1. Introduction
Remote sensing of nutrient disorders enables growers to rapidly scout their crops and manage fertilization practices with greater precision. Automation using spectral sensors mounted to trucks, tractor spray booms, or unmanned aerial vehicles (UAVs) is becoming more common in agricultural production settings [
1,
2]. Spectral sensors measure crop reflectance at various wavelengths in the electromagnetic spectrum, which can be correlated to the symptoms of specific nutrient disorders. Plants exposed to various biotic and abiotic stressors exhibit different visual symptoms, due in part to changes in how light interacts with plant tissues [
1,
3]. This characteristic enables spectral sensors to diagnose plant health problems before symptoms are visually apparent [
3]. For instance, Osborne et al. [
4] found several wavelengths in the visible and near-infrared (NIR) spectra that could be used to estimate nitrogen (N) content in field-grown corn measured under ambient light conditions. Zhang et al. [
5] reported several wavelengths that were correlated with N, phosphorus (P), or potassium (K) deficiencies in field-grown rapeseed (
Brassica napus L.) measured under artificial lighting within a laboratory setting. These studies demonstrate that spectral signatures can be correlated to specific nutrient disorders and can then be used to distinguish among disorders.
Certain wavelengths are already associated with plant stress in general and nutrient stress specifically. Many of these bands fall within the red edge, the portion of the spectrum between the visible red and NIR spectra. These wavelengths are located at approximately 700, 720, and 740 nm [
6]. Other wavelengths of interest fall in the blue, (375, 466, and 490 nm), green (515, 520, 525, 550, and 575 nm) and red (675 and 682 nm) spectra [
6]. These wavelengths have been used to classify nutrient stress or pigment changes [
6]. Soil-Plant Analysis Development (SPAD) meters emit targeted electromagnetic radiation (EMR) near 650 and 940 nm and subsequently measure the amount of radiation transmitted through the leaf [
7]. The values obtained from these sensors provide a measure of the green (high values) or yellow (low values) coloration exhibited in the foliage [
7]. These values correlate to leaf chlorophyll concentrations which in turn correlate to leaf N concentrations. The primary goals of plant spectral analysis in terms of plant nutrition are the detection and discrimination of nutrient disorders or estimating foliar nutrient concentrations.
Rustioni et al. [
8] reported successful discrimination among N, K, magnesium (Mg), and iron (Fe) deficiencies in greenhouse-grown grape (
Vitis vinifera L.) leaves using hyperspectral reflectance measurements under artificial lighting. The effects of each deficiency on foliar pigment concentrations leads to the observed spectral differences [
8]. Other studies by Adams et al. [
9,
10] demonstrate spectral separability of various micronutrient deficiencies. They found that copper (Cu)-deficient soybean [
Glycine max (L.) Merr.] leaves from growth-chamber-grown plants were most successfully distinguishable, and manganese (Mn)-deficient leaves were also distinguishable in many cases; however, Fe and zinc (Zn) deficiencies were more difficult to classify [
10]. Adams et al. [
9] suggested that Cu, Fe, and Mn may affect spectral reflectance due to their specific roles in chlorophyll synthesis and electron transport.
Although there are at least 17 essential elements required to complete the cycle of plant growth and maturation, monitoring macronutrient status is considered most important for maintaining crop quality and yield. The essential macronutrients including N, P, K, calcium (Ca), Mg, and sulfur (S) are needed in relatively high concentrations [
11]. Each of the essential nutrients perform crucial roles in plant development, and deficiencies of each nutrient cause unique symptoms that are visually diagnosable in many instances. Some of the primary effects of nutrient deficiencies include stunting and changes in leaf shape or orientation [
12]. Nutrient deficiencies often cause changes in foliar coloration which may appear yellow (chlorotic), white (bleached), brown (necrotic), red, or black [
12]. It is important to consider that the mobility of nutrients within plant tissues also dictate the location symptoms occur. For instance, deficiencies of mobile nutrients such as N, P, K, and Mg typically lead to symptom development on the mature lower foliage while relatively immobile nutrients such as Ca and S cause symptoms to develop on the upper foliage. Additionally, the pattern and distribution of symptoms on a single leaf are important to consider when diagnosing symptoms.
