
Novel Tetraploid Triticale (Einkorn Wheat × Rye)—A Source of Stem Rust Resistance

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
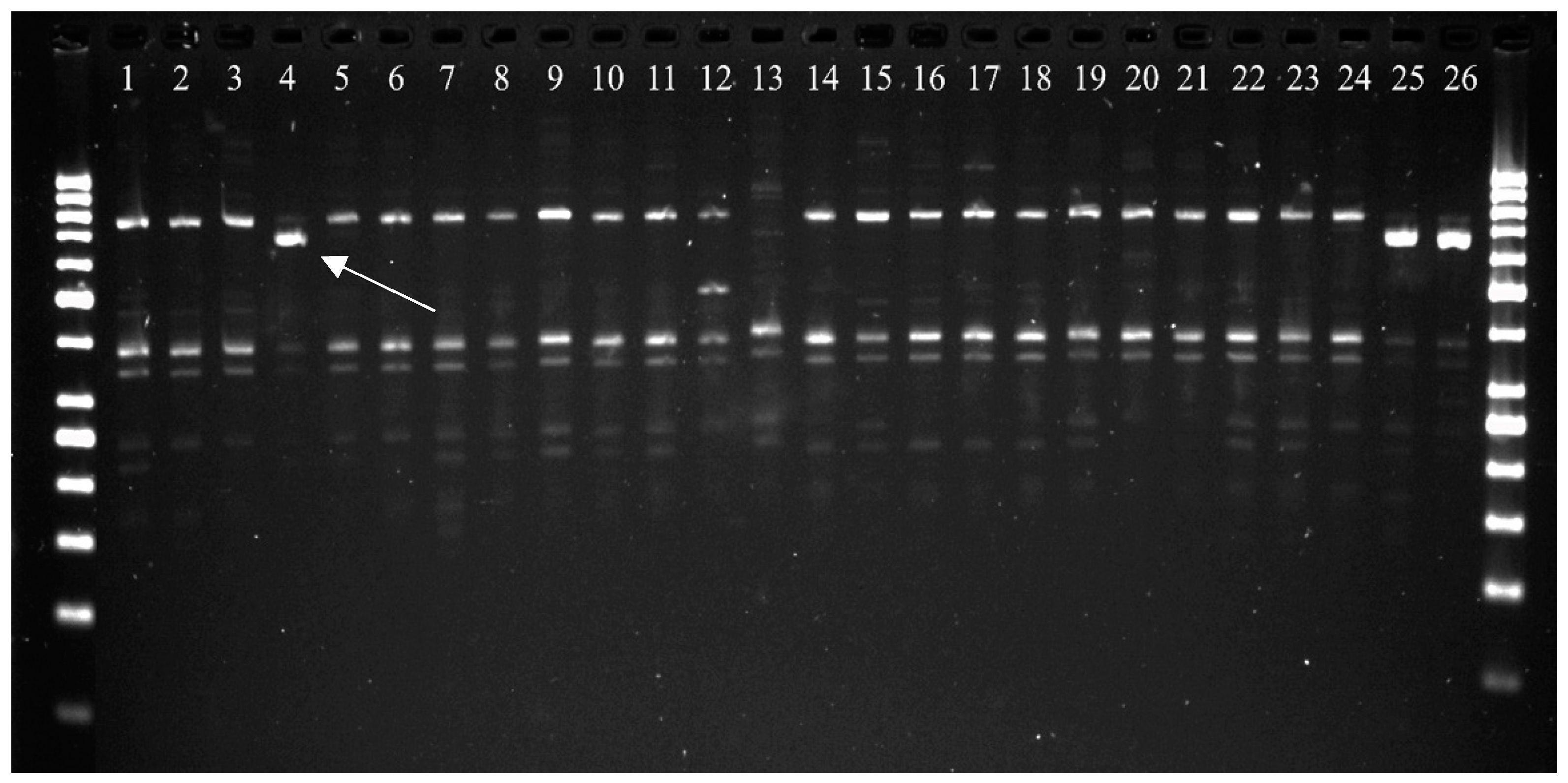
2.1. Identification of Molecular Markers Linked to Sr22 and Sr35 loci
2.2. Cross-Hybridizations and Self-Pollinations
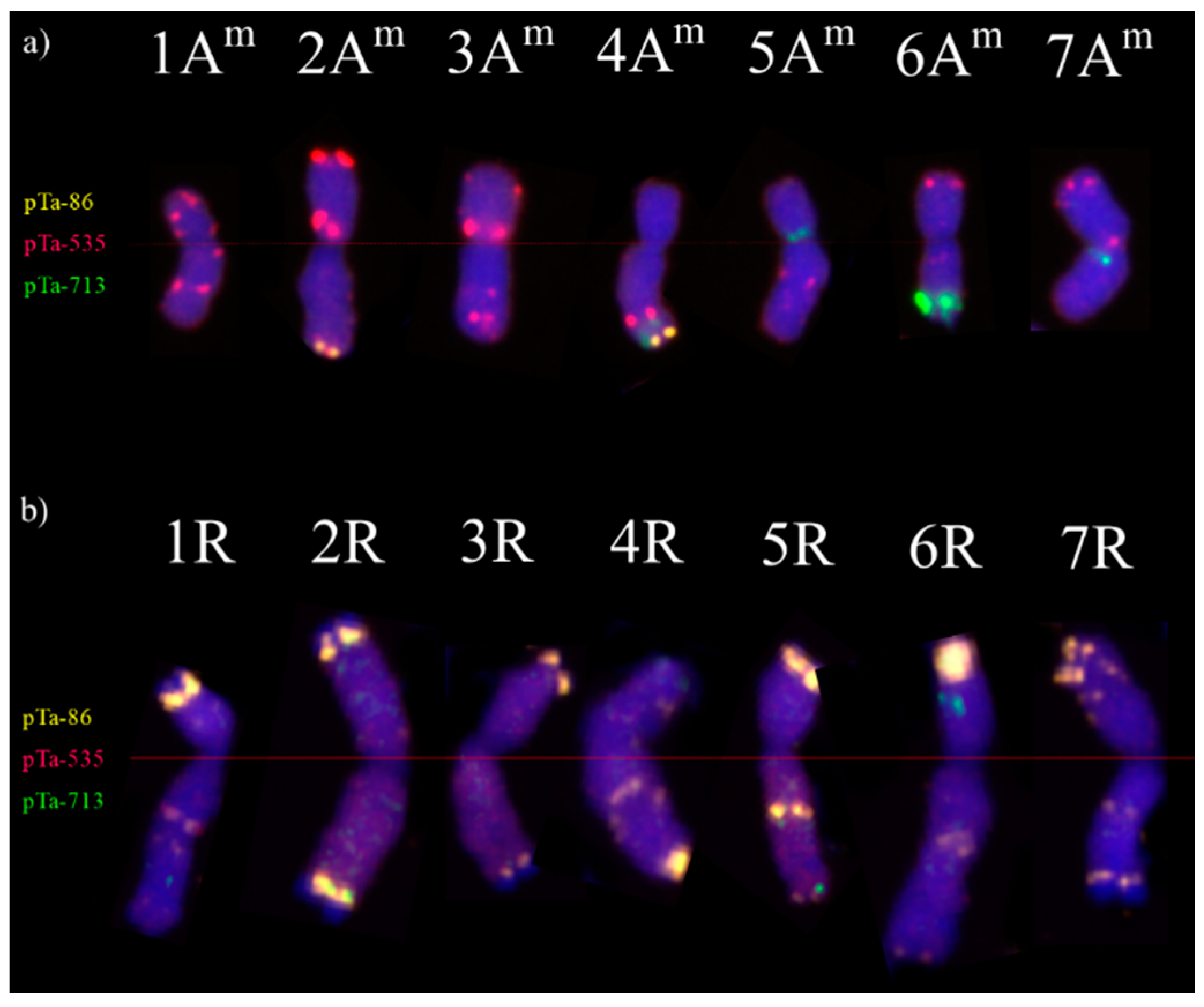
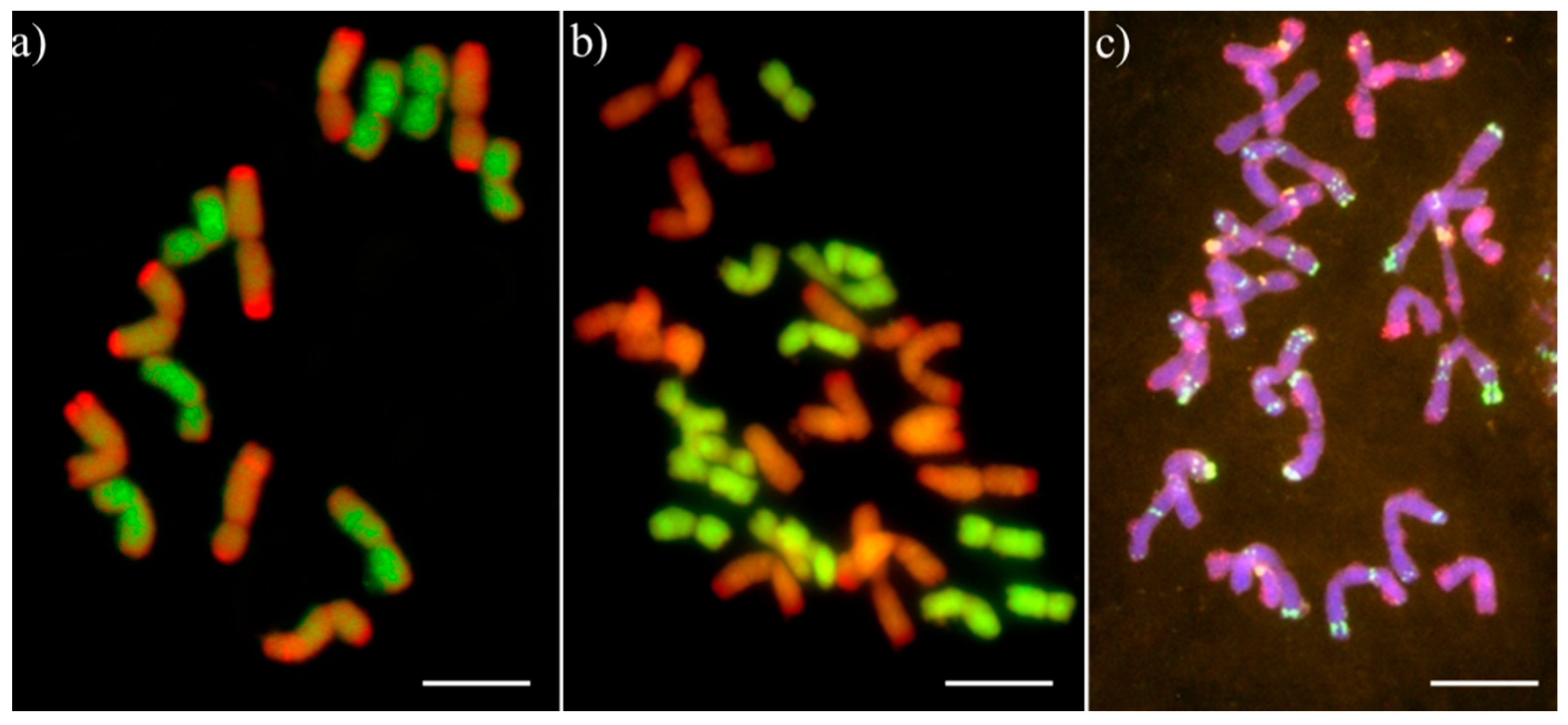
2.3. Karyotyping
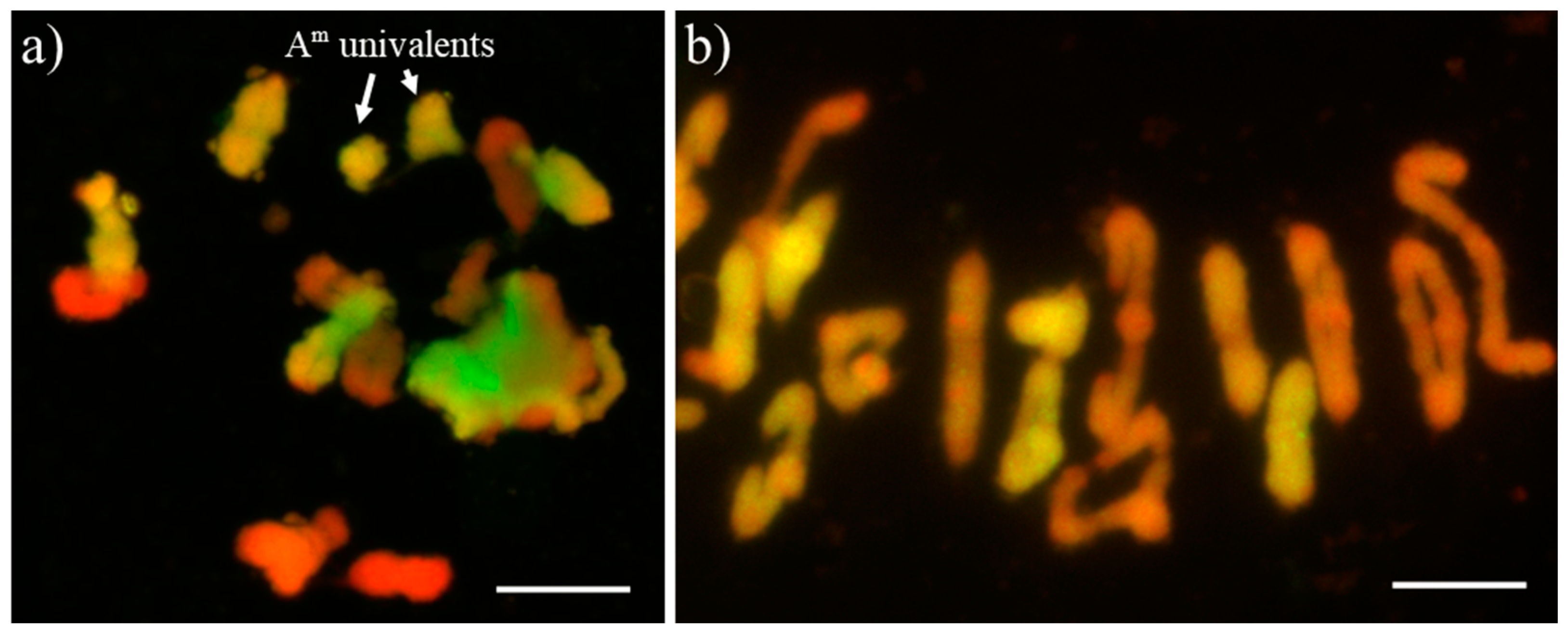
2.4. Meiosis Analysis in Pollen Mother Cells
2.5. Ptg Infection Tests
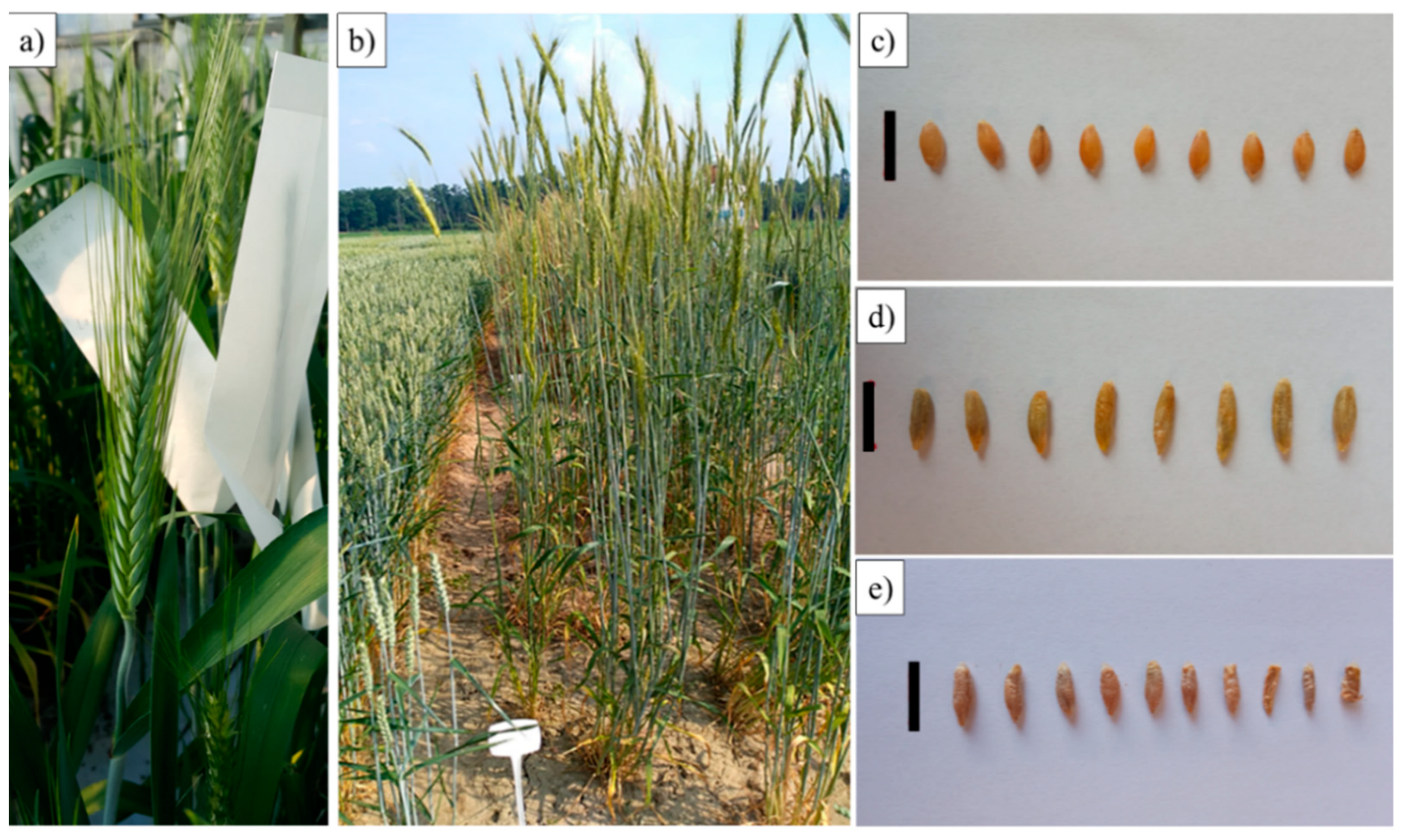
2.6. Evaluation of Yield Related Traits in Progeny of S3 Generation Plants
3. Discussion
4. Methods
4.1. Plant Material
4.2. Cross-Hybridizations
4.3. Identification of NL9F5 Molecular Markers Linked to Sr35 Loci
4.4. Preparation of Chromosome Spreads
4.5. Karyotyping Using Fluorescence In Situ Hybridization (FISH)
4.6. Genomic In Situ Hybridization (GISH)
4.7. Ptg Infection Test
4.8. Evaluation of Yield Related Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanaei, S.; Stuart, P. Systematic Assessment of Triticale-Based Biorefinery Strategies: Techno-Economic Analysis to Identify Investment Opportunities. Biofuels Bioprod. Biorefin. 2018, 12, S46–S59. [Google Scholar] [CrossRef]
- OECD/FAO. OECD-FAO Agricultural Outlook 2018–2027; OECD/FAO: Rome, Italy, 2018; ISBN 978-92-64-06203-0. [Google Scholar]
- Mupondwa, E.; Li, X.; Tabil, L. Integrated Bioethanol Production from Triticale Grain and Lignocellulosic Straw in Western Canada. Ind. Crops Prod. 2018, 117, 75–87. [Google Scholar] [CrossRef]
- Pretorius, Z.A.; Jin, Y.; Bender, C.M.; Herselman, L.; Prins, R. Seedling Resistance to Stem Rust Race Ug99 and Marker Analysis for Sr2, Sr24 and Sr31 in South African Wheat Cultivars and Lines. Euphytica 2012, 186, 15–23. [Google Scholar] [CrossRef]
- Arseniuk, E. Triticale Diseases—A Review. In Triticale: Today and Tomorrow; Springer: Berlin/Heidelberg, Germany, 1996; pp. 499–525. ISBN 978-94-010-6634-1. [Google Scholar]
- Zhang, J.; Wellings, C.R.; McIntosh, R.A.; Park, R.F.; Zhang, J.; Wellings, C.R.; McIntosh, R.A.; Park, R.F. Seedling Resistances to Rust Diseases in International Triticale Germplasm. Crop. Pasture Sci. 2010, 61, 1036–1048. [Google Scholar] [CrossRef]
