desiccation tolerance in selected angiosperm resurrection plants. Plant Stress 2007, 1, 72–84. [Google Scholar] [CrossRef]
- Rakić, T.; Lazarević, M.; Jovanović, Ž.S.; Radović, S.; Siljak-Yakovlev, S.; Stevanović, B.; Stevanović, V. Resurrection plants of the genus Ramonda: Prospective survival strategies-unlock further capacity of adaptation, or embark on the path of evolution? Front. Plant Sci. 2014, 4, 550. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, K.; Dagnon, S.; Gesheva, E.; Bojilov, D.; Mihailova, G.; Doncheva, S. Antioxidant defense during desiccation of the resurrection plant Haberlea rhodopensis. Plant Physiol. Biochem. 2017, 114, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Gashi, B.; Babani, F.; Kongjika, E. Chlorophyll fluorescence imaging of photosynthetic activity and pigment contents of the resurrection plants Ramonda serbica and Ramonda nathaliae during dehydration and rehydration. Physiol. Mol. Biol. Plants 2013, 19, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Marín, B.; Nadal, M.; Gago, J.; Fernie, A.R.; López-Pozo, M.; Artetxe, U.; García-Plazaola, J.I.; Verhoeven, A. Born to revive: Molecular and physiological mechanisms of double tolerance in a paleotropical and resurrection plant. New Phytol. 2020, 226, 741–759. [Google Scholar] [CrossRef]
- Petrova, G.; Moyankova, D.; Nishii, K.; Forrest, L.; Tsiripidis, I.; Drouzas, A.D.; Djilianov, D.; Möller, M. The European paleoendemic Haberlea rhodopensis (Gesneriaceae) has an Oligocene origin and a Pleistocene diversification and occurs in a long-persisting refugial area in Southeastern Europe. Int. J. Plant Sci. 2015, 176, 499–514. [Google Scholar] [CrossRef]
- Tuba, Z.; Protor, C.F.; Csintalan, Z. Ecophysiological responses of homoiochlorophyllous and poikilochlorophyllous desiccation tolerant plants: A comparison and an ecological perspective. Plant Growth Regul. 1998, 24, 211–217. [Google Scholar] [CrossRef]
- Daskalova, E.; Dontcheva, S.; Yahubyan, G.; Minkov, I.; Toneva, V. Ecological characteristics and conservation of the protected resurrection species Haberlea rhodopensis Friv. as in vitro plants through a modified micropropagation system. Biotechnol. Biotechnol. Equip. 2010, 24, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Sárvári, É.; Mihailova, G.; Solti, Á.; Keresztes, Á.; Velitchkova, M.; Georgieva, K. Comparison of thylakoid structure and organization in sun and shade Haberlea rhodopensis populations under desiccation and rehydration. J. Plant Physiol. 2014, 171, 1591–1600. [Google Scholar] [CrossRef]
- Rapparini, F.; Neri, L.; Mihailova, G.; Petkova, S.; Georgieva, K. Growth irradiance affects the photoprotective mechanisms of the resurrection angiosperm Haberlea rhodopensis Friv. in response to desiccation and rehydration at morphological, physiological and biochemical levels. Environ. Exp. Bot. 2015, 113, 67–69. [Google Scholar] [CrossRef]
- Mihailova, G.; Solti, Á.; Sárvári, É.; Keresztes, Á.; Francesca Rapparini, F.; Velitchkova, M.; Simova-Stoilova, L.; Aleksandrov, V.; Georgieva, K. Freezing tolerance of photosynthetic apparatus in the homoiochlorophyllous resurrection plant Haberlea rhodopensis. Environ. Exp. Bot. 2020, 178, 104157. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solti, Á.; Lenk, S.; Mihailova, G.; Mayer, P.; Barócsi, A.; Georgieva, K. Effects of habitat light conditions on the excitation quenching pathways in desiccating Haberlea rhodopensis leaves: An Intelligent FluoroSensor study. J. Photochem. Photobiol. B Biol. 2014, 130, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 252, 275–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyall, R.; Gechev, T. Multi-omics insights into the evolution of angiosperm resurrection plants. Annu. Plant Rev. Online 2020, 3, 77–110. [Google Scholar] [CrossRef]