Henry et al. [
13,
14] induced nutrient disorders of flue-cured tobacco and provided detailed descriptions of the symptoms exhibited from each deficiency and toxicity. The results indicated that several nutrient disorders were readily induced, and symptoms had unique characteristics that made them visually distinguishable. Nitrogen deficiency appeared as a pale-yellow chlorosis that occurred uniformly on individual leaves but was more severe on the lower leaves [
14]. Similarly, S deficiency led to symptoms of pale chlorosis; however, these symptoms occurred uniformly throughout the canopy [
13]. Phosphorus deficiency resulted in symptoms of nonuniform chlorosis on the lower leaves with olive-green leaf spots and large necrotic lesions [
13]. Potassium deficiency led to the development of marginal chlorosis on the lower to central leaves, while Mg deficiency caused symptoms of interveinal chlorosis in the same region of the plant [
13].
Although numerous studies have been done to measure the spectral response of various nutrient deficiencies, fewer studies have compared the spectral responses of multiple nutrient deficiencies in one particular species. Besides nutrient status, numerous factors including plant species and maturity are known to impact spectral reflectance. Research on the effect of plant and leaf maturity on the spectral profile of healthy and nutrient deficient plants is limited. Studies investigating spectral reflectance in relation to foliar N concentrations in flue-cured tobacco have been conducted [
15], but studies investigating multiple nutrient deficiencies in flue-cured tobacco have yet to be reported. Spectral remote sensing has significant potential to aid in modern crop production, especially for high-value crops such as flue-cured tobacco. Hyperspectral remote sensing can be used to diagnose plant nutrient status but is currently impractical to use on a commercial scale due to the excessive quantities of data and computational processing requirements. Using hyperspectral data to identify relevant wavelengths associated with particular nutrient deficiencies can lead to the development of relatively inexpensive sensors, offering greater opportunities for commercial utilization.
The purpose of this study was to determine if macronutrient deficiency symptoms could be distinguished among one another using spectral reflectance data. We hypothesized that discrimination of certain nutrient deficiencies would be more successful than others. In particular, we believed the symptoms of N and S deficiencies would be difficult to uncouple due to their visual similarities. This study was achieved by inducing N, P, K, Mg, and S deficiencies in a controlled environment and recording the spectral reflectance from each disorder at different stages of plant maturity.
3. Discussion
Li et al. [
18] studied hyperspectral reflectance for estimating N in upper, middle, and lower leaves of oilseed rape plants. They used partial least squares regression (PLSR) and reported optimal wavelengths of 437, 565, 667, 724, 993, 1084, and 1189 nm for upper leaves; 423, 570, 598, 659, 725, and 877 nm for middle leaves; and 420, 573, 597, 667, and 718 nm for lower leaves [
18]. Li et al. [
19] also used PLSR to model and predict foliar N and P concentrations in oilseed rape. They compared PLSR with other data transformations, such as first derivatives and continuum removal, and found that PLSR with first derivatives was most effective for nutrient estimation. The optimal wavelengths selected for N were 445, 556, 657, 764, 985, 1082, and 1994 nm while those selected for P included 755, 832, 891, 999, 1196, and 1267 nm [
19]. Zhang et al. [
5] investigated the spectral response of oilseed rape to N, P, and K deficiencies using PLSR. They reported optimal wavelengths of 440, 473, 513, 542, 659, 718, 744, 865, 928, 965, 986, and 1015 nm for N; 468, 522, 698, 721, 817, 967, 979, and 1025 nm for P; and 456, 554, 667, 720, and 1027 nm for K [
5].
