
A Tight Interaction between the Native Seagrass Cymodocea nodosa and the Exotic Halophila stipulacea in the Aegean Sea Highlights Seagrass Holobiont Variations



Abstract
:1. Introduction
2. Results
2.1. Abiotic Setting
2.2. Biotic Setting
2.3. Bacterial Communities
2.3.1. Bacterial Community Diversity
2.3.2. Environmental Bacterial Communities’ Composition and Indicator Species
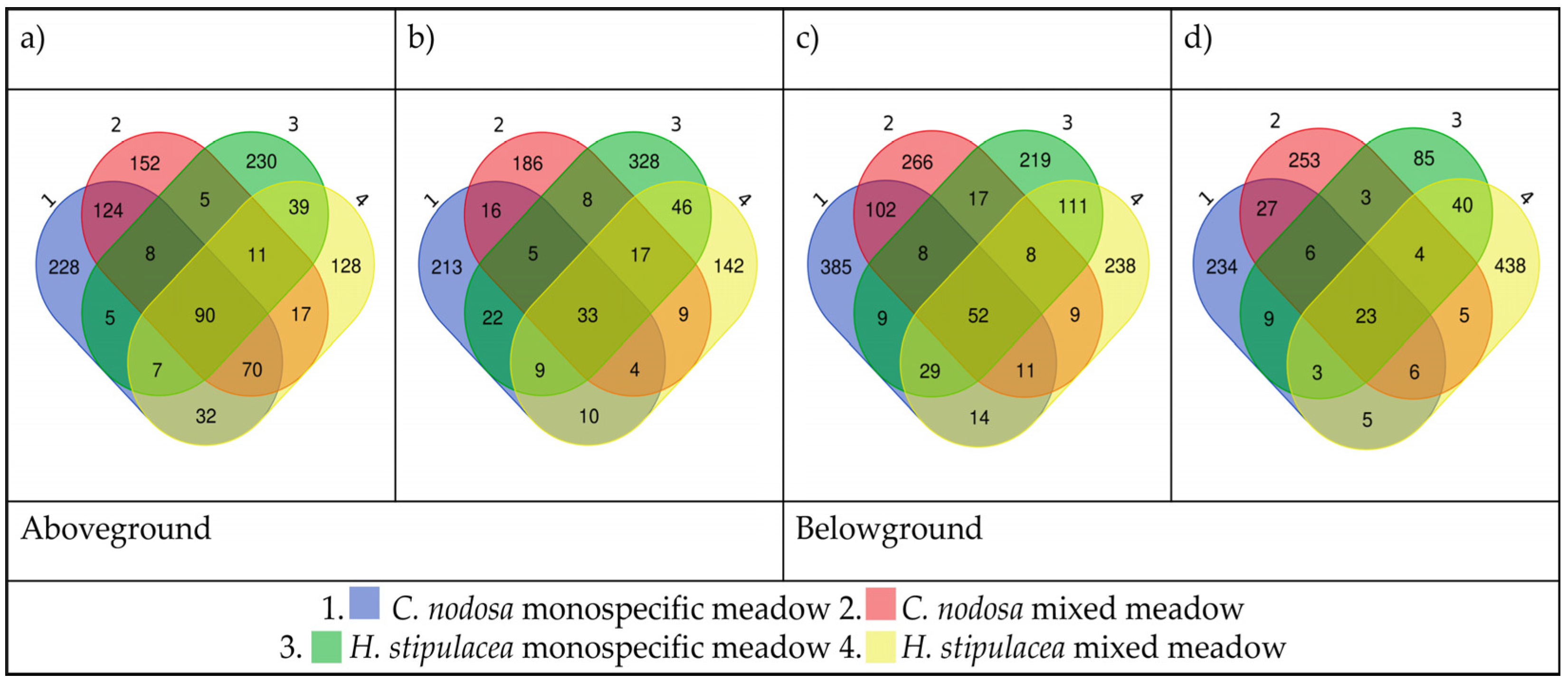
2.3.3. Composition of Seagrass Core Communities
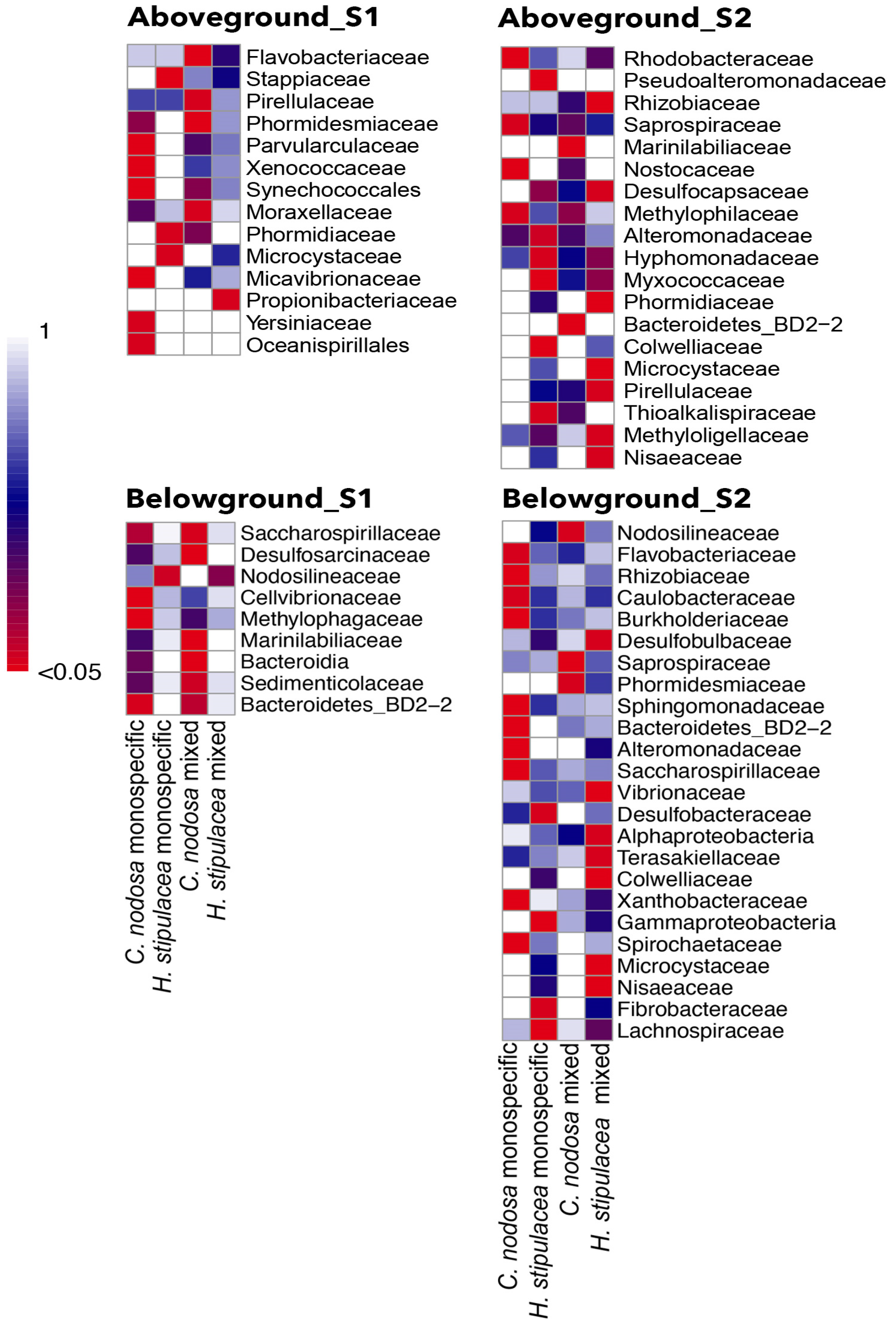
2.3.4. Host-Specific Bacterial Associations and Indicator Species
3. Discussion
3.1. The Plant Partner of the Holobiont Highlights Competition Dynamic
3.2. The Bacterial Partner of the Holobiont
3.2.1. The Bacterial Core May Support the Basic Plants’ Functions
3.2.2. Host-Specific Bacterial Component Highlights Different Seagrass–Microbiota Relationship under Competition
4. Materials and Methods
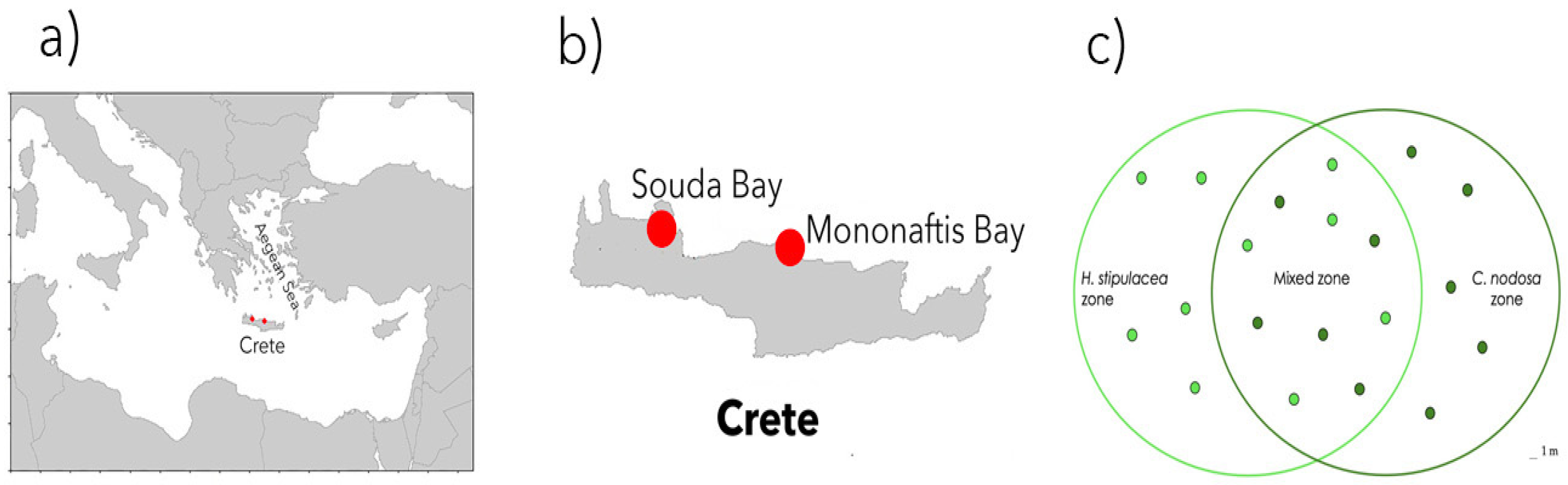
4.1. Sampling Design and Site Description
4.2. Abiotic Settings
4.3. Biotic Settings
4.4. Bacterial Community Analysis
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.; Zilber-Rosenberg, I. Microbes drive evolution of animals and plants: The hologenome concept. mBio 2016, 7, e01395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution after 10 years. Microbiome 2018, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, L.; Leray, M.; O’Dea, A.; Yuen, B.; Peixoto, R.S.; Pereira, T.J.; Bik, H.M.; Coil, D.A.; Duffy, J.E.; Herre, E.A.; et al. Host-associated microbiomes drive structure and function of marine ecosystems. PLoS Bio 2019, 17, e3000533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittami, S.M.; Arboleda, E.; Auguet, J.-C.; Bigalke, A.; Briand, E.; Cárdenas, P.; Cardini, U.; Decelle, J.; Engelen, A.H.; Eveillard, D.; et al. A community perspective on the concept of marine holobionts: Current status, challenges, and future directions. PeerJ 2021, 9, e10911. [Google Scholar] [CrossRef] [PubMed]
- Bang, C.; Dagan, T.; Deines, P.; Dubilier, N.; Duschl, W.J.; Fraune, S.; Hentschel, U.; Hirt, H.; Hülter, N.F.; Lachnit, T.; et al. Metaorganisms in extreme environments: Do microbes play a role in organismal adaptation? Zoology 2018, 127, 1–19. [Google Scholar] [CrossRef]
- Duarte, B.; Martins, I.; Rosa, R.; Matos, A.R.; Roleda, M.Y.; Reusch, T.B.H.; Engelen, A.H.; Serrão, E.A.; Pearson, G.A.; Marques, J.C.; et al. Climate change impacts on seagrass meadows and macroalgal forests: An integrative perspective on acclimation and adaptation potential. Front. Mar. Sci. 2018, 5, 190. [Google Scholar] [CrossRef] [Green Version]
- Simon, J.C.; Marchesi, J.R.; Mougel, C.; Selosse, M.A. Host-microbiota interactions: From holobiont theory to analysis. Microbiome 2019, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Ugarelli, K.; Chakrabarti, S.; Laas, P.; Stingl, U. The seagrass holobiont and its microbiome. Microorganisms 2017, 5, 81. [Google Scholar] [CrossRef] [Green Version]
- Crump, B.C.; Wojahn, J.M.; Tomas, F.; Mueller, R.S. Metatranscriptomics and amplicon sequencing reveal mutualisms in seagrass microbiomes. Front. Microbiol. 2018, 9, 388. [Google Scholar] [CrossRef]
- Seymour, J.R.; Laverock, B.; Nielsen, D.A.; Trevathan-Tackett, S.M.; Macreadie, P.I. The microbiology of seagrasses. In Seagrasses of Australia; Larkum, A.W.D., Kendrick, G.A., Ralph, P.J., Eds.; Springer: Cham, Switzerland, 2018; pp. 343–392. [Google Scholar] [CrossRef]
- Tarquinio, F.; Hyndes, G.A.; Laverock, B.; Koenders, A.; Säwström, C. The seagrass holobiont: Understanding seagrass-bacteria interactions and their role in seagrass ecosystem functioning. FEMS Microbiol. Lett. 2019, 366, fnz057. [Google Scholar] [CrossRef]
- Tarquinio, F.; Bourgoure, J.; Koenders, A.; Hyndes, G.A. Microorganisms facilitate uptake of dissolved organic nitrogen by seagrass leaves. ISME J. 2018, 12, 2796–2800. [Google Scholar] [CrossRef] [Green Version]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 2064, 1196–1206. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, X.; Zhang, Q.; Liu, F.; Zhang, J.; Gong, J. Seagrass (Zostera marina) colonization promotes the accumulation of diazotrophic bacteria and alters the relative abundances of specific bacterial lineages involved in benthic carbon and sulfur cycling. Appl. Environ. Microbiol. 2015, 81, 6901–6914. [Google Scholar] [CrossRef] [Green Version]
- Brodersen, K.E.; Michael, K.; Nielsen, D.A.; Pedersen, O.; Larkum, A.W.D. Rhizome, root/sediment interactions, aerenchyma and internal pressure changes in seagrasses. In Seagrasses of Australia; Larkum, A.W.D., Kendrick, G.A., Ralph, P.J., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 393–418. [Google Scholar]
- Brodersen, K.E.; Siboni, N.; Nielsen, D.A.; Pernice, M.; Ralph, P.J.; Seymour, J. Seagrass rhizosphere microenvironment alters plant-associated bacterial community composition. SFAM Environ. Microbiol. 2018, 20, 2854–2864. [Google Scholar] [CrossRef]
- Martin, B.C.; Bougoure, J.; Ryan, M.H.; Bennett, W.W.; Colmer, T.D.; Joyce, N.K.; Olsen, Y.S.; Kendrick, G.A. Oxygen loss from seagrass roots coincides with colonisation of sulphide-oxidising cable bacteria and reduces sulphide stress. ISME J. 2019, 13, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.C.; Gleeson, D.; Statton, J.; Siebers, A.R.; Grierson, P.; Ryan, M.H.; Kendrick, G.A. Low light availability alters root exudation and reduces putative beneficial microorganisms in seagrass roots. Front. Microbiol. 2018, 8, 2667. [Google Scholar] [CrossRef] [Green Version]
- Conte, C.; Rotini, A.; Manfra, L.; D’Andrea, M.M.; Winters, G.; Migliore, L. The seagrass holobiont: What we know and what we still need to disclose for its possible use as an ecological indicator. Water 2021, 13, 406. [Google Scholar] [CrossRef]
- Rivett, D.W.; Bell, T. Abundance determines the functional role of bacterial phylotypes in complex communities. Nat. Microbiol. 2018, 3, 767–772. [Google Scholar] [CrossRef]