- Hoekstra, F.A.; Golovina, E.A.; Buitink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef]
- Close, T.J. Dehydrins: A commonalty in the response of plants to dehydration and low temperature. Physiol. Plant. 1997, 100, 291–296. [Google Scholar] [CrossRef]
- Basha, E.; O’Neill, H.; Vierling, E. Small heat shock proteins and α-crystallins: Dynamic proteins with flexible functions. Trends Biochem. Sci. 2012, 37, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Close, T.J. Dehydrins: Emergence of a biochemical role of a family of plant dehydration proteins. Physiol. Plant. 1996, 97, 795–803. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, X.; Zhang, L. Structural and functional dynamics of dehydrins: A plant protector protein under abiotic stress. Int. J. Mol. Sci. 2018, 19, 3420. [Google Scholar] [CrossRef]
- Graether, S.P.; Boddington, K.F. Disorder and function: A review of the dehydrin protein family. Front. Plant Sci. 2014, 5, 576. [Google Scholar] [CrossRef] [Green Version]
- Giarola, V.; Challabathula, D.; Bartels, D. Quantification of expression of dehydrin isoforms in the desiccation tolerant plant Craterostigma plantagineum using specifically designed reference genes. Plant Sci. 2015, 236, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.R.; Vierling, E. Plant small heat shock proteins–evolutionary and functional diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieva, K.; Szigeti, Z.; Sarvari, E.; Gaspar, L.; Maslenkova, L.; Peeva, V.; Peli, E.; Tuba, Z. Photosynthetic activity of homoiochlorophyllous desiccation tolerant plant Haberlea rhodopensis during dehydration and rehydration. Planta 2007, 225, 955–964. [Google Scholar] [CrossRef]
- Georgieva, K.; Sárvári, E.; Keresztes, A. Protection of thylakoids against combined light and drought by a lumenal substance in the resurrection plant Haberlea rhodopensis. Ann. Bot. 2010, 105, 117–126. [Google Scholar] [CrossRef]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Pérez-Pérez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox regulation of the Calvin–Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef] [Green Version]
- Gurrieri, L.; Fermani, S.; Zaffagnini, M.; Sparla, F.; Trost, P. Calvin–Benson cycle regulation is getting complex. Trends Plant Sci. 2021, 26, 898–912. [Google Scholar] [CrossRef]
- Xu, X.; Legay, S.; Sergeant, K.; Zorzan, S.; Leclercq, C.C.; Charton, S.; Giarola, V.; Liu, X.; Challabathula, D.; Renaut, J.; et al. Molecular insights into plant desiccation tolerance: Transcriptomics, proteomics and targeted metabolite profiling in Craterostigma plantagineum. Plant J. 2021, 107, 377–398. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, K.; Rapparini, F.; Bertazza, G.; Mihailova, G.; Sárvári, É.; Solti, Á.; Keresztes, Á. Alterations in the sugar metabolism and in the vacuolar system of mesophyll cells contribute to the desiccation tolerance of Haberlea rhodopensis ecotypes. Protoplasma 2017, 254, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Mihailova, G.; Abakumov, D.; Büchel, C.; Dietzel, L.; Georgieva, K. Drought-responsive gene expression in sun and shade plants of Haberlea rhodopensis under controlled environment. Plant Mol. Biol. Rep. 2017, 35, 313–322. [Google Scholar] [CrossRef]
- Mihailova, G.; Büchel, C.; Dietzel, L.; Georgieva, K. Desiccation induced changes in photosynthesis related proteins of shade and sun Haberlea rhodopensis plants. C. R. Acad. Bulg. Sci. 2016, 69, 37–44. [Google Scholar]