These past studies demonstrate similar results to what we obtained in the present study. In general, the majority of observed variation appears to be in the visible spectrum and falls specifically between 400 and 750 nm. However, other studies reported several significant wavelengths >800 nm [
5,
18,
19], which was not the case in our study. Furthermore, we identified bands of interest in the UV region, whereas most other studies did not. This may be due in part to the specific spectroradiometer used in this study. It is also likely that the species and nutrient deficiencies used in this study were different from the other studies, and plants are known to demonstrate different spectral responses by species [
6] and in response to different stimuli. Further work should investigate whether UV EMR is a good indicator of nutrient deficiencies in species other than tobacco.
Overall, PCA-based classification was highly accurate in distinguishing among the five macronutrient deficiencies induced in this study. Accuracy decreased with increasing maturity, likely due to the greater proportion of asymptomatic leaves on these plants. This proposed effect was especially impactful for the discrimination of P and K deficiencies. Nitrogen and S-deficient leaves were hypothesized to be similar enough to prevent accurate classification between these symptoms. However, in only three instances were S-deficient plants misclassified as N-deficient, and in no instance was a N-deficient plant misclassified as S-deficient. Phosphorus-deficient plants exhibited some of the lowest overall classification accuracies which may be attributed to the non-uniform appearance of the leaf surface. Similarly, mature K-deficient plants had poor classification accuracy, possibly because a large portion of the upper leaves appeared completely asymptomatic, and those leaves remained in the analysis. Therefore, it may be inferred that N-, Mg-, and S-deficient plants have a greater proportion of leaves that are visibly and spectrally different from the leaves of control plants. Future work may find that dividing leaves by visual symptoms could yield helpful insights.
4. Materials and Methods
4.1. Plant Material and Experimental Design
The experiment was replicated twice over time, and a total of 2563 individual sample readings were taken over the 2 trials. Each replication began by sowing pelletized K-326 tobacco seeds (GoldLeaf Seed Co., Hartsville, SC, USA) into 128-cell plug flats and placing them in a glass-glazed greenhouse at North Carolina State University in Raleigh, NC (35°47′41″ N lat, 78°41′57″ W long). The substrate was an 80:20 (v:v) mix of Canadian sphagnum peat moss (Conrad Fafard, Agawam, MA, USA) and horticultural coarse perlite (Perlite Vermiculite Packaging Industries, Inc., North Bloomfield, OH, USA), amended with mesh size #100 dolomitic limestone (Rockydale Agricultural, Roanoke, VA, USA) at 8.9 kg m−3 and wetting agent (AquaGro 2000 G; Aquatrols, Cherry Hill, NJ, USA) at 0.6 kg m−3. This custom substrate was used to limit nutrient contamination that would be present in a commercial substrate. Flats were irrigated by hand with nonfertilized water until seeds germinated and cotyledons fully expanded.