- Conte, C.; Rotini, A.; Winters, G.; Vasquez, M.I.; Piazza, G.; Kletou, D.; Migliore, L. Elective affinities or random choice within the seagrass holobiont? The case of the native Posidonia oceanica (L.) Delile and the exotic Halophila stipulacea (Forssk.) Asch. from the same site (Limassol, Cyprus). Aquatic. Bot. 2021, 174, 103420. [Google Scholar] [CrossRef]
- Szitenberg, A.; Beca-Carretero, P.; Azcárate-García, T.; Yergaliyev, T.; Alexander-Shani, R.; Winters, G. Teasing apart the host-related, nutrient-related and temperature-related effects shaping the phenology and microbiome of the tropical seagrass Halophila stipulacea. Environ. Microbiome 2022, 17, 18. [Google Scholar] [CrossRef] [PubMed]
- Garcias-Bonet, N.; Eguíluz, V.; Díaz-Rúa, R.; Duarte, C. Host-association as major driver of microbiome structure and composition in Red Sea seagrass ecosystems. Environ. Microbiol. 2020, 23, 2021–2034. [Google Scholar] [CrossRef] [PubMed]
- Cúcio, C.; Engelen, A.H.; Costa, R.; Muyzer, G.; Costa, R. Rhizosphere microbiomes of European seagrasses are selected by the plant, but are not species specific. Front. Microbiol. 2016, 7, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejia, A.Y.; Rotini, A.; Lacasella, F.; Bookman, R.; Cristina, M.; Shem-tov, R.; Migliore, L. Assessing the ecological status of seagrasses using morphology, biochemical descriptors and bacterial community analyses. A study in Halophila stipulacea (Forsk) Aschers meadows in the northern Red Sea. Ecol. Ind. 2016, 60, 1150–1163. [Google Scholar] [CrossRef]
- Rotini, A.; Conte, C.; Seveso, D.; Montano, S.; Galli, P.; Vai, M.; Migliore, L.; Mejia, A. Daily variation of the associated bacterial community and the Hsp60 expression in the Maldivian seagrass Thalassia hemprichii. J. Sea. Res. 2020, 156, 101835. [Google Scholar] [CrossRef]
- Rotini, A.; Mejia, A.Y.; Costa, R.; Migliore, L.; Winters, G. Ecophysiological plasticity and bacteriome shift in the seagrass Halophila stipulacea along a depth gradient in the Northern Red Sea. Front. Plant Sci. 2017, 7, 2015. [Google Scholar] [CrossRef] [Green Version]
- Den Hartog, C. The Seagrasses of the World; North Holland Publishing, Co.: Amsterdam, The Netherland, 1970. [Google Scholar]
- Kuo, J.; den Hartog, C. Seagrass morphology, anatomy, and ultrastructure. In Seagrasses: Biology, Ecology and Conservation; Larkum, A., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 51–87. [Google Scholar]
- Peralta, G.; Godoy, O.; Egea, L.G.; de los Santos, C.B.; Jiménez-Ramos, R.; Lara, M.; Pérez-Lloréns, J.L. The morphometric acclimation to depth explains the long-term resilience of the seagrass Cymodocea nodosa in a shallow tidal lagoon. J. Environ. Manag. 2021, 299, 113452. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Bernard, G.; Pergent, G.; Shimabukuro, H.; Verlaque, M. Regression of Mediterranean seagrasses caused by natural processes and anthropogenic disturbances and stress: A critical review. Bot. Mar. 2009, 52, 395–418. [Google Scholar] [CrossRef]
- Banister, R.B.; Schwarz, M.T.; Fine, M.; Ritchie, K.B.; Muller, E.M. Instability and stasis among the microbiome of seagrass leaves, roots and rhizomes, and nearby sediments within a natural pH gradient. Microb. Ecol. 2022, 84, 703–716. [Google Scholar] [CrossRef]
- Korlević, M.; Markovski, M.; Zhao, Z.; Herndl, G.J.; Najdek, M. Seasonal dynamics of epiphytic bacterial community on marine macrophytes surface. Front. Microbiol. 2021, 12, 671342. [Google Scholar] [CrossRef]
- Winters, G.; Beer, S.; Willette, D.; Viana, I.G.; Chiquillo, K.L.; Beca-Carretero, P.; Villamayor, B.; Azcárate-García, T.; Shem-Tov, R.; Mwabvu, B.; et al. The tropical seagrass Halophila stipulacea: Reviewing what we know from its native and invasive habitats, alongside identifying knowledge gaps. Front. Mar. Sci. 2020, 7, 300. [Google Scholar] [CrossRef]
- Azcárate-García, T.; Beca-Carretero, P.; Villamayor, B.; Stengel, D.B.; Winters, G. Responses of the seagrass Halophila stipulacea to depth and spatial gradients in its native region (Red Sea): Morphology, in situ growth and biomass production. Aquat. Bot. 2020, 165, 103252. [Google Scholar] [CrossRef]
- Sharon, Y.; Beer, S. Diurnal movements of chloroplasts in Halophila stipulacea and their effect on PAM fluorometric measurements of photosynthetic rates. Aquat. Bot. 2008, 88, 273–276. [Google Scholar] [CrossRef]
- Sharon, Y.; Silva, J.; Santos, R.; Runcie, J.W.; Chernihovsky, M.; Beer, S. Photosynthetic responses of Halophila stipulacea to a light gradient. II. Acclimations following transplantation. Aquat. Biol. 2009, 7, 153–157. [Google Scholar] [CrossRef]
- Sharon, Y.; Dishon, G.; Beer, S. The effects of UV radiation on chloroplast clumping and photosynthesis in the seagrass Halophila stipulacea grown under high-PAR conditions. J. Mar. Sci. 2011, 2011, 483428. [Google Scholar] [CrossRef]
- Sharon, Y.; Levitan, O.; Spungin, D.; Berman-Frank, I.; Beer, S. Photoacclimation of the seagrass Halophila stipulacea to the DIM irradiance at its 48-meter depth limit. Lim. Ocean. 2011, 56, 357–362. [Google Scholar] [CrossRef]
- Van Tussenbroek, B.I.; van Katwijk, M.; Bouma, T.; van der Heide, T.; Govers, L.; Leuven, R. Non-native seagrass Halophila stipulacea forms dense mats under eutrophic conditions in the Caribbean. J. Sea Res. 2016, 115, 1–5. [Google Scholar] [CrossRef]
- Helber, S.B.; Winters, G.; Stuhr, M.; Belshe, E.F.; Bröhl, S.; Schmid, M.; Reuter, H.; Teichberg, M. Nutrient history affects the response and resilience of the tropical seagrass Halophila stipulacea to further enrichment in its native habitat. Front. Plant Sci. 2021, 12, 678341. [Google Scholar] [CrossRef]
- Apostolaki, E.T.; Holmer, M.; Santinelli, V.; Karakassis, I. Species-specific response to sulfide intrusion in native and exotic Mediterranean seagrasses under stress. Mar. Environ. Res. 2018, 134, 85–95. [Google Scholar] [CrossRef]
- Hernández-Delgado, E.A.; Toledo-Hernández, C.; Ruiz-Diaz, C.P.; Gómez-Andújar, N.X.; Medina-Muñiz, J.L.; Canals-Silander, M.F.; Suleimán-Ramos, S.E. Hurricane impacts and the resilience of the invasive Sea Vine, Halophila stipulacea: A case study from Puerto Rico. Estuar. Coasts 2020, 43, 1263–1283. [Google Scholar] [CrossRef]