- Yahubyan, G.; Gozmanova, M.; Denev, I.; Toneva, V.; Minkov, I. Prompt response of superoxide dismutase and peroxidase to dehydration and rehydration of the resurrection plant Haberlea rhodopensis. Plant Growth Regul. 2009, 57, 49–56. [Google Scholar] [CrossRef]
- Tebele, S.M.; Marks, R.A.; Farrant, J.M. Two decades of desiccation biology: A systematic review of the best studied angiosperm resurrection plants. Plants 2021, 10, 2784. [Google Scholar] [CrossRef] [PubMed]
- Morse, M.; Rafudeen, M.; Farrant, J.M. An overview of the current understanding of desiccation tolerance in the vegetative tissues of higher plants. Adv. Bot. Res. 2011, 57, 319–347. [Google Scholar] [CrossRef]
- Mladenov, P.; Zasheva, D.; Planchon, S.; Leclercq, C.C.; Falconet, D.; Moyet, L.; Brugière, S.; Moyankova, D.; Tchorbadjieva, M.; Ferro, M.; et al. Proteomics evidence of a systemic response to desiccation in the resurrection plant Haberlea rhodopensis. Int. J. Mol. Sci. 2022, 23, 8520. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Benina, M.; Obata, T.; Tohge, T.; Sujeeth, N.; Minkov, I.; Hille, J.; Temanni, M.-R.; Marriott, A.S.; Bergström, E.; et al. Molecular mechanisms of desiccation tolerance in the resurrection glacial relic Haberlea rhodopensis. Cell Mol. Life Sci. 2013, 70, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Ly, L.; Wasinger, V.C. Protein and peptide fractionation, enrichment and depletion: Tools for the complex proteome. Proteomics 2011, 11, 513–534. [Google Scholar] [CrossRef]
- Herman, E.M.; Li, X.; Su, R.T.; Larsen, P.; Hsu, H.T.; Sze, H. Vacuolar-type H+-ATPases are associated with the endoplasmic reticulum and provacuoles of root tip cells. Plant Physiol. 1994, 106, 1313–1324. [Google Scholar] [CrossRef]
- Martinoia, E.; Maeshima, M.; Neuhaus, H.E. Vacuolar transporters and their essential role in plant metabolism. J. Exp. Bot. 2007, 58, 83–102. [Google Scholar] [CrossRef]
- Mundree, S.G.; Farrant, J.M. Some physiological and molecular insights into the mechanisms of desiccation tolerance in the resurrection plant Xerophyta viscosa Baker. In Plant Tolerance to Abiotic Stresses in Agriculture: Role of Genetic Engineering; Cherry, J.H., Locy, R.D., Rychter, A., Eds.; NATO Science Series; Springer: Dordrecht, The Netherlands, 2000; Volume 83, pp. 201–222. [Google Scholar]
- Zhang, H.; Ni, Z.; Chen, Q.; Guo, Z.; Gao, W.; Su, X.; Qu, Y. Proteomic responses of drought-tolerant and drought-sensitive cotton varieties to drought stress. Mol. Genet. Genom. 2016, 291, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Ni, Z.; Zhang, H.; Chen, Q.; Gao, W.; Cai, Y.; Li, M.; Sun, G.; Qu, Y.Y. The gene encoding subunit A of the vacuolar H+-ATPase from cotton plays an important role in conferring tolerance to water deficit. Front. Plant Sci. 2018, 9, 758. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.G.; Wang, S.H.; Luo, H.; Ma, Q.J.; Yao, Y.X.; You, C.X.; Hao, Y.J. Overexpression of MdVHA-B, a V-ATPase gene from apple, confers tolerance to drought in transgenic tomato. Sci. Hortic. 2012, 145, 94–101. [Google Scholar] [CrossRef]
- Vasanthakumar, T.; Rubinstein, J.L. Structure and Roles of V-type ATPases. Trends Biochem. Sci. 2020, 45, 295–307. [Google Scholar] [CrossRef]
- Wang, C.; Xiang, Y.; Qian, D. Current progress in plant V-ATPase: From biochemical properties to physiological functions. J. Plant Physiol. 2021, 266, 153525. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Martinoia, E.; Meyer, S.; De Angeli, A.; Nagy, R. Vacuolar transporters in their physiological context. Annu. Rev. Plant Biol. 2012, 63, 183–213. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Sprenger, N.; Bortlik, K.; Boller, T.; Wiemken, A. Desiccation increases sucrose levels in Ramonda and Haberlea, two genera of resurrection plants in the Gesneriaceae. Physiol. Plant. 1997, 100, 153–158. [Google Scholar] [CrossRef]