Following germination, seedlings were fertilized with a solution consisting of 7.5 mmol L
−1 N, 0.5 mmol L
−1 P, 3.0 mmol L
−1 K, and 2.5 mmol L
−1 Ca. Previous experiments demonstrated tobacco seedlings require a low concentration of primary macronutrients to develop sufficiently for studies investigating nutrient disorders [
13,
14]. The selected concentrations were a half rate of the primary macronutrient concentrations used in the nutrient-sufficient control solution. The control solution was a modified all-nitrate Hoagland solution consisting of 15.0 mmol L
−1 N, 1.0 mmol L
−1 P, 6.0 mmol L
−1 K, 5.0 mmol L
−1 Ca, 2.0 mmol L
−1 Mg, and 2.0 mmol L
−1 S, 72.0 µmol L
−1 iron (Fe), 18.0 µmol L
−1 Mn, 3.0 µmol L
−1 Cu, 3.0 µmol L
−1 Zn, 45.0 µmol L
−1 boron (B), and 0.1 µmol L
−1 molybdenum (Mo) [
20] mixed with deionized (DI) water of 18 megohm purity. All nutrients were provided by the following technical grade salts (Fisher Scientific, Pittsburg, PA, USA): calcium nitrate tetrahydrate [Ca(NO
3)
2·4H
2O], potassium nitrate (KNO
3), potassium dihydrogen phosphate (KH
2PO
4), potassium sulfate (K
2SO
4), magnesium sulfate heptahydrate (MgSO
4·7H
2O), potassium chloride (KCl), calcium chloride dihydrate (CaCl
2·2H
2O), sodium nitrate (NaNO
3), sodium phosphate dihydrate (NaH
2PO
4·2H
2O), iron chelate (Fe-DTPA), manganese chloride tetrahydrate (MnCl
2·4H
2O), zinc chloride heptahydrate (ZnCl
2·7H
2O), copper chloride dihydrate CuCl
2·2H
2O, boric acid (H
3BO
3), and sodium molybdate dihydrate (Na
2MoO
4·2H
2O). Solution pH was adjusted to ~6.0 using sodium hydroxide (NaOH). Macronutrient deficiencies were induced by replacing each cation (K
+ and Mg
+) or anion [(nitrate (NO
3−), phosphate (H
2PO
4−), and sulfate (SO
42−)] containing salt with a sodium (Na) or chloride (Cl
−) containing salt, respectively. Fertilizer treatments began upon transplanting into a sand culture system.
The seedlings were thoroughly drenched with DI water three consecutive times to leach any remaining nutrients prior to transplant. Seedlings were transplanted into 12.4 cm diameter pots (Dillen, Middlefield, OH, USA) filled with silica sand [Millersville #2 (0.8 to 1.2 mm diameter); Southern Products & Silica Co., Hoffman, NC, USA] that was soaked in sulfuric acid and triple-rinsed with DI water prior to use. The transplanted seedlings were placed into an automated recirculating sand culture system. The system was built on benches in a glass-glazed greenhouse in Raleigh, NC and utilized a completely randomized design. Each segment of the system (referred to as “lines” hereafter) consisted of 10.2 cm diameter polyvinyl chloride (PVC) piping (Charlotte Plastics, Charlotte, NC, USA) fit with 12.7 cm diameter PVC reducer couplings (Charlotte Plastics). Six couplings were used to hold pots and recapture irrigation solutions in each line. Fertilizer solutions were delivered via drip tubes fed from individual 20 L plastic buckets equipped with submersible pumps (model 1A; Little Giant Pump Co., Oklahoma City, OK, USA). Each line contained one treatment. Solutions were delivered for 1 min each cycle with as many cycles necessary per day to prevent apparent water stress (i.e., wilting) between 6:00 and 18:00 h. Nutrient solutions were replaced on a weekly basis.
4.2. Spectroscopic Measurements
A handheld spectrometer (PSM-2500; Spectral Evolution, Lawrence, MA, USA) was used to collect spectral reflectance data throughout the study. The sensor had a spectral range of 300 to 2500 nm and a spectral resolution of 3.5 nm at 700 nm, 22 nm at 1500 nm, and 22 nm at 2100 nm with the percent reflectance output in 1 nm increments. The sensor was equipped with a fiberoptic leaf clip capable of taking leaf-level measurements. This leaf clip had a self-contained light source with two light settings, of which the higher setting was used. Leaf clip measurements were taken inside the greenhouse under ambient light conditions. Measured plants were irrigated prior to measurement to ensure drought stress would not be a confounding factor.
Throughout the experiment, symptomatic and asymptomatic control plants were selected to obtain hyperspectral measurements of the individual leaves. At each measurement date, four individual plant replicates were selected. Reflectance data were collected from each leaf, except for the small bottom two to three leaves exhibiting natural senescence and the few immature upper leaves that were narrow with limited expansion. Measurements using the leaf clip were taken approximately 2 to 3 cm from the leaf margin and one-third of the leaf length away from the leaf tip. This location was selected because it typically represented the average degree of symptoms for each leaf. Areas of necrosis were avoided as necrotic tissues reflected much different patterns than non-necrotic tissues, regardless of symptomology.