- Gruner, P.; Schmitt, A.K.; Flath, K.; Schmiedchen, B.; Eifler, J.; Gordillo, A.; Schmidt, M.; Korzun, V.; Fromme, F.J.; Siekmann, D.; et al. Mapping Stem Rust (Puccinia graminis f. sp. secalis) Resistance in Self-Fertile Winter Rye Populations. Front. Plant Sci. 2020, 11, 667. [Google Scholar] [CrossRef] [PubMed]
- Martis, M.M.; Zhou, R.; Haseneyer, G.; Schmutzer, T.; Vrána, J.; Kubaláková, M.; König, S.; Kugler, K.G.; Scholz, U.; Hackauf, B.; et al. Reticulate Evolution of the Rye Genome. Plant Cell 2013, 25, 3685–3698. [Google Scholar] [CrossRef] [Green Version]
- Boshoff, W.H.P.; Bender, C.M.; Pretorius, Z.A. Reaction of South African Rye, Triticale and Barley Forage Cultivars to Stem and Leaf Rust. S. Afr. J. Plant Soil. 2019, 36, 77–82. [Google Scholar] [CrossRef]
- Boshoff, W.H.P.; Pretorius, Z.A.; Terefe, T.G.; Bender, C.M.; Herselman, L.; Maree, G.J.; Visser, B. Phenotypic and Genotypic Description of Puccinia graminis f. sp. tritici Race 2SA55 in South Africa. Eur. J. Plant Pathol. 2018, 152, 783–789. [Google Scholar] [CrossRef]
- Leonard, K.J.; Szabo, L.J. Stem Rust of Small Grains and Grasses Caused by Puccinia graminis. Mol. Plant Pathol. 2005, 6, 99–111. [Google Scholar] [CrossRef]
- Terefe, T.G.; Visser, B.; Pretorius, Z.A. Variation in Puccinia graminis f. sp. tritici Detected on Wheat and Triticale in South Africa from 2009 to 2013. Crop Prot. 2016, 86, 9–16. [Google Scholar] [CrossRef]
- Bhavani, S.; Hodson, D.P.; Huerta-Espino, J.; Randhawa, M.S.; Singh, R.P. Progress in Breeding for Resistance to Ug99 and Other Races of the Stem Rust Fungus in CIMMYT Wheat Germplasm. Front. Agric. Sci. Eng. 2019, 6, 210–224. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Hodson, D.P.; Jin, Y.; Lagudah, E.S.; Ayliffe, M.A.; Bhavani, S.; Rouse, M.N.; Pretorius, Z.A.; Szabo, L.J.; Huerta-Espino, J.; et al. Emergence and Spread of New Races of Wheat Stem Rust Fungus: Continued Threat to Food Security and Prospects of Genetic Control. Phytopathology 2015, 105, 872–884. [Google Scholar] [CrossRef] [Green Version]
- Fetch, T.G.; Park, R.F.; Pretorius, Z.A.; Depauw, R.M. Stem Rust: Its History in Kenya and Research to Combat a Global Wheat Threat. Can. J. Plant Pathol. 2021, 43, S275–S297. [Google Scholar] [CrossRef]