- Moyankova, D.; Mladenov, P.; Berkov, S.; Peshev, D.; Georgieva, D.; Djilianov, D. Metabolic profiling of the resurrection plant Haberlea rhodopensis during desiccation and recovery. Physiol. Plant. 2014, 152, 675–687. [Google Scholar] [CrossRef]
- Koo, K.M.; Jung, S.; Lee, B.S.; Kim, J.B.; Jo, Y.D.; Choi, H.I.; Kang, S.Y.; Chung, G.H.; Jeong, W.J.; Ahn, J.W. The mechanism of starch over-accumulation in Chlamydomonas reinhardtii high-starch mutants identified by comparative transcriptome analysis. Front. Microbiol. 2017, 8, 858. [Google Scholar] [CrossRef] [PubMed]
- Sheshukova, E.V.; Komarova, T.V.; Pozdyshev, D.V.; Ershova, N.M.; Shindyapina, A.V.; Tashlitsky, V.N.; Sheval, E.V.; Dorokhov, Y.L. The intergenic interplay between aldose 1-epimerase-like protein and pectin methylesterase in abiotic and biotic stress control. Front. Plant Sci. 2017, 8, 1646. [Google Scholar] [CrossRef] [Green Version]
- Van Der Straeten, D.; Rodrigues-Pousada, R.A.; Goodman, H.M.; Van Montagu, M. Plant enolase: Gene structure, expression, and evolution. Plant Cell 1991, 3, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Dong, K.; Ge, P.; Bian, Y.; Dong, L.; Deng, X.; Li, X.; Yan, Y. Identification of leaf proteins differentially accumulated between wheat cultivars distinct in their levels of drought tolerance. PLoS ONE 2015, 10, e0125302. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Guo, Y.; Ohta, M.; Xiong, L.; Stevenson, B.; Zhu, J.K. LOS2, a genetic locus required for cold-responsive gene transcription encodes a bi-functional enolase. EMBO J. 2002, 21, 2692–2702. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, K.M.; Weaver, L.M. The shikimate pathway. Annu. Rev. Plant Biol. 1999, 50, 473. [Google Scholar] [CrossRef]
- Doltchinkova, V.; Andreeva, T.; Georgieva, K.; Mihailova, G.; Balashev, K. Desiccation-induced alterations in surface topography of thylakoids from resurrection plant Haberlea rhodopensis studied by atomic force microscopy, electrokinetic and optical measurements. Physiol. Plant. 2019, 166, 585–595. [Google Scholar] [CrossRef]
- Smith, J.; Yang, Y.; Levy, S.; Adelusi, O.O.; Hahn, M.G.; O’Neill, M.A.; Bar-Peled, M. Functional characterization of UDP-apiose synthases from bryophytes and green algae provides insight into the appearance of apiose-containing glycans during plant evolution. J. Biol. Chem. 2016, 291, 21434–21447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, S.R. Caffeoyl phenylethanoid glycosides in Sanango racemosum and in the Gesneriaceae. Phytochemistry 1996, 43, 777–783. [Google Scholar] [CrossRef]
- Fu, X.; Chen, J.; Xie, R.; Zhou, L.; Wei, Y.; Yuan, C.; Huang, L.; Hu, Z.; Hao, X.; Gu, W. Phytochemical and chemotaxonomic studies on Paraboea rufescens (Gesneriaceae). Biochem. Syst. Ecol. 2022, 102, 104414. [Google Scholar] [CrossRef]
- Xu, H.; Huang, C.; Jiang, X.; Zhu, J.; Gao, X.; Yu, C. Impact of cold stress on leaf structure, photosynthesis, and metabolites in Camellia weiningensis and C. oleifera seedlings. Horticulturae 2022, 8, 494. [Google Scholar] [CrossRef]
- Liu, C.J.; Huhman, D.; Sumner, L.W.; Dixon, R.A. Regiospecific hydroxylation of isoflavones by cytochrome p450 81E enzymes from Medicago truncatula. Plant J. 2003, 36, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Rani, D.; Kobtrakul, K.; Luckanagul, J.A.; Thaweesest, W.; Rojsitthisak, P.; De-Eknamkul, W.; Vimolmangkang, S. Differential gene expression levels, chemical profiles, and biological activities of Pueraria candollei var. mirifica callus cultures at different growth stages. Plant Cell Tissue Organ Cult. 2021, 147, 61–72. [Google Scholar] [CrossRef]