- O’Brien, K.R.; Waycott, M.; Maxwell, P.; Kendrick, G.A.; Udy, J.W.; Ferguson, A.J.; Kilminster, K.; Scanes, P.; McKenzie, L.J.; McMahon, K.; et al. Seagrass ecosystem trajectory depends on the relative timescales of resistance, recovery and disturbance. Mar. Pollut. Bull. 2018, 134, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipkin, Y. Halophila stipulacea, a review of a successful immigration. Aquat. Bot. 1975, 1, 203–215. [Google Scholar] [CrossRef]
- Fritsch, C. Über die auffindung einer marinen Hydrocharidee im Mittelmeer. Verh. Zool. Bot. Ver. Wien 1895, 45, 104–106. [Google Scholar]
- Gambi, M.; Barbieri, F.; Bianchi, C. New record of the alien seagrass Halophila stipulacea (Hydrocharitaceae) in the western Mediterranean: A further clue to changing Mediterranean Sea biogeography. Mar. Biol. Records 2009, 2, E84. [Google Scholar] [CrossRef]
- Sghaier, Y.; Zakhama-Sraieb, R.; Benamer, I.; Charfi-Cheikhrouha, F. Occurrence of the seagrass Halophila stipulacea (Hydrocharitaceae) in the southern Mediterranean Sea. Bot. Mar. 2011, 54, 575–582. [Google Scholar] [CrossRef]
- Sghaier, Y.; Zakhama-Sraieb, R.; Charfi- Cheikhrouha, F. Effects of the invasive seagrass Halophila stipulacea on the native seagrass Cymodocea nodosa. In Proceedings of the 5ème Symposium Méditerranéen sur la Végétation Marine, Portorož, Slovénie, 27–28 October 2014. [Google Scholar]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.; Holon, F.; Agel, N.; Descamps, P.; Deter, J.; Pavy, T.; Delaruelle, G.; Verlaque, M. Distribution of the seagrass Halophila stipulacea: A big jump to the northwestern Mediterranean Sea. Aquat. Bot. 2022, 76, 103465. [Google Scholar] [CrossRef]
- Beca-Carretero, P.; Rotini, A.; Mejia, A.; Migliore, L.; Vizzini, S.; Winters, G. Halophila stipulacea descriptors in the native area (Red Sea): A baseline for future comparisons with native and non-native populations. Mar. Environ. Res. 2020, 153, 104828. [Google Scholar] [CrossRef]
- Beca-Carretero, P.; Teichberg, M.; Winters, G.; Procaccini, G.; Reuter, H. Projected rapid habitat expansion of tropical seagrass species in the Mediterranean Sea as climate change progresses. Front. Plant Sci. 2020, 11, 555376. [Google Scholar] [CrossRef]
- Ruiz, H.; Ballantine, D.L. Occurrence of the seagrass Halophila stipulacea in the tropical west Atlantic. Bull. Mar. Sci. 2004, 75, 131–135. [Google Scholar]
- Willette, D.A.; Ambrose, R.F. The distribution and expansion of the invasive seagrass Halophila stipulacea in Dominica, West Indies, with a preliminary report from St. Lucia. Aquat. Bot. 2009, 91, 137–142. [Google Scholar] [CrossRef]
- Vera, B.; Collado-Vides, L.; Moreno, C.; Van Tussenbroek, B.I. Halophila stipulacea (Hydrocharitaceae): A recent introduction to the continental waters of Venezuela. Carib. J. Sci. 2014, 48, 66–70. [Google Scholar] [CrossRef]
- Scheibling, R.E.; Patriquin, D.G.; Filbee-Dexter, K. Distribution and abundance of the invasive seagrass Halophila stipulacea and associated benthic macrofauna in Carriacou, Grenadines, Eastern Caribbean. Aquat. Bot. 2018, 144, 1–8. [Google Scholar] [CrossRef]
- Willette, D.A.; Ambrose, R.F. Effects of the invasive seagrass Halophila stipulacea on the native seagrass, Syringodium filiforme, and associated fish and epibiota communities in the Eastern Caribbean. Aquat. Bot. 2012, 103, 74–82. [Google Scholar] [CrossRef]
- Wahbeh, M.I.; Mahasneh, A.M. Heterotrophic bacteria attached to leaves, rhizomes and roots of three seagrass species from Aqaba (Jordan). Aquat. Bot. 1984, 20, 87–96. [Google Scholar] [CrossRef]
- Pereg, L.L.; Lipkin, Y.; Sar, N. Different niches of the Halophila stipulacea seagrass bed harbor distinct populations of nitrogen fixing bacteria. Mar. Biol. 1994, 119, 327–333. [Google Scholar] [CrossRef]
- Weidner, S.; Arnold, W.; Pühler, A. Diversity of uncultured microorganisms associated with the seagrass Halophila stipulacea estimated by restriction fragment length polymorphism analysis of PCR-amplified 16s rRNA genes. Appl. Environ. Microbiol. 1996, 62, 766–771. [Google Scholar] [CrossRef] [Green Version]
- Weidner, S.; Arnold, W.; Stackebrandt, E.; Pühler, A. Phylogenetic analysis of bacterial communities associated with leaves of the seagrass Halophila stipulacea by a culture-independent small-subunit rRNA gene approach. Microb. Ecol. 2000, 39, 22–31. [Google Scholar] [CrossRef]
- Aires, T.; Stuij, T.M.; Muyzer, G.; Serrão, E.A.; Engelen, A.H. Characterization and comparison of bacterial communities of an invasive and two native Caribbean seagrass species sheds light on the possible influence of the microbiome on invasive mechanisms. Front. Microbiol. 2021, 12, 653998. [Google Scholar] [CrossRef]
- Othman, A.A. Nitrogen-fixing bacteria and change of the macrophyte vegetations in Bardawil Lagoon, Egypt. Egypt. J. Aquat. Biol. Fish. 2022, 24, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Georgiou, D.; Alexandre, A.; Luis, J.; Santos, R. Temperature is not a limiting factor for the expansion of Halophila stipulacea throughout the Mediterranean Sea. Mar. Ecol. Prog. Ser. 2016, 544, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Wesselmann, M.; Anton, A.; Duarte, C.M.; Hendriks, I.E.; Agustí, S.; Savva, I.; Apostolaki, E.T.; Marbà, N. Tropical seagrass Halophila stipulacea shifts thermal tolerance during Mediterranean invasion. Proc. Biol. Sci. 2020, 287, 20193001. [Google Scholar] [CrossRef] [PubMed]
- Wesselmann, M.; Chefaoui, R.M.; Marbà, N.; Serrao, E.A.; Duarte, C.M. Warming threatens to propel the expansion of the exotic seagrass Halophila stipulacea. Front. Mar. Sci. 2021. [CrossRef]
- Di Genio, S.; Gaglioti, M.; Meneghesso, C.; Barbieri, F.; Cerrano, C.; Gambi, M.C. Phenology and ecology of the alien seagrass Halophila stipulacea in its northern range limit in the Mediterranean Sea. Aquat. Bot. 2021, 168, 103304. [Google Scholar] [CrossRef]