- Zaharieva, M.; Monneveux, P. Cultivated Einkorn Wheat (Triticum monococcum L. subsp. monococcum): The Long Life of a Founder Crop of Agriculture. Genet Resour. Crop. Evol. 2014, 61, 677–706. [Google Scholar] [CrossRef]
- Kilian, B.; Özkan, H.; Walther, A.; Kohl, J.; Dagan, T.; Salamini, F.; Martin, W. Molecular Diversity at 18 Loci in 321 Wild and 92 Domesticate Lines Reveal No Reduction of Nucleotide Diversity during Triticum monococcum (Einkorn) Domestication: Implications for the Origin of Agriculture. Mol. Biol. Evol. 2007, 24, 2657–2668. [Google Scholar] [CrossRef] [Green Version]
- Noweiska, A.; Bobrowska, R.; Kwiatek, M.T. Structural Polymorphisms of Chromosome 3Am Containing Lr63 Leaf Rust Resistance Loci Reflect the Geographical Distribution of Triticum monococcum L. and Related Diploid Wheats. Agriculture 2022, 12, 966. [Google Scholar] [CrossRef]
- Zohary, D. Monophyletic vs. Polyphyletic Origin of the Crops on Which Agriculture Was Founded in the Near East. Genet. Resour. Crop. Evol. 1999, 46, 133–142. [Google Scholar] [CrossRef]
- Plamenov, D.; Belchev, I.; Kiryakova, V.; Spetsov, P. Fungal Resistance of Triticum Durum—T. monococcum ssp. aegilopoides Amphiploid. J. Plant Dis. Prot. 2016, 116, 60–62. [Google Scholar] [CrossRef]
- Neumann, M.; Sodkiewicz, W.; Skiebe, K. On Possibilities of Genetic Information Transfer from Triticum monococcum to Triticale. Genet. Pol. 1985, 26, 209–215. [Google Scholar]
- Sodkiewicz, W. Amphiploid Triticum monococcum L. × Secale cereale L. (AARR)—A New Form of Tetraploid Triticale. Cereal. Res. Commun. 1984, 12, 35–40. [Google Scholar]
- Sodkiewicz, W.; Strzembicka, A. Application of Triticum monococcum for the Improvement of Triticale Resistance to Leaf Rust (Puccinia Triticina). Plant Breed. 2004, 123, 39–42. [Google Scholar] [CrossRef]
- Cox, T.S.; Harrell, L.G.; Chen, P.; Gill, B.S. Reproductive Behavior of Hexaploid/Diploid Wheat Hybrids 1. Plant Breed. 1991, 107, 105–118. [Google Scholar] [CrossRef]
- Loginova, D.B.; Silkova, O.G. Mitotic Behavior of Centromeres in Meiosis as the Fertility Restoration Mechanism in Wheat-Rye Amphihaploids. Russ. J. Genet. 2014, 50, 818–827. [Google Scholar] [CrossRef]