- Georgiev, Y.N.; Ognyanov, M.H.; Denev, P.N. The ancient Thracian endemic plant Haberlea rhodopensis Friv. and related species: A review. J. Ethnopharmacol. 2020, 249, 112359. [Google Scholar] [CrossRef]
- Spyridopoulou, K.; Kyriakou, S.; Nomikou, A.; Roupas, A.; Ermogenous, A.; Karamanoli, K.; Moyankova, D.; Djilianov, D.; Galanis, A.; Panayiotidis, M.I.; et al. Chemical profiling, antiproliferative and antimigratory capacity of Haberlea rhodopensis extracts in an in vitro platform of various human cancer cell lines. Antioxidants 2022, 11, 2305. [Google Scholar] [CrossRef]
- Mulako, I.; Farrant, J.M.; Collett, H.; Illing, N. Expression of Xhdsi-1VOC, a novel member of the vicinal oxygen chelate (VOC) metalloenzyme superfamily, is up-regulated in leaves and roots during desiccation in the resurrection plant Xerophyta humilis (Bak) Dur and Schinz. J. Exp. Bot. 2008, 59, 3885–3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, L.; Zhang, C.Y.; Wang, X.D.; Wang, H.; Long, Y.; Yin, Y.T.; Li, D.R.; Tian, J.H.; Li, Z.Y.; Lin, Z.W.; et al. Proteomic and comparative genomic analysis of two Brassica napus lines differing in oil content. J. Proteome Res. 2013, 12, 4965–4978. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Wan, N.; Cheng, Z.; Mo, Y.; Liu, B.; Liu, H.; Raboanatahiry, N.; Yin, Y.; Li, M. Whole-genome identification and expression pattern of the vicinal oxygen chelate family in rapeseed (Brassica napus L.). Front. Plant Sci. 2017, 8, 745. [Google Scholar] [CrossRef] [Green Version]
- Hoque, T.S.; Hossain, M.A.; Mostofa, M.G.; Burritt, D.J.; Fujita, M.; Tran, L.S.P. Methylglyoxal: An emerging signaling molecule in plant abiotic stress responses and tolerance. Front. Plant Sci. 2016, 7, 1341. [Google Scholar] [CrossRef] [Green Version]
- Mostofa, M.G.; Ghosh, A.; Li, Z.G.; Siddiqui, M.N.; Fujita, M.; Tran, L.S.P. Methylglyoxal—A signaling molecule in plant abiotic stress responses. Free Radic. Biol. Med. 2018, 122, 96–109. [Google Scholar] [CrossRef]
- Georgieva, K.; Doncheva, S.; Mihailova, G.; Petkova, S. Response of sun- and shade- adapted plants of Haberlea rhodopensis to desiccation. Plant Growth Regul. 2012, 67, 121–132. [Google Scholar] [CrossRef]
- Solti, Á.; Mihailova, G.; Sárvári, É.; Georgieva, K. Antioxidative defence mechanism contributes to desiccation tolerance in Haberlea rhodopensis population naturally exposed to high irradiation. Acta Biol. Szeged. 2014, 58, 11–14. [Google Scholar]
- Bowler, C.; Van Camp, W.; Van Montagu, M.; Inzé, D.; Asada, K. Superoxide dismutase in plants. Critical Rev.Plant Sci. 1994, 13, 199–218. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Skotnicová, P.; Staleva-Musto, H.; Kuznetsova, V.; Bína, D.; Konert, M.M.; Lu, S.; Polívka, T.; Sobotka, R. Plant LHC-like proteins show robust folding and static non-photochemical quenching. Nat. Commun. 2021, 12, 6890. [Google Scholar] [CrossRef] [PubMed]
- Alamillo, J.M.; Bartels, D. Effects of desiccation on photosynthesis pigments and the ELIP-like dsp22 protein complex in the resurrection plant Craterostigma plantagineum. Plant Sci. 2001, 160, 1161–1170. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Yang, G.; Zhang, L.; Yang, X.; Zhao, S.; Ji, Z.; Zhou, Q.; Hu, M.; Wang, Y.; Chen, M.; et al. The resurrection genome of Boea hygrometrica: A blueprint for survival of dehydration. Proc. Natl. Acad. Sci. USA 2015, 112, 5833–5837. [Google Scholar] [CrossRef] [Green Version]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Van Montagu, M.; Verbruggen, N. Small heat shock proteins and stress tolerance in plants. Biochim. Biophys. Acta Gene Struct. Expr. 2002, 1577, 1–9. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, M.; Li, X.; Cao, B.; Ma, X. Identification of differentially expressed genes in leaf of Reaumuria soongorica under PEG-induced drought stress by digital gene expression profiling. PLoS ONE 2014, 9, e104142. [Google Scholar] [CrossRef]