4.3. Leaf Tissue Analysis
Leaf tissues were dried at 70 °C for 72 h and then ground in a sample mill (Thomas Wiley® Mini-Mill; Thomas Scientific, Swedesboro, NJ, USA), and analyzed for nutrient concentrations (AgSource Laboratories, Lincoln, NE, USA). Total N was processed by Kjeldahl digestion and determined via flow injection analysis (FIA). Extractable K was processed by 2% acetic acid digestion and determined via inductively coupled plasma mass spectrometry (ICP-MS). Total P and all other plant minerals were processed by nitric acid/hydrogen peroxide digestion and determined via ICP-MS. Pairwise differences between control and deficient plant foliar nutrient concentrations were determined using PROC ANOVA in SAS (version 9.4; SAS Institute, Cary, NC, USA).
4.4. Data Preparation
Reflectance data were manipulated and analyzed using R statistical software (R Core Team, Auckland, New Zealand). Measurements from each leaf of each treatment were assigned into several subgroups depending on various characteristics. Leaves received an objective symptom severity rating of four possible categories: (1) none, (2) low, (3) intermediate, or (4) high (
Figure 5). We divided the symptom severity into four categories so that we could compare statistics for leaves that were visually symptomatic versus those that were not. Categorization did not affect the results of the study but allowed us to refine and better understand the symptoms that were observed. Examples of leaves from each category can be seen in