- Fahimipour, A.K.; Kardish, M.R.; Lang, J.M.; Green, J.L.; Eisen, J.A.; Stachowicz, J.J. Global scale structure of the eelgrass microbiome. Appl. Environ. Microbiol. 2017, 83, e03391-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, E.P.; Ng, H.J.; Webb, H.K. The Family Pseudoalteromonadaceae. In The Prokaryotes—Gammaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Nakagawa, T.; Tsuchiya, Y.; Ueda, S.; Fukui, M.; Takahashi, R. Eelgrass sediment microbiome as a nitrous oxide sink in brackish Lake Akkeshi, Japan. Microbes Environ. 2019, 34, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Toyonaga, M.; Ohashi, A.; Tourlousse, D.M.; Matsuura, N.; Meng, X.-Y.; Tamaki, H.; Hanada, S.; Cruz, R.; Yamaguchi, T.; et al. Lentimicrobium saccharophilum gen. nov., sp. nov., a strictly anaerobic bacterium representing a new family in the phylum Bacteroidetes, and proposal of Lentimicrobiaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 2635–2642. [Google Scholar] [CrossRef] [Green Version]
- Dedysh, S.N.; Losey, N.A.; Lawson, P. Thermoanaerobaculaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; Trujillo, M.E., Dedysh, S., DeVos, P., Hedlund, B., Kämpfer, P., Rainey, F.A., Whitman, W.B., Eds.; John Wiley & Sons, Inc.: New Jersey, USA, 2021; in association with Bergey’s Manual Trust. [Google Scholar] [CrossRef]
- Muwawa, E.M.; Obieze, C.C.; Makonde, H.M.; Jefwa, J.M.; Kahindi, J.H.P.; Khasa, D.P. 16S rRNA gene amplicon-based metagenomic analysis of bacterial communities in the rhizospheres of selected mangrove species from Mida Creek and Gazi Bay, Kenya. PLoS ONE 2021, 16, e0248485. [Google Scholar] [CrossRef]
- Liu, S.; Jiang, Z.; Wu, Y.; Zhang, J.; Arbi, I.; Ye, F.; Huang, X.; Macreadie, P.I. Effects of nutrient load on bacterial activities within a seagrass-dominated ecosystem: Implications of changes in seagrass blue carbon. Mar. Pollut. Bull. 2017, 117, 214–221. [Google Scholar] [CrossRef]
- Sun, Y.; Song, Z.; Zhang, H.; Liu, P.; Hu, X. Seagrass vegetation affect the vertical organization of bacterial communities in sediment. Mar. Environ. Res. 2021, 162, 105174. [Google Scholar] [CrossRef]
- López-Pérez, M.; Rodriguez-Valera, F. The Family Alteromonadaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Trevathan-Tackett, S.M.; Seymour, J.; Nielsen, D.A.; Macreadie, P.I.; Jeffries, T.; Sanderman, J.; Baldock, J.; Howes, J.M.; Steven, A.D.L.; Ralph, P. Sediment anoxia limits bacterial-driven seagrass carbon remineralization under warming conditions. FEMS Microb. Ecol. 2017, 93, fix033. [Google Scholar] [CrossRef] [Green Version]
- Olsen, I. The Family Fusobacteriaceae. In The Prokaryotes—Gammaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Wasmund, K.; Pelikan, C.; Schintlmeister, A.; Wagner, M.; Watzka, M.; Richter, A.; Bhatnagar, S.; Noel, A.; Hubert, C.R.J.; Rattei, T.; et al. Genomic insights into diverse bacterial taxa that degrade extracellular DNA in marine sediments. Nat. Microbiol. 2019, 6, 885–898. [Google Scholar] [CrossRef]
- Dashiff, A.; Junka, R.A.; Libera, M.; Kadouri, D.E. Predation of human pathogens by the predatory bacteria Micavibrio aeruginosavorus and Bdellovibrio bacteriovorus. J. App. Microbiol. 2011, 110, 431–444. [Google Scholar] [CrossRef]
- Sarhan, M.S.; Hamza, M.A.; Youssef, H.H.; Patz, S.; Becker, M.; ElSawey, H.; Nemr, R.; Daanaa, H.-S.A.; Mourad, E.F.; Morsi, A.T.; et al. Culturomics of the plant prokaryotic microbiome and the dawn of plant-based culture media—A review. J. Adv. Res. 2019, 19, 15–27. [Google Scholar] [CrossRef]
- Alotaibi, B.S.; Khan, M.; Shamim, S. Unraveling the Underlying Heavy Metal Detoxification Mechanisms of Bacillus species. Microorganisms 2021, 9, 1628. [Google Scholar] [CrossRef]
- Uku, J.; Björk, M.; Bergman, B.; Díez, B. Characterization and comparison of prokaryotic epiphytes associated with three East African seagrasses. J. Phycol. 2007, 43, 768–779. [Google Scholar] [CrossRef]
- Cavalcanti, G.; Thompson, F. The Family Parvularculaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Tarquinio, F.; Attlan, O.; Vanderklift, M.A.; Berry, O.; Bissett, A. Distinct endophytic bacterial communities inhabiting seagrass seeds. Front. Microb. 2021, 12, 703014. [Google Scholar] [CrossRef]
- McBride, M.J. The Family Flavobacteriaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Teixeira, L.; Merquior, V. The family Moraxellaceae. In The Prokaryotes.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Onishi, Y.; Mohri, Y.; Tuji, A.; Ohgi, K.; Yamaguchi, A.; Imai, I. The seagrass Zostera marina harbors growth-inhibiting bacteria against the toxic dinoflagellate Alexandrium tamarense. Fish Sci. 2014, 80, 353–362. [Google Scholar] [CrossRef]
- Kohn, T.; Rast, P.; Kallscheuer, N.; Wiegand, S.; Boedeker, C.; Jetten, M.S.M.; Jeske, O.; Vollmers, J.; Kaster, A.-K.; Rohde, M.; et al. The Microbiome of Posidonia. oceanica seagrass leaves can be dominated by Planctomycetes. Front. Microbiol. 2020, 11, 1458. [Google Scholar] [CrossRef]
- Delmont, O.T.; Pierella Karlusich, J.J.; Veseli, I.; Fuessel, J.; Eren, A.M.; Foster, R.; Bowler, C.; Wincker, P.; Pelletier, E. Heterotrophic bacterial diazotrophs are more abundant than their cyanobacterial counterparts in metagenomes covering most of the sunlit ocean. ISME J. 2020, 16, 927–936. [Google Scholar] [CrossRef]
- Rubio-Portillo, E.; Martin-Cuadrado, A.-B.; Ramos-Esplá, A.Á.; Antón, J. Metagenomics unveils Posidonia oceanica “banquettes” as a potential source of novel bioactive compounds and carbohydrate active enzymes (CAZymes). mSystems 2021, 6, e00866-21. [Google Scholar] [CrossRef]
- Heimann, K.; Cirés, S. N2-fixing cyanobacteria: Ecology and biotechnological applications. In Handbook of Marine Microalgae; Se-Kwon, K., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 33, pp. 501–515. [Google Scholar] [CrossRef]
- Weber, C.; King, G. Physiological, ecological, and phylogenetic characterization of Stappia, a marine CO-oxidizing bacterial genus. Appl. Environ. Microbiol. 2007, 73, 1266–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Chen, W.; Wang, G.; Dai, S.; Zhou, D.; Zhao, H.; Guo, Y.; Ouyang, Y.; Li, X. Culture-dependent diversity of Actinobacteria associated with seagrass (Thalassia hemprichii). Afr. J. Microbiol. Res. 2012, 6, 87–94. [Google Scholar] [CrossRef]