- Waters, E.R. The evolution, function, structure, and expression of the plant sHSPs. J. Exp. Bot. 2013, 64, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Sun, C.; Li, Z.; Hu, Q.; Han, L.; Luo, H. AsHSP17, a creeping bentgrass small heat shock protein modulates plant photosynthesis and ABA-dependent and independent signalling to attenuate plant response to abiotic stress. Plant Cell Environ. 2016, 39, 1320–1337. [Google Scholar] [CrossRef] [Green Version]
- Alamillo, J.; Almoguera, C.; Bartels, D.; Jordano, J. Constitutive expression of small heat shock proteins in vegetative tissues of the resurrection plant Craterostigma plantagineum. Plant Mol. Biol. 1995, 29, 1093–1099. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, B.; Sun, D.; Deng, X. Molecular cloning and differential expression of sHSP gene family members from the resurrection plant Boea hygrometrica in response to abiotic stresses. Biologia 2013, 68, 651–661. [Google Scholar] [CrossRef]
- Liu, J.; Moyankova, D.; Lin, C.T.; Mladenov, P.; Sun, R.Z.; Djilianov, D.; Deng, X. Transcriptome reprogramming during severe dehydration contributes to physiological and metabolic changes in the resurrection plant Haberlea rhodopensis. BMC Plant Biol. 2018, 18, 351. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, Ž.; Rakić, T.; Stevanović, B.; Radović, S. Characterization of oxidative and antioxidative events during dehydration and rehydration of resurrection plant Ramonda nathaliae. Plant Growth Regul. 2011, 64, 231–240. [Google Scholar] [CrossRef]
- Mihailova, G.; Christov, N.K.; Sárvári, É.; Solti, Á.; Hembrom, R.; Solymosi, K.; Keresztes, Á.; Velitchkova, M.; Popova, A.V.; Simova-Stoilova, L.; et al. Reactivation of the photosynthetic apparatus of resurrection plant Haberlea rhodopensis during the early phase of recovery from drought- and freezing-induced desiccation. Plants 2022, 11, 2185. [Google Scholar] [CrossRef] [PubMed]
- Collett, H.; Shen, A.; Gardner, M.; Farrant, J.M.; Denby, K.J.; Illing, N. Towards transcript profiling of desiccation tolerance in Xerophyta humilis: Construction of a normalized 11 k X. humilis cDNA set and microarray expression analysis of 424 cDNAs in response to dehydration. Physiol. Plant. 2004, 122, 39–53. [Google Scholar] [CrossRef]
- Moore, J.P.; Le, N.T.; Brandt, W.F.; Driouich, A.; Farrant, J.M. Towards a systems-based understanding of plant desiccation tolerance. Trends Plant Sci. 2009, 14, 110–117. [Google Scholar] [CrossRef]
- Jaramillo, M.; Browning, K.; Dever, T.E.; Blum, S.; Trachsel, H.; Merrick, W.C.; Ravel, J.M.; Sonenberg, N. Translation initiation factors that function as RNA helicases from mammals, plants and yeast. Biochim. Biophys. Acta Gene Regul. Mech. 1990, 1050, 134–139. [Google Scholar] [CrossRef]
- Rogers, G.W.; Richter, N.J.; Merrick, W.C. Biochemical and kinetic characterization of the RNA helicase activity of eukaryotic initiation factor 4A. J. Biol. Chem. 1999, 274, 12236–12244. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Pestova, T.V.; Hellen, C.U.; Tiedge, H. Translational control by a small RNA: Dendritic BC1 RNA targets the eukaryotic initiation factor 4A helicase mechanism. Mol. Cell. Biol. 2008, 28, 3008–3019. [Google Scholar] [CrossRef] [Green Version]