- Tayalé, A.; Parisod, C. Natural Pathways to Polyploidy in Plants and Consequences for Genome Reorganization. Cytogenet. Genome Res. 2013, 140, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Ślusarkiewicz-Jarzina, A.; Pudelska, H.; Woźna, J.; Pniewski, T. Improved Production of Doubled Haploids of Winter and Spring Triticale Hybrids via Combination of Colchicine Treatments on Anthers and Regenerated Plants. J. Appl. Genet. 2017, 58, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Megyeri, M.; Mikó, P.; Farkas, A.; Molnár-Láng, M.; Molnár, I. Cytomolecular Discrimination of the Am Chromosomes of Triticum monococcum and the A Chromosomes of Triticum aestivum Using Microsatellite DNA Repeats. J. Appl. Genet. 2016, 58, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Komuro, S.; Endo, R.; Shikata, K.; Kato, A. Genomic and Chromosomal Distribution Patterns of Various Repeated DNA Sequences in Wheat Revealed by a Fluorescence in Situ Hybridization Procedure. Genome/Natl. Res. Counc. Can. 2013, 56, 131–137. [Google Scholar] [CrossRef]
- Kwiatek, M.T.; Banaszak, Z.; Skowrońska, R.; Kurasiak-Popowska, D.; Mikołajczyk, S.; Niemann, J.; Tomkowiak, A.; Weigt, D.; Nawracała, J. Spike Morphology Alternations in Androgenic Progeny of Hexaploid Triticale (× Triticosecale Wittmack) Caused by Nullisomy of 2R and 5R Chromosomes. Vitr. Cell. Dev. Biol. -Plant 2020, 56, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Kwiatek, M.T.; Wiśniewska, H.; Belter, J.; Ulaszewski, W.; Phillips, D. Development and Cytomolecular Identification of Monosomic Alien Addition and Substitution of 2Sk Chromosome from Aegilops Kotschyi to Triticale (×Triticosecale Wittmack). Front. Plant Sci. 2020, 1722. [Google Scholar] [CrossRef]
- Salmanowicz, B.P.; Langner, M.; Wiśniewska, H.; Apolinarska, B.; Kwiatek, M.; Błaszczyk, L. Molecular, Physicochemical and Rheological Characteristics of Introgressive Triticale/Triticum monococcum ssp. monococcum Lines with Wheat 1D/1A Chromosome Substitution. Int. J. Mol. Sci. 2013, 14, 15595–15614. [Google Scholar] [CrossRef]
- Ulaszewski, W.; Belter, J.; Wiśniewska, H.; Szymczak, J.; Skowrońska, R.; Phillips, D.; Kwiatek, M.T. Recovery of 2R.2Sk Triticale-Aegilops Kotschyi Robertsonian Chromosome Translocations. Agronomy 2019, 9, 646. [Google Scholar] [CrossRef] [Green Version]