Figure 5. Furthermore, measurement stages were separated by maturity: (1) young, (2) intermediate, or (3) mature. Young plants were those that had 6 or fewer leaves, intermediate plants had 7 to 12 leaves, and mature plants had more than 12 leaves. Fully mature tobacco plants developed between 18 and 20 expanded leaves prior to anthesis.
Water absorption bands were excluded from the analysis in the ranges of 1355 to 1450 nm and 1800 to 1950 nm. These wavelengths in the short-wave infrared (SWIR) region can be important in the detection of water related stress, but can also contribute noise, leading to the distortion of spectral measurements used for the detection of nutrient disorders [
1].
4.5. Band Selection
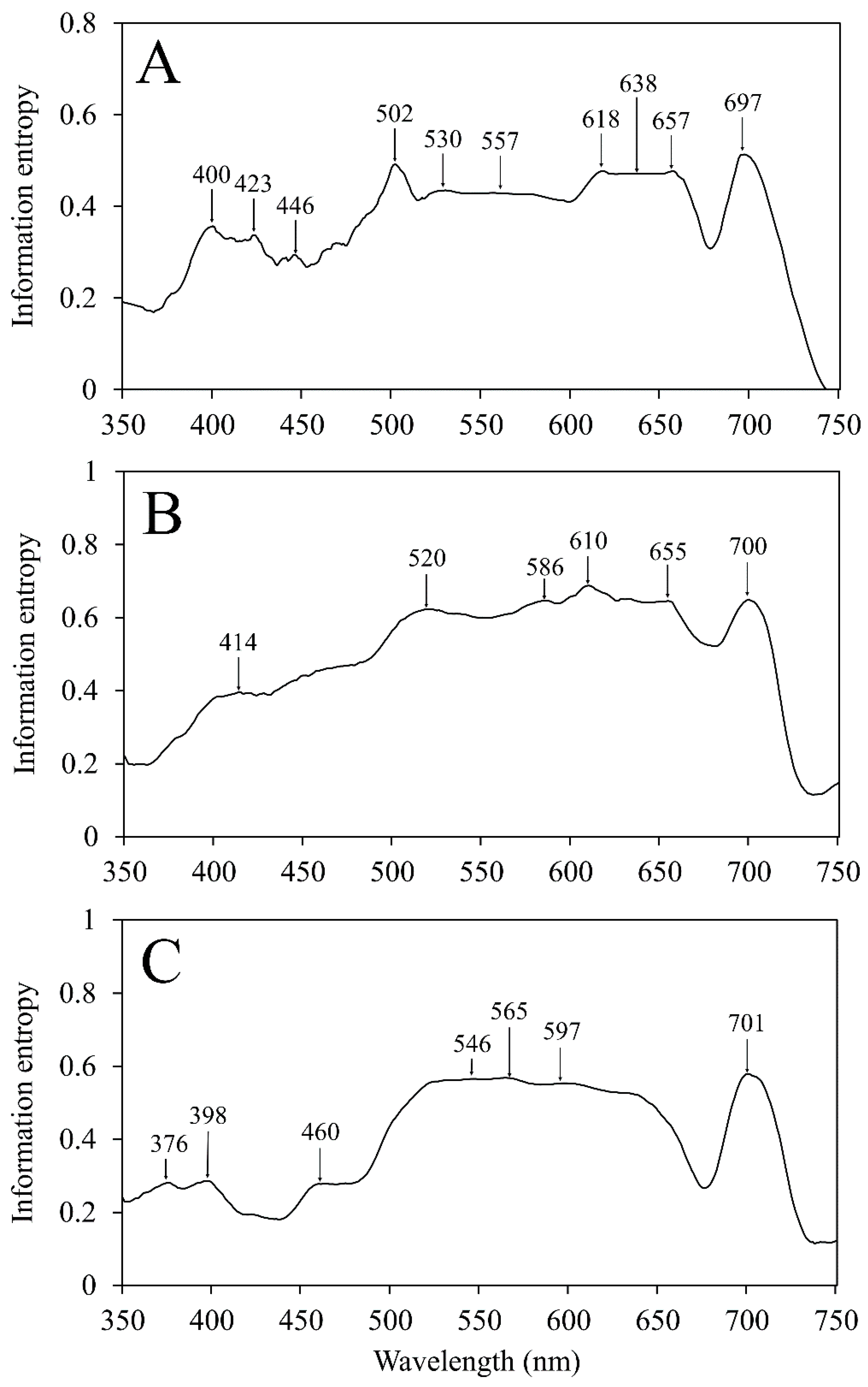
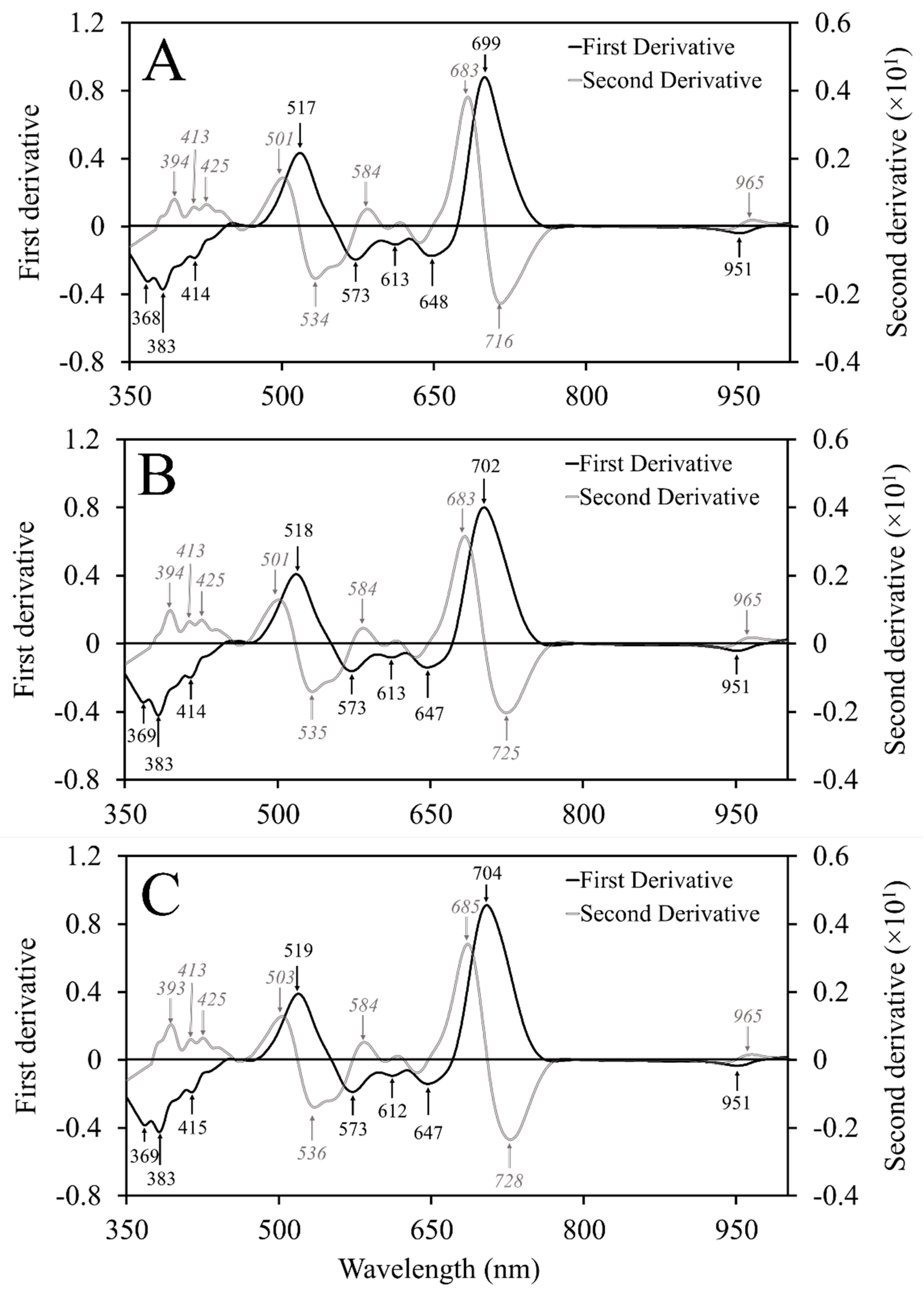
Spectra were used to calculate information entropy as well as first and second spectral derivatives to identify which wavelengths contribute most to the variability observed among the nutrient-deficient leaves. Information entropy is a measurement for how much variability is contained within a band and it is commonly used for hyperspectral band selection [
16,
17,
21]. Derivative measures are also useful for band selection by identifying areas with rapid changes in direction or pattern within spectra [
16]. Information entropy was calculated using the FSelector package [
22] while derivatives were calculated using the prospectr package [
23] in R. Savitzky–Golay filtering was applied using a third-order polynomial with a smoothing window size of 10 bands to information entropy data to reduce noise and smooth the data. Derivatives were calculated and smoothed using the gap-segment algorithm with a filter length of 11 bands for both first and second order derivatives. Peaks and valleys were then calculated using the splus2R package [
24] to determine the greatest individual absolute values or ranges of values within a span of 10 bands. Ranges of continuous values with uniform importance were aggregated and used to determine the central wavelength representing that range. The identified wavelengths represented those with the greatest impact on observed variability.
4.6. Symptom Classification
Reflectance spectra were analyzed by growth stage using PCA, with 10 PCs selected for each growth stage. Linear discriminant analysis was used to develop classification models to distinguish among the various symptoms. The classification models were conducted using a (custom-built) five-fold cross validation approach, where 80% of the data was randomly selected for model training and the remaining 20% used for testing. Following the first validation, a second validation was conducted with a new set of testing data previously used to train the model. This process was repeated to optimize the model until all data were used for both training and testing functions. The final classification results were put into a confusion matrix for each growth stage and analyzed using the PredPsych package [
25] in R. The matrices were used to determine the overall accuracy and misclassification rates for each model. Overall accuracy was calculated by dividing the number of correctly classified observations by the total number of observations.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}