- Santosh Rama Bhadra Rao, T.; Vijaya Naresh, J.; Sudhakar Reddy, P.; Reddy, M.K.; Mallikarjuna, G. Expression of Pennisetum glaucum eukaryotic translational initiation factor 4A (PgeIF4A) confers improved drought, salinity, and oxidative stress tolerance in groundnut. Front. Plant Sci. 2017, 8, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Deng, G.; Fan, W.; Yuan, L.; Wang, H.; Zhang, P. NHX1 and eIF4A1-stacked transgenic sweet potato shows enhanced tolerance to drought stress. Plant Cell Rep. 2019, 38, 1427–1438. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Prasad, A.; Sharma, N.; Prasad, M. Linking the plant stress responses with RNA helicases. Plant Sci. 2020, 299, 110607. [Google Scholar] [CrossRef] [PubMed]
- Djukić, N.; Knežević, D.; Pantelić, D.; Živančev, D.; Torbica, A.; Marković, S. Expression of protein synthesis elongation factors in winter wheat and oat in response to heat stress. J. Plant Physiol. 2019, 240, 153015. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Liu, Z.; Cai, L.; Yan, X.; Hu, Y.; Hao, B.; Xu, Z.; Tian, Y.; Liu, X.; Liu, L.; et al. Nuclear encoded elongation factor EF-Tu is required for chloroplast development in rice grown under low-temperature conditions. J. Genet. Genom. 2022, 49, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Li, Y.; Qin, Z.; Guo, S.; Li, Y.; Miao, Y.; Song, C.; Chen, S.; Dai, S. Plant chloroplast stress response: Insights from thiol redox proteomics. Antioxid. Redox Signal. 2020, 33, 35–57. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: II. Purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol. 1977, 59, 315–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sárvári, É.; Gellén, G.; Sági-Kazár, M.; Schlosser, G.; Solymosi, K.; Solti, Á. Qualitative and quantitative evaluation of thylakoid complexes separated by Blue Native PAGE. Plant Methods 2022, 18, 23. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band | ACC# | Species | Protein Name | MW/kDa | No. of Peptides | % Cov |
|---|---|---|---|---|---|---|
| #1 | KZV21119.1 | Dorcoceras hygrometricum | Enolase | 47 | 3 | 11.3 |
| KZV25501.1 | Dorcoceras hygrometricum | UDP-D-apiose/UDP-D-xylose synthase 2 | 43 | 3 | 11.8 | |
| KZV20320.1 | Dorcoceras hygrometricum | V-type proton ATPase subunit H | 53 | 1 | 2.8 | |
| KZV34753.1 | Dorcoceras hygrometricum | Hypothetical protein F511_00655 (elongation factor Tu, chloroplastic) | 52 | 2 | 6.7 | |
| KZV24609.1 | Dorcoceras hygrometricum | Hypothetical protein F511_06435 (cytochrome P450) | 55 | 1 | 3.9 | |
| KZV18901.1 | Dorcoceras hygrometricum | Hypothetical protein F511_12006 (initiation factor 4A) | 47 | 2 | 7.0 | |
| #8 | KZV14262.1 | Dorcoceras hygrometricum | Triosephosphate isomerase | 23 | 2 | 13.9 |
| KZV28384.1 | Dorcoceras hygrometricum | Early light-induced protein | 20 | 1 | 7.3 | |
| KZV47616.1 | Dorcoceras hygrometricum | Hypothetical protein F511_12885 (desiccation-induced 1VOC superfamily) | 15 | 1 | 11.1 | |
| Q27U82 | Eucalyptus globulus | Pectin methylesterase (fragment) | 11 | 2 | 18.4 | |
| A0A2Z7AJ43 | Dorcoceras hygrometricum | Galactose mutarotase-like superfamily protein isoform 1 (fragment) | 36 | 1 | 4.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihailova, G.; Solti, Á.; Sárvári, É.; Hunyadi-Gulyás, É.; Georgieva, K. Protein Changes in Shade and Sun Haberlea rhodopensis Leaves during Dehydration at Optimal and Low Temperatures. Plants 2023, 12, 401. https://doi.org/10.3390/plants12020401
Mihailova G, Solti Á, Sárvári É, Hunyadi-Gulyás É, Georgieva K. Protein Changes in Shade and Sun Haberlea rhodopensis Leaves during Dehydration at Optimal and Low Temperatures. Plants. 2023; 12(2):401. https://doi.org/10.3390/plants12020401
Chicago/Turabian StyleMihailova, Gergana, Ádám Solti, Éva Sárvári, Éva Hunyadi-Gulyás, and Katya Georgieva. 2023. "Protein Changes in Shade and Sun Haberlea rhodopensis Leaves during Dehydration at Optimal and Low Temperatures" Plants 12, no. 2: 401. https://doi.org/10.3390/plants12020401