- McClintock, B. The Significance of Responses of the Genome to Challenge. Science 1984, 226, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Comai, L. The Advantages and Disadvantages of Being Polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Levy, A.A. Genome Evolution Due to Allopolyploidization in Wheat. Genetics 2012, 192, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Rouse, M.N.; Jin, Y. Stem Rust Resistance in A-Genome Diploid Relatives of Wheat. Plant Dis. 2011, 95, 941–944. [Google Scholar] [CrossRef] [Green Version]
- Kosgey, Z.C.; Edae, E.A.; Dill-Macky, R.; Jin, Y.; Bulbula, W.D.; Gemechu, A.; Macharia, G.; Bhavani, S.; Randhawa, M.S.; Rouse, M.N. Mapping and Validation of Stem Rust Resistance Loci in Spring Wheat Line CI 14275. Front. Plant Sci. 2021, 11, 2147. [Google Scholar] [CrossRef]
- Cantale, C.; Petrazzuolo, F.; Correnti, A.; Farneti, A.; Felici, F.; Latini, A.; Galeffi, P. Triticale for Bioenergy Production. Agric. Agric. Sci. Procedia 2016, 8, 609–616. [Google Scholar] [CrossRef]
- Kwiatek, M.T.; Wiśniewska, H.; Ślusarkiewicz-Jarzina, A.; Majka, J.; Majka, M.; Belter, J.; Pudelska, H. Gametocidal Factor Transferred from Aegilops Geniculata Roth Can Be Adapted for Large-Scale Chromosome Manipulations in Cereals. Front. Plant Sci. 2017, 8, 409. [Google Scholar] [CrossRef] [Green Version]
- Kwiatek, M.; Majka, M.; Majka, J.; Belter, J.; Suchowilska, E.; Wachowska, U.; Wiwart, M.; Wiśniewska, H. Intraspecific Polymorphisms of Cytogenetic Markers Mapped on Chromosomes of Triticum polonicum L. PLoS ONE 2016, 11, e0158883. [Google Scholar] [CrossRef] [Green Version]
- Kwiatek, M.T.; Majka, J.; Majka, M.; Belter, J.; Wisniewska, H. Adaptation of the Pivotal-Differential Genome Pattern for the Induction of Intergenomic Chromosome Recombination in Hybrids of Synthetic Amphidiploids within Triticeae Tribe. Front. Plant Sci. 2017, 8, 1300. [Google Scholar] [CrossRef]
- Roelfs, A.P. Rust Diseases of Wheat: Concepts and Methods of Disease Management; Cimmyt: Veracruz, Mexico, 1992; ISBN 968612747X. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession Number | Name/Signature | Origin | PCR Products (Base Pairs) for NL9F5 (Sr35) Marker |
|---|---|---|---|---|
| Triticum monococcum L. subsp. monococcum | ||||