- Pujalte, M.J.; Lucena, T.; Ruvira, M.A.; Arahal, D.R.; Macin, M.C. The family Rhodobacteraceae. In The Prokaryotes—Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 439–512. [Google Scholar]
- Elifantz, H.; Horn, G.; Ayon, M.; Cohen, Y.; Minz, D. Rhodobacteraceae are the key members of the bacterial community of the initial biofilm formed in Eastern Mediterranean coastal seawater. FEMS Microbiol. Ecol. 2013, 85, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Hamisi, M.; Díez, B.; Lyimo, T.; Ininbergs, K.; Bergman, B. Epiphytic cyanobacteria of the seagrass Cymodocea rotundata: Diversity, diel nifH expression and nitrogenase activity. Environ. Microbiol. Rep. 2013, 5, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Doronina, N.; Kaparullina, E.; Trotsenko, Y. The Family Methylophilaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- McIlroy, S.J.; Nielsen, P.H. The Family Saprospiraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.; Jiang, Z.; Wu, Y.; Huang, X. Gradient of microbial communities around seagrass roots was mediated by sediment grain size. Ecosph. 2022, 13, e3942. [Google Scholar] [CrossRef]
- Ludwig, W.; Euzéby, J.; Whitman, W.B. Family II. Marinilabiliaceae fam. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Krieg, N.R., Staley, J.T., Brown, D.R., Hedlund, B.P., Paster, B.J., Ward, N.L., Ludwig, W., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2010; Volume 4, p. 49. [Google Scholar]
- Holmström, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microb. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Garcia, R.; Müller, R. The Family Myxococcaceae. In The Prokaryotes—Deltaproteobacteria and Epsilonproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Mohr, W.; Lehnen, N.; Ahmerkamp, S.; Marchant, H.K.; Graf, J.S.; Tschitschko, B.; Yilmaz, P.; Littmann, S.; Gruber-Vodicka, H.; Leisch, N.; et al. Terrestrial-type nitrogen-fixing symbiosis between seagrass and a marine bacterium. Nature 2021, 600, 105–109. [Google Scholar] [CrossRef]
- Mori, K.; Suzuki, K. The Family Thioalkalispiraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Abraham, W.R.; Rohde, M. The Family Hyphomonadaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Abanda-Nkpwatt, D.; Müsch, M.; Tschiersch, J.; Boettner, M.; Schwab, W. Molecular interaction between Methylobacterium extorquens and seedlings: Growth promotion, methanol consumption, and localization of the methanol emission site. J. Exp. Bot. 2006, 57, 4025–4032. [Google Scholar] [CrossRef]
- Song, J.; Hwang, J.; Kang, I.; Cho, J.-C. A sulfate-reducing bacterial genus, Desulfosediminicola gen. nov., comprising two novel species cultivated from tidal-flat sediments. Sci. Rep. 2011, 11, 19978. [Google Scholar] [CrossRef]
- Glaeser, S.P.; Kämpfer, P. The Family Sphingomonadaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Agnolucci, M.; Avio, L.; Pepe, A.; Turrini, A.; Cristani, C.; Bonini, P.; Cirino, V.; Colosimo, F.; Ruzzi, M.; Giovannetti, M. Bacteria associated with a commercial Mycorrhizal inoculum: Community composition and multifunctional activity as assessed by Illumina sequencing and culture-dependent tools. Front. Plant Sci. 2019, 9, 1956. [Google Scholar] [CrossRef]
- Kumar, M.; Kour, D.; Yadav, A.N.; Saxena, R.; Rai, P.K.; Jyoti, A.; Tomar, R.S. Biodiversity of methylotrophic bacterial communities and their potential role in mitigation of abiotic stresses in plants. Biologia 2019, 74, 287–308. [Google Scholar] [CrossRef]
- Wang, L.; English, M.K.; Tomas, F.; Mueller, R.S. Recovery and community succession of the Zostera marina rhizobiome after transplantation. Appl. Environ. Microbiol. 2021, 87, e02326-20. [Google Scholar] [CrossRef]
- Hurtado-McCormick, V.; Kahlke, T.; Petrou, K.; Jeffries, T.; Ralph, P.J.; Seymour, J.R. Regional and microenvironmental scale characterization of the Zostera muelleri seagrass microbiome. Front. Microbiol. 2019, 10, 1011. [Google Scholar] [CrossRef]
- Leadbeater, D.R.; Oates, N.C.; Bennett, J.P.; Li, Y.; Dowle, A.A.; Taylor, J.D.; Alponti, J.S.; Setchfield, A.T.; Alessi, A.M.; Helgason, T.; et al. Mechanistic strategies of bacterial communities regulating lignocellulose deconstruction in a UK salt marsh. Microbiome 2021, 9, 48. [Google Scholar] [CrossRef]
- Ettinger, C.L.; Voerman, S.E.; Lang, J.M.; Stachowicz, J.J.; Eisen, J.A. Bacterial communities in sediment from Zostera marina patches, but not the Z. marina leaf or root microbiomes, vary in relation to distance from patch edge. PeerJ 2021, 5, e3246. [Google Scholar] [CrossRef] [Green Version]
- Bourque, A.S.; Vega-Thurber, R.; Fourqurean, J.W. Bacterial community structure and dynamics in restored subtropical seagrass sediments. Aquat. Microb. Ecol. 2015, 74, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Abraham, W.R.; Rohde, M.; Bennasar, A. The Family Caulobacteraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Mhedbi-Hajri, N.; Jacques, M.A.; Koebnik, R. Adhesion Mechanisms of Plant-Pathogenic Xanthomonadaceae. In Bacterial Adhesion Advances in Experimental Medicine and. Biology; Linke, D., Goldman, A., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 715, pp. 71–89. [Google Scholar] [CrossRef]
- Garcias-Bonet, N.; Arrieta, J.M.; de Santana, C.N.; Duarte, C.M.; Marbà, N. Endophytic bacterial community of a Mediterranean marine angiosperm (Posidonia oceanica). Front. Microbiol. 2012, 3, 342. [Google Scholar] [CrossRef] [Green Version]
- Rivas, L.R. A Reinterpretation of the Concepts “Sympatric” and “Allopatric” with Proposal of the Additional Terms “Syntopic” and “Allotopic”. Syst. Biol. 1964, 13, 42–43. [Google Scholar] [CrossRef]
- Martin, A.; Manriquez, B.; Pompa, C.; Saper, A.; Grice, K.A.; Bystriansky, J. On the composition of Cymodocea nodosa root exudate under artificial blue, green and natural light conditions. DePaul. Discov. 2019, 8, 11. Available online: https://via.library.depaul.edu/depaul-disc/vol8/iss1/11 (accessed on 1 December 2022).