| 1. | PI 167627 | 3412 | Turkey | 800 |
| 2. | PI 190945 | 3962 | Portugal | 800 |
| 3. | PI 191381 | Flavescens | Ethiopia | 800 |
| 4. | PI 191383 | Laetissimum | Ethiopia | 719 |
| 5. | PI 277190 | ATRI 613/59 | no data | 800 |
| 6. | PI 286068 | no data | Poland | 800 |
| 7. | PI 290508 | V.J.388 | Hungary | 800 |
| 8. | PI 307984 | K930 | Morocco | 800 |
| 9. | PI 326317 | WIR18140 | Azerbaijan | 800 |
| 10. | PI 352480 | Kelcyras | Albania | 800 |
| 11. | PI 355515 | 6975.2 | Asia Minor | 800 |
| 12. | PI 355550 | 69Z5.40 | Switzerland | 800 |
| 13. | PI 427627 | G2158 | Turkey | 800 |
| 14. | PI 427812 | G2550 | Iran | 800 |
| 15. | PI 427927 | G2824 | Iraq | 800 |
| 16. | PI 591871 | H Tri 13605/87 | Georgia | 800 |
| 17. | PI 94740 | 287 | Spain | 800 |
| 18. | PI 94743 | 290 | Russian Federation | 800 |
| 19. | CLTR 13965 | Metzger G68-32 | U.S.A. | 800 |
| Triticum monococcum L. subsp. aegilopoides (Link) Thell. | ||||
| 20. | PI 428011 | G3224 | Azerbaijan | 800 |
| 21. | PI 428012 | G3225 | Armenia | 800 |
| 22. | PI 554513 | 84TK154-02800 | Russian Federation | 800 |
| 23. | PI 614649 | UKR-99-075 | Ukraine | 800 |
| 24. | PI 662221 | GR05-052 | Greece | 800 |
| Secale cereale L. | ||||
| 25. | Piastowskie | Poland | 0 | |
| 26. | Poznańskie | Poland | 0 | |
| 27. | Antonińskie | Poland | 0 | |
| 28. | Hadron | Poland | 0 | |
| 29. | Granat | Poland | 0 | |
| 30. | Turkus | Poland | 0 | |
| Einkorn (Triticum monococcum) PI 191383 | Rye (Secale cereale) Cultivars (Pollen Donors) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Granat | Hadron | Turkus | Antoninskie | Piastowskie | Poznańske | |||||||||||||
| Spike Number | Number of Pollinated Florets | Number of Immature Embryos | CE | Number of Pollinated Florets | Number of Immature Embryos | CE | Number of Pollinated Florets | Number of Immature Embryos | CE | Number of Pollinated Florets | Number of Immature Embryos | CE | Number of Pollinated Florets | Number of Immature Embryos | CE | Number of Pollinated Florets | Number of Immature Embryos | CE |
| 1. | 44 | 3 | 0.07 | 50 | 3 | 0.06 | 42 | 0 | 0.00 | 42 | 1 | 0.02 | 44 | 1 | 0.02 | 38 | 1 | 0.03 |
| 2. | 42 | 1 | 0.02 | 44 | 4 | 0.09 | 38 | 2 | 0.05 | 40 | 0 | 0.00 | 42 | 0 | 0.00 | 36 | 4 | 0.11 |
| 3. | 34 | 2 | 0.06 | 42 | 1 | 0.02 | 44 | 3 | 0.07 | 40 | 1 | 0.03 | 44 | 3 | 0.07 | 52 | 0 | 0.00 |
| 4. | 42 | 4 | 0.10 | 44 | 5 | 0.11 | 38 | 3 | 0.08 | 38 | 3 | 0.08 | 42 | 3 | 0.07 | 50 | 1 | 0.02 |
| 5. | 42 | 0 | 0.00 | 42 | 3 | 0.07 | 44 | 1 | 0.02 | 44 | 3 | 0.07 | 40 | 4 | 0.10 | 48 | 2 | 0.04 |
| 6. | 38 | 4 | 0.11 | 44 | 1 | 0.02 | 38 | 2 | 0.05 | 44 | 1 | 0.02 | 40 | 4 | 0.10 | 52 | 4 | 0.08 |
| 7. | 44 | 3 | 0.07 | 42 | 4 | 0.1 | 34 | 0 | 0.00 | 42 | 2 | 0.05 | 36 | 3 | 0.08 | 54 | 2 | 0.04 |
| 8. | 38 | 3 | 0.08 | 40 | 0 | 0 | 34 | 3 | 0.09 | 40 | 1 | 0.03 | 52 | 0 | 0.00 | 52 | 0 | 0.00 |
| 9. | 44 | 1 | 0.02 | 40 | 3 | 0.08 | 36 | 1 | 0.03 | 40 | 0 | 0.00 | 48 | 2 | 0.04 | 44 | 2 | 0.05 |
| 10. | 38 | 0 | 0.00 | 36 | 3 | 0.08 | 34 | 2 | 0.06 | 36 | 3 | 0.08 | 46 | 2 | 0.04 | 42 | 1 | 0.02 |
| 11. | 34 | 0 | 0.00 | 52 | 3 | 0.06 | 42 | 1 | 0.02 | 52 | 3 | 0.06 | 42 | 1 | 0.02 | 44 | 3 | 0.07 |
| 12. | 34 | 3 | 0.09 | 50 | 0 | 0 | 44 | 2 | 0.05 | 50 | 4 | 0.08 | 44 | 0 | 0.00 | 42 | 3 | 0.07 |
| 13. | 36 | 1 | 0.03 | 48 | 4 | 0.08 | 42 | 4 | 0.10 | 48 | 3 | 0.06 | 42 | 1 | 0.02 | 40 | 1 | 0.03 |
| 14. | 34 | 2 | 0.06 | 52 | 3 | 0.06 | 44 | 1 | 0.02 | 46 | 3 | 0.07 | 44 | 3 | 0.07 | 40 | 1 | 0.03 |
| 15. | 36 | 0 | 0.00 | 54 | 3 | 0.06 | 42 | 4 | 0.10 | 42 | 3 | 0.07 | 42 | 3 | 0.07 | 36 | 4 | 0.11 |
| 16. | 36 | 2 | 0.06 | 52 | 2 | 0.04 | 40 | 0 | 0.00 | 46 | 0 | 0.00 | 40 | 3 | 0.08 | 42 | 3 | 0.07 |
| 17. | 34 | 5 | 0.15 | 50 | 2 | 0.04 | 40 | 3 | 0.08 | 40 | 2 | 0.05 | 40 | 4 | 0.10 | 44 | 1 | 0.02 |
| 18. | 36 | 2 | 0.06 | 52 | 1 | 0.02 | 36 | 3 | 0.08 | 46 | 1 | 0.02 | 36 | 1 | 0.03 | 44 | 2 | 0.05 |
| 19. | 36 | 0 | 0.00 | 42 | 1 | 0.02 | 52 | 3 | 0.06 | 42 | 0 | 0.00 | 42 | 3 | 0.07 | 42 | 4 | 0.10 |
| 20. | 44 | 3 | 0.07 | 40 | 1 | 0.03 | 50 | 0 | 0.00 | 42 | 2 | 0.05 | 44 | 0 | 0.00 | 40 | 3 | 0.08 |
| Summary: | 766 | 39 | 0.05 | 916 | 47 | 0.05 | 814 | 38 | 0.05 | 860 | 36 | 0.04 | 850 | 41 | 0.05 | 882 | 42 | 0.05 |
| No. | Crossing Combination | Number of Mature S1 Plants | Number of S2 Seeds | Number of Mature S2 Plants | Number of S3 Seeds | Number of Mature S3 Plants |
|---|---|---|---|---|---|---|
| 1. | PI 191383 × Granat | 19 | 7 | 0 | 0 | 0 |