- Masson-Boivin, C.; Sachs, J.L. Symbiotic nitrogen fixation by rhizobia—The roots of a success story. Curr. Opin. Plant Biol. 2018, 44, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Marbà, N.; Duarte, C.M. Growth and sediment space occupation by seagrass Cymodocea nodosa roots. Mar. Ecol. Prog. Ser. 2001, 224, 291–298. [Google Scholar] [CrossRef]
- Morris, J.; Hartl, D.L.; Knoll, A.H.; Robert, A.L.; Michael, M. Biologia: Come Funziona la Vita, 3rd ed.; Zanichelli: Bologna, Italy, 2011. [Google Scholar]
- Ben Brahim, M.; Mabrouk, L.; Hamza, A.; Jribi, I. Comparison of spatial scale variability of shoot density and epiphytic leaf assemblages of Halophila stipulacea and Cymodocea nodosa on the Eastern coast of Tunisia. Plant Biosyst. 2020, 154, 413–426. [Google Scholar] [CrossRef]
- Migliore, L.; Rotini, A.; Randazzo, D.; Albanese, N.N.; Giallongo, A. Phenols content and 2-D electrophoresis protein pattern: A promising tool to monitor Posidonia meadows health state. BMC Ecol. 2007, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotini, A.; Anello, L.; Di Bernardo, M.; Giallongo, A.; Valiante, L.; Migliore, L. Comparative analysis of bed density, total phenol content and protein expression pattern in Posidonia oceanica (L.) Delile. Open J. Ecol. 2013, 3, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.; Barrote, I.; Costa, M.M.; Albano, S.; Santos, R. Physiological responses of Zostera marina and Cymodocea nodosa to light-limitation stress. PLoS ONE 2013, 8, e81058. [Google Scholar] [CrossRef] [PubMed]
- Ceccherelli, G.; Oliva, S.; Pinna, S.; Piazzi, L.; Procaccini, G.; Marin-Guirao, L.; Dattolo, E.; Gallia, R.; La Manna, G.; Gennaro, P.; et al. Seagrass collapse due to synergistic stressors is not anticipated by phenological changes. Oecologia 2018, 186, 1137–1152. [Google Scholar] [CrossRef]
- Mannino, A.M.; Micheli, C. Ecological function of phenolic compounds from Mediterranean fucoid algae and seagrasses: An overview on the genus Cystoseira sensu lato and Posidonia oceanica (L.) Delile. J. Mar. Sci. Eng. 2021, 8, 19. [Google Scholar] [CrossRef]
- Cuny, P.; Serve, L.; Jupin, H.; Boudouresque, C.F. Water soluble phenolic compounds of the marine phanerogam Posidonia oceanica in a Mediterranean area colonised by the introduced chlorophyte Caulerpa taxifolia. Aquat. Bot. 1995, 52, 237–242. [Google Scholar] [CrossRef]
- Dumay, O.; Costa, J.; Desjobert, J.M.; Pergent, G. Variations in the concentration of phenolic compounds in the seagrass Posidonia oceanica under conditions of competition. Phytochem 2004, 65, 3211–3220. [Google Scholar] [CrossRef]
- Pergent, G.; Boudouresque, C.F.; Dumay, O.; Pergent-Martini, C.; Wyllie-Echeverria, S. Competition between the invasive macrophyte Caulerpa taxifolia and the seagrass Posidonia oceanica: Contrasting strategies. BMC Ecol. 2008, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.E.; Fourqurean, J.W. Interspecific variation in the elemental and stable isotope content of seagrasses in South Florida. Mar. Ecol. Prog. Ser. 2009, 387, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M. Seagrass nutrient content. Mar. Ecol. Progr. Ser. 1990, 6, 201–207. [Google Scholar] [CrossRef]
- Roth-Schulze, A.J.; Zozaya-Valdés, E.; Steinberg, P.D.; Thomas, T. Partitioning of functional and taxonomic diversity in surface-associated bacterial communities. Environ. Microbiol. 2016, 18, 4391–4402. [Google Scholar] [CrossRef] [PubMed]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Willems, A. The family Comamonadaceae. In The Prokaryotes—Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 777–851. [Google Scholar]
- Han, J.; Sun, L.; Dong, X.; Cai, Z.; Sun, X.; Yang, H.; Wang, Y.; Song, W. Characterization of a novel plant growth-promoting bacteria strain Delftia tsuruhatensis HR4 both as a diazotroph and a potential biocontrol agent against various plant pathogens. Syst. Appl. Microbiol. 2005, 28, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Rothballer, M.; Engel, M.; Hoser, J.; Schmidt, T. Phenotypic variation in Acidovorax radicis N35 influences plant growth promotion. FEMS Microbiol. Ecol. 2012, 7, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.; Li, T.; Chen, M.; Huang, G. Cross-ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Env. Microbiol. 2008, 74, 52–60. [Google Scholar] [CrossRef]
- Oren, A. The Family Rhodocyclaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Dimitrieva, G.Y.; Crawford, R.L.; Yüksel, G.U. The nature of plant growth-promoting effects of a pseudoalteromonadaceae associated with the marine algae Laminaria japonica and linked to catalase excretion. J. Appl. Microbiol. 2006, 100, 1159–1169. [Google Scholar] [CrossRef]
- Garrity, G.M.; Bell, J.A.; Lilburn, T.; Family, I. Rhodobacteraceae fam. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Garrity, G.M., Eds.; Springer: New York, NY, USA, 2005; Volume 2, p. 161. [Google Scholar]
- Nešvera, J.; Rucká, L.; Pátek, M. Catabolism of Phenol and Its Derivatives in Bacteria: Genes, Their Regulation, and Use in the Biodegradation of Toxic Pollutants. Adv. Appl. Microbiol. 2015, 93, 107–160. [Google Scholar] [CrossRef]
- Salas, A.; Cabrera, J.J.; Jiménez-Leiva, A.; Mesa, S.; Bedmar, E.J.; Richardson, D.J.; Gates, A.J.; Delgado, M.J. Bacterial nitric oxide metabolism: Recent insights in rhizobia. Adv. Microb. Physiol. 2021, 78, 259–315. [Google Scholar] [CrossRef]
- Miroshnichenko, M.L.; Kostrikina, N.A.; Chernyh, N.A.; Pimenov, N.V.; Tourova, T.P.; Antipov, A.N.; Spring, S.; Stackebrandt, E.; Bonch-Osmolovskaya, E.A. Caldithrix abyssi gen. nov., sp. nov., a nitrate-reducing, thermophilic, anaerobic bacterium isolated from a Mid-Atlantic Ridge hydrothermal vent, represents a novel bacterial lineage. Int. J. Syst. Evol. Microbiol. 2003, 53, 323–329. [Google Scholar] [CrossRef]
- Zhang, W.; Zhou, X.; Yuan, Y.; Liu, B.; Epstein, S.S.; He, S. Complete genome sequence of Saccharospirillum mangrovi HK-33T sheds light on the ecological role of a bacterium in mangrove sediment environment. Curr. Microbiol. 2019, 76, 231–236. [Google Scholar] [CrossRef]
- Bengtsson, M.; Sjøtun, K.; Lanzén, A.; Øvreås, L. Bacterial diversity in relation to secondary production and succession on surfaces of the kelp Laminaria. yperborean. ISME J. 2012, 6, 2188–2198. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Fish. Res. Board of Canada, Bull. 167; The Alger Press Ltd.: Ottawa, Ontario, Canada, 1972. [Google Scholar]
- Ivancic, I.; Degobbis, D. An optimal manual procedure for Ammonia analysis in natural waters by the indophenol blue method. Water Res. 1974, 18, 1143–1147. [Google Scholar] [CrossRef]
- Vizzini, S.; Mazzola, A. The effects of anthropogenic organic matter inputs on stable carbon and nitrogen isotopes in organisms from different trophic levels in a southern Mediterranean coastal area. Sci. Total Environ. 2006, 368, 723–731. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Meth. 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and characterization by UV-VIS Spectroscopy. Curr. Prot. Food Analytic. Chem. 2012, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Kadivar, H.; Stapleton, A.E. Ultraviolet radiation alters maize phyllosphere bacterial diversity. Microb. Ecol. 2003, 45, 353–361. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Meth. 2016, 13, 581. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2016, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeont. Electron. 2002, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 15 January 2021).
- RStudio Team. RStudio: Integrated Development for R. RStudio; PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 15 January 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bottom Water | Site #1 | Site #2 |
|---|---|---|
| NH4+ (µM) | 0.39 ± 0.16 | 0.6 ± 0.35 |
| NO3− (µM) | 0.46 ± 0.17 A | 0.14 ± 0.00 B |
| NO2 (µM) | 0.00 ± 0.00 | 0.00 ± 0.00 |
| PO43− (µM) | 0.06 ± 0.01 | 0.03 ± 0.01 |
| Site | Site #1 | Site #2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | C. nodosa | H. stipulacea | Mixed | C. nodosa | H. stipulacea | Mixed | ||||
| Pore water | NH4+(µM) | 5.7 ± 3.2 | 4.8 ± 2.2 | 2.3 ± 0.3 | 2.3 ± 1.0 | 2.4 ± 0.6 | 3.3 ± 1.3 | |||
| PO43− (µM) | 0.4 ± 0.2 | 0.3 ± 0.0 | 0.4 ± 0.0 | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.5 ± 0.1 | ||||
| Sediment | TC (%) | 8.4 ± 0.9 A | 8.3 ± 1.2 A | 8.2 ± 1.0 A | 4.2 ± 0.3 B | 5.7 ± 0.7 B | 4.7 ± 0.5 B | |||
| TN (%) | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | ||||
| δ13C (‰) | −16.8 ± 0.8 Aa | −18.4 ± 1.1 Aa | −15.6 ± 0.8 Ab | −24.4 ± 0.8 Ba | −20.7 ± 0.9 Bb | −22.5 ± 0.4 Ba | ||||
| δ15Ν (‰) | 1.6 ± 0.8 Aa | 0.3 ± 0.1 b | 0.6 ± 0.0 | −0.9 ± 0.3 B | 0.1 ± 0.2 | 0.0 ± −0.2 | ||||
| C/N | 317.0 ± 6.0 | 318.7 ± 6.0 | 406.1 ± 8.0 A | 301.5 ± 75.0 | 199.6 ± 34.5 | 213.4 ± 38.5 B | ||||
| Sediment granulometry | ||||||||||
| Size (mm) | >1000 | 1000–250 | 250–125 | 125–63 | <63 | >1000 | 1000–250 | 250–125 | 125–63 | <63 |
| (%) | 16.4 | 71.5 | 4.8 | 4.0 | 3.4 | 14.1 | 74.7 | 10.7 | 0.5 | 0.1 |
| SITE #1—Shallow | SITE #2—Deep | |||||||
|---|---|---|---|---|---|---|---|---|
| Species | C. nodosa | H. stipulacea | C. nodosa | H. stipulacea | ||||
| Meadow | Monospecific | Mixed | Monospecific | Mixed | Monospecific | Mixed | Monospecific | Mixed |
| Shoot density (m−2) SE | 2059.80 413.0 Aa | 486.9 114.0 Ab | 3497.6 554.0 | 4640.0 1028.0 | 306.4 33.0 B | 134.0 47.0 B | 3017.0 209.0 | 2960.0 44.5 |
| Area (cm−2) SE | 9.2 0.3 a | 8.4 0.6 b | 1.0 0.1 | 0.7 0.2 A | 10.8 0.9 a | 8.4 1.5 b | 1.4 0.2 | 1.3 0.1 B |
| TLR * (m m−2) SE | 5.9 2.2 Aa | 1.8 0.9 Ab | 5.2 2.1 | 6.3 2.4 A | 0.4 0.2 B | 0.4 0.1 B | 3.7 0.8 | 4.3 0.2 B |
| Leaves | ||||||||
| Area (cm−2) | 9.2 0.3 a | 8.4 0.6 b | 1.0 0.1 | 0.7 0.2 A | 10.8 0.9 a | 8.4 1.5 b | 1.4 0.2 | 1.3 0.1 B |
| Chl a (mg g−1) | 0.5 ± 0.0 | 0.5 ± 0.0 | 0.5 ± 0.1 | 0.3 ± 0.12 | 1.0 ± 0.3 | 0.5 ± 0.3 | 0.5 ± 0.2 | 0.5 ± 0.3 |
| Chl b (mg g−1) | 0.1 ± 0.0 a | 0.3 ± 0.1 b | 0.3 ± 0.0 a | 0.5 ± 0.2 Ab | 0.2 ± 0.3 | 0.2 ± 0.1 | 0.1 ± 0.2 | 0.1 ± 0.1 B |
| Chl a/b ratio | 0.2 ± 0.1 | 0.1 ± 0.1 A | 0.1 ± 0.0 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.0 B | 0.3 ± 0.1 | 0.2 ± 0.1 |
| Car (mg g−1) | 5.0 ± 0.7 | 2.0 ± 0.00 | 1.8 ± 0.2 | 0.4 ± 0.1 | 4.4 ± 0. | 1.3 ± 1.3 | 4.6 ± 1.2 | 4.01 ± 1.9 |
| Chl/Car ratio | 7.0 ± 4.1 | 4.2 ± 0.9 | 6.7 ± 2.4 | 6.1 ± 0.7 | 5.9 ± 1.9 | 2.7 ± 1.2 | 2.0 ± 0.6 | 3.5 ± 2.2 |
| Total phenols (mg g−1) | 3.1 ± 0.5 a | 5.7 ± 0.5 Ab | 3.0 ± 1.8 Aa | 5.8 ± 0.8 Ab | 5.6 ± 1.2a | 10.1 ± 3.1 Bb | 5.67 ± 1.3 Ba | 8.52 ± 1.6 Bb |
| TC (%) | 33.8 ± 0.4 Aa | 33.2 ± 0.8 Aa | 28.7 ± 0.4 Ab | 29.1 ± 0.4 Ab | 36 ± 0.5 Baa | 30.6 ± 0.5 Bb | 26.7 ± 0.5 Bab | 27.6 ± 1.2 Bb |
| TN (%) | 1.6 ± 0.1 | 1.4 ± 0.1 | 2.1 ± 0.1 | 1.8 ± 0.1 | 1.3 ± 0.01 | 1.3 ± 0.1 | 1.4 ± 0.1 | 1.5 ± 0.1 |
| C/N | 20.1 ± 1.3 Aa | 22.3 ± 0.6 a | 13.4 ± 0.7 ab | 13.4 ± 0.7 bb | 26.0 ± 21.0 Baa | 22.3 ± 1.2 ba | 18.3 ± 0.6 b | 18.2 ± 0.8 b |
| δ13C (‰) | −7.9 ± 0.1 a | −7.3 ± 0.1 b | −8.0 ± 0.2 | −8.0 ± 0.2 | −7.8 ± 0.1 a | −7.5 ± 0.0 b | −9.0 ± 0.1 | −9.9 ± 0.1 |
| δ15Ν (‰) | 1.2 ± 0.1 a | 1.0 ± 0.0A a | 1.9 ± 0.2 Aab | 2.1 ± 0.3 Abb | 1.2 ± 0.1 aa | 0.5 ± 0.0 Bba | 2.6 ± 0.1 Bab | 2.2 ± 0.0 Bbb |
| Rhizomes | ||||||||
| TC (%) | 31.2 ± 1.0 A | 32.9 ± 0.9 A | 27.7 ± 1.6 A | 29.5 ± 1.2 | 35.9 ± 0.7 Baa | 29.9 ± 2.7 Bb | 25.5 ± 1.9 Bab | 29.3 ± 0.8 b |
| TN (%) | 0.4 ± 0.1 Aa | 0.4 ± 0.1 Aa | 0.5 ± 0.1 Ab | 0.6 ± 0.0 Ab | 0.7 ± 0.0 Ba | 0.6 ± 0.1 Ba | 0.4 ± 0.0 Bb | 0.4 ± 0.1 Bb |
| C/N | 68.2 ± 5.6 Aa | 72.7 ± 6.2 Aa | 48.4 ± 2.3 Ab | 47.1 ± 3.1 Ab | 50.3 ± 1.8 Ba | 46.4 ± 0.1 Ba | 58.3 ± 1.0 Bb | 59.2 ± 1.7 Bb |
| δ13C (‰) | −8.0 ± 0.1 Aaa | −7.4 ± 0.0 Aba | −8.9 ± 0.1 Ab | −9 ± 0.1 Ab | −8.1 ± 0.2 Baa | −7.8 ± 0.1 Bba | −9.7 ± 0.1 Bab | −10.4 ± 0.0 Bbb |
| δ15Ν (‰) | −0.5 ± 0.0 Aaa | 0.6 ± 0.0 Ab | 0.5 ± 0.4 Ab | 0.5 ± 0.3 A | −0.4 ± 0.1 Ba | −0.5 ± 0.3 Ba | 0.8 ± 0.1 Bab | 0.9 ± 0.0 Bbb |
| Roots | ||||||||
| TC (%) | 33.8 ± 2.6 a | 25.2 ± 1.3 | 32.8 ± 0.61 ab | 30.8 ± 2.3 b | 33.5 ± 0.3 a | 30 ± 4.2 | 25.6 ± 2.6 b | 28 ± 1.2 |
| TN (%) | 0.6 ± 0.0 | 0.5 ± 0.0 Aa | 0.5 ± 0.0 a | 2.9 ± 0.5 Abb | 0.6 ± 0.0 a | 0.5 ± 0.0 Bb | 0.4 ± 0.0 a | 0.5 ± 0.0 Bb |
| C/N | 48.4 ± 2 | 47.0 ± 1.1 ba | 57.5 ± 7.9 Aa | 10.5 ± 0.9 Abb | 48.2 ± 1.2 a | 51.4 ± 2.7 ba | 55.4 ± 1.9 B | 53.9 ± 3.8 Bb |
| δ13C (‰) | −8.2 ± 0.2 a | −8.0 ± 0.0 ba | −7.6 ± 0.2 Aa | −8.4 ± 0.0 Abb | −8.1 ± 0.2 aa | −7.9 ± 0.1 ba | −9.2 ± 0.0 Bab | −9.7 ± 0.0 Bbb |
| δ15Ν (‰) | 1.1 ± 0.1 Aa | 0.7 ± 0.0 A | 0.8 ± 0.1 ab | 1.1 ± 0.0 Ab | 0.8 ± 0.0 Ba | 0.9 ± 0.0 Bba | 0.8 ± 0.0 a | 1.9 ± 0.0 Bbb |
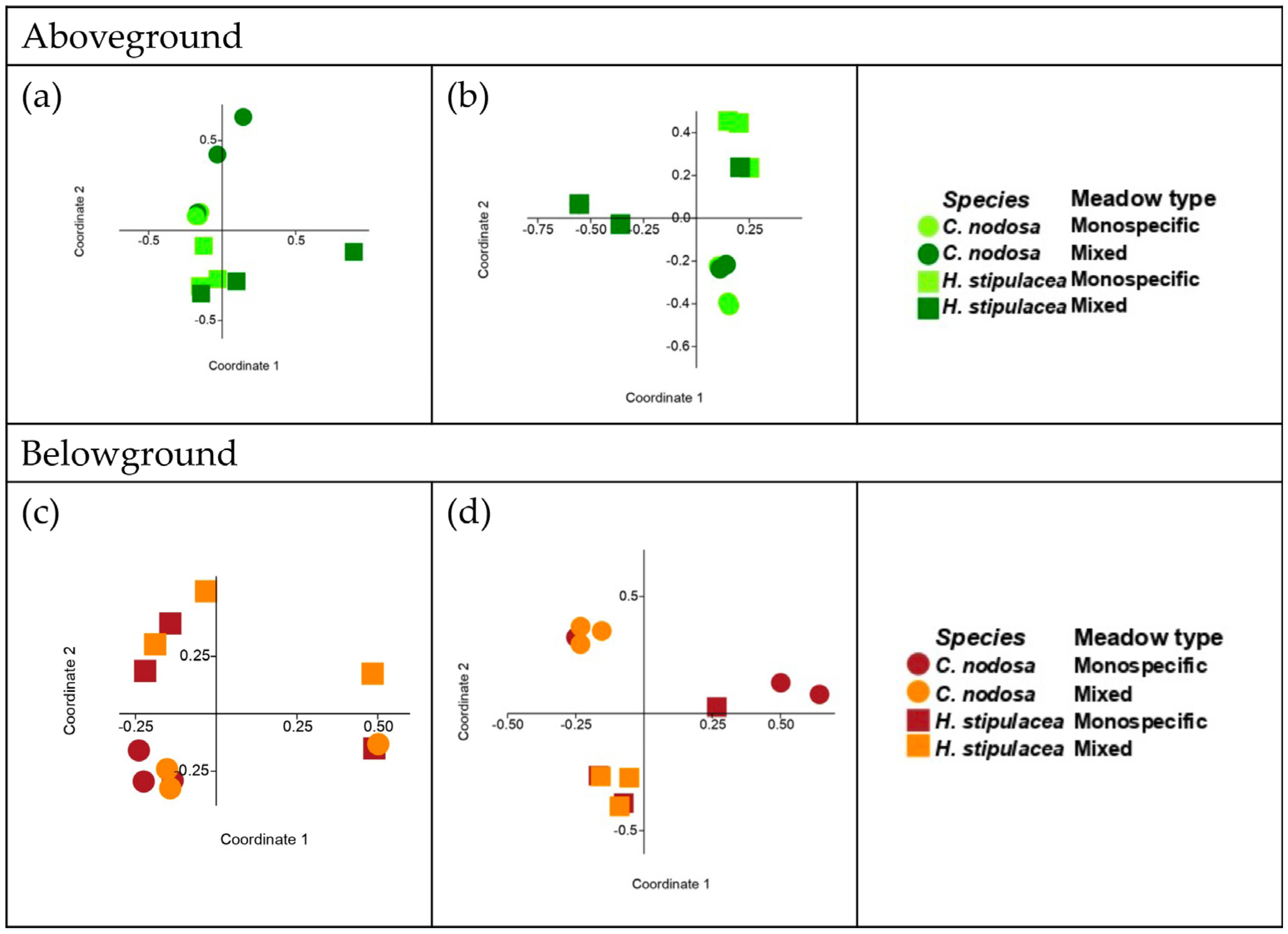
| Comparison (Two-Way ANOSIM) | |||||
|---|---|---|---|---|---|
| Site | Plant Part | Seagrass Species | Meadow Zone | ||
| R | p | R | p | ||
| Shallow | Aboveground | 0.40 | <0.01 | 0.15 | NS |
| Belowground | 0.35 | <0.01 | 0.07 | NS | |