| 2. | PI 191383 × Hadron | 26 | 5 | 2 | 79 | 68 |
| 3. | PI 191383 × Turkus | 24 | 8 | 0 | 0 | 0 |
| 4. | PI 191383 × Antoninskie | 13 | 5 | 0 | 0 | 0 |
| 5. | PI 191383 × Piastowskie | 31 | 4 | 0 | 0 | 0 |
| 6. | PI 191383 × Poznańskie | 28 | 9 | 0 | 0 | 0 |
| Summary | 141 | 38 | 2 | 79 | 68 | |
| Generation | Plant No. | Number of Analysed PMCs | Mean Number of Bivalents | Mean Number of Univalents | Mean Number of Multivalents | |||
|---|---|---|---|---|---|---|---|---|
| T. monococcum | S. cereale | T. monococcum | S. cereale | T. monococcum | S. cereale | |||
| S2 | S2_1 | 10 | 6.8 | 4.8 | 0.4 | 0 | 0 | 1.2 |
| S2_2 | 10 | 6.6 | 4.9 | 0.8 | 0 | 0 | 1.3 | |
| summary | 2 | 20 | 6.7 | 4.85 | 0.6 | 0 | 0 | 1.25 |
| S3 | S3_1 | 10 | 6.9 | 6.8 | 0.2 | 0 | 0 | 0.1 |
| S3_2 | 10 | 6.8 | 6.8 | 0.4 | 0 | 0 | 0.1 | |
| S3_3 | 10 | 7 | 7 | 0 | 0 | 0 | 0 | |
| S3_4 | 10 | 7 | 7 | 0 | 0 | 0 | 0 | |
| S3_5 | 10 | 6.9 | 6.8 | 0.2 | 0 | 0 | 0.1 | |
| S3_6 | 10 | 7 | 7 | 0 | 0 | 0 | 0 | |
| S3_7 | 10 | 7 | 7 | 0 | 0 | 0 | 0 | |
| S3_8 | 10 | 6.8 | 6.8 | 0.4 | 0 | 0 | 0.1 | |
| S3_9 | 10 | 7 | 7 | 0 | 0 | 0 | 0 | |
| S3_10 | 10 | 7 | 7 | 0 | 0 | 0 | 0 | |
| summary | 10 | 100 | 6.94 | 6.92 | 0.12 | 0 | 0 | 0.04 |
| Genotypes | Plants | Infection Scores | PCR Products (Base Pairs) NL9F5 (Sr35) | ||
|---|---|---|---|---|---|
| Group 1 | Group 2 | Group 3 | |||
| AmAmRR amphiloids (S3 generation) | 1 | 0 | 0.5 | 0 | 719 * |
| 2 | 0 | 0.5 | 0 | 719 * | |
| 3 | 0 | 0 | 0.5 | 719 * | |
| 4 | 0.5 | 0 | 0.5 | 719 * | |
| 5 | 0.5 | 0.5 | 0.5 | 719 * | |
| 6 | 1 | 0 | 1 | 719 * | |
| 7 | 0.5 | 0 | 0.5 | 719 * | |
| 8 | 0 | 0.5 | 0 | 719 * | |
| 9 | 0.5 | 0.5 | 1 | 719 * | |
| 10 | 0 | 0.5 | 0.5 | 719 * | |
| 11 | 0 | 0.5 | 1 | 719 * | |
| 12 | 0.5 | 0.5 | 0.5 | 719 * | |
| 13 | 0 | 1 | 0.5 | 719 * | |
| 14 | 0.5 | 1 | 0.5 | 719 * | |
| 15 | 1 | 0.5 | 1 | 719 * | |
| 16 | 0.5 | 0.5 | 1 | 719 * | |
| 17 | 0.5 | 0 | 1 | 719 * | |
| 18 | 1 | 1 | 0.5 | 719 * | |
| 19 | 0.5 | 0 | 0 | 719 * | |
| 20 | 0 | 0.5 | 0.5 | 719 * | |
| T. aestivum Thatcher + Sr57 | Mod-1 | 2 | 1 | 2 | 800 |
| Mod-2 | 2 | 2 | 1 | 800 | |
| Mod-3 | 1 | 1 | 2 | 800 | |
| Mod-4 | 2 | 2 | 2 | 800 | |
| Mod-5 | 2 | 1 | 1 | 800 | |
| Mod-6 | 1 | 1 | 1 | 800 | |
| Mod-7 | 1 | 1 | 2 | 800 | |
| Mod