| Deep | Aboveground | 0.27 | <0.05 | 0.22 | <0.05 |
| Belowground | 0.29 | <0.05 | 0.15 | NS | |
| Bacterial Taxon | Putative Metabolic Traits or Functions | References |
|---|---|---|
| Shallow C. nodosa sediment | ||
| Flavobacteriaceae, Pseudoalteromonadaceae | Strictly or facultative anaerobic, involved in sulfur and nitrogen cycling | [70,71] |
| Lentimicrobiaceae | Involved in sulfur cycling | [72] |
| Deep C. nodosa sediment | ||
| Thermoanaerobaculaceae | Nitrogen cycling | [73,74] |
| Vibrionaceae | Degradative bacteria, associated with seagrass decline | [75,76] |
| Alteromonadaceae | Degradative bacteria | [77,78] |
| Shallow and deep H. stipulacea sediment | ||
| No host-specific associations | ||
| Shallow and deep mixed sediment | ||
| Fusobacteriaceae, Izemoplasmataceae | Extracellular DNA degradation | [79,80] |
| Bacterial Taxon | Putative Metabolic Traits and Functions | References |
|---|---|---|
| Shallow C. nodosa aboveground—monospecific | ||
| Micavibrionaceae | Predators of other microbes, including pathogens | [81] |
| Yersiniaceae | Plant growth promoters | [82,83] |
| Xenococcaceae, Synechococcales_Incertae_Sedis | Microbes involved in nitrogen cycle | [84] |
| Parvularculaceae, Oceanospirillales | Halotolerant bacteria associated with other marine organisms | [85,86] |
| Shallow C. nodosa aboveground—mixed | ||
| Flavobacteraceae, Moraxellaceae | Complex carbon compounds (including phenols) degraders | [87,88] |
| Flavobacteraceae | Producer of compound against dinoflagellates | [89] |
| Moraxellaceae, Pirellulaceae (Planctomycetes) | Biosurfactant producers | [90,91,92] |
| Phormidesmiaceae (Cyanobacteria) | Diazotrophs | [92] |
| Shallow H. stipulacea aboveground—monospecific | ||
| Phormidiaceae, Microcystaceae | Nitrogen fixing Cyanobacteria | [93] |
| Stappiaceae | CO2 fixing and nitrifier bacteria | [94] |
| Shallow H. stipulacea aboveground—mixed | ||
| Propionibacteriaceae (Actinobacteria) | Antibacterial compounds producers | [95] |
| Deep C. nodosa aboveground—monospecific | ||
| Rhodobacteraceae | Biofilm producers | [96,97] |
| Nostocaceae (Cyanobacteria) | Nitrogen fixing bacteria | [59,85,98] |
| Methylophilaceae | Methanol degraders | [99] |
| Saprospiraceae | Complex carbon compounds degraders | [100] |
| Deep C. nodosa aboveground—mixed | ||
| Bacteroidetes | Biofilm producers found in seagrass roots | [101] |
| Marinibiliaceae | Heterotrophic fermentative metabolism | [102] |
| Pseudoalteromonadaceae, Myxococcaceae | Anti-bacterial, bacteriolytic and antiviral producers | [103,104] |
| Colwelliaceae | Nitrogen fixing bacteria | [105] |
| Thioalkalispiraceae | Sulfur oxidizers | [106] |
| Hyphomonadaceae | Nitrate reducing | [107] |
| Deep H. stipulacea aboveground—mixed | ||
| Microcystaceae, Nisaeaceae, Phormidiaceae (Cyanobacteria) | Nitrogen fixing bacteria | [10,94] |
| Methyloligellaceae | Methylotrophs Nitrogen fixer | [108] |
| Desulfocapsacaceae | Sulfur compound reducers | [109] |
| Pirellulaceae | Leaves colonizers | [90] |
| Shallow C. nodosa belowground—monospecific | ||
| Cellvibrionaceae | Nitrogen fixer, growth promoter, lignocellulose degrader | [110,111] |
| Methylophagaceae | Plant hormones producers, methanol degraders | [112,113,114,115,116] |
| Bacteroidia | Root associated bacteria | |
| Shallow C. nodosa belowground—mixed | ||
| Saccharospirillaceae | Lignocellulose degraders | [117] |
| Marinibilaceae | Root associated, complex organic materials degraders | [114] |
| Desulfosarcina | Sulfur compound reducers, root associated bacteria | [22,111] |
| Sedimenticolaceae | Sulfur-oxidizing bacteria | [115,113] |
| Bacteroidetes_BD2-2 | Root associated bacteria | [116] |
| Shallow H. stipulacea belowground—monospecific | ||
| Nodosilinaceae | Nitrogen fixing bacteria | [108] |
| Shallow H. stipulacea belowground—mixed | ||
| No association | ||
| Deep C. nodosa belowground—monospecific | ||
| Saccharospirillaceae | Lignocellulose degraders | [117] |
| Sphingomonadaceae | Aerobic chemoheterotrophs | [112] |
| Caulobacteraceae, Alteromonadaceae, Flavobacteraceae | Facultative anaerobic degraders | [78,88,118] |
| Xanthomonadaceae | Potential plant pathogens | [119] |
| Rhizobiaceae, Bacteroidetes_BD2-2, Spirochaetaceae, | Root associated bacteria | [120] |
| Burkholderiaceae | [116] | |
| Flavobacteraceae | Rhizosphere microbes of successfully restored seagrass | [113] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conte, C.; Apostolaki, E.T.; Vizzini, S.; Migliore, L. A Tight Interaction between the Native Seagrass Cymodocea nodosa and the Exotic Halophila stipulacea in the Aegean Sea Highlights Seagrass Holobiont Variations. Plants 2023, 12, 350. https://doi.org/10.3390/plants12020350
Conte C, Apostolaki ET, Vizzini S, Migliore L. A Tight Interaction between the Native Seagrass Cymodocea nodosa and the Exotic Halophila stipulacea in the Aegean Sea Highlights Seagrass Holobiont Variations. Plants. 2023; 12(2):350. https://doi.org/10.3390/plants12020350
Chicago/Turabian StyleConte, Chiara, Eugenia T. Apostolaki, Salvatrice Vizzini, and Luciana Migliore. 2023. "A Tight Interaction between the Native Seagrass Cymodocea nodosa and the Exotic Halophila stipulacea in the Aegean Sea Highlights Seagrass Holobiont Variations" Plants 12, no. 2: 350. https://doi.org/10.3390/plants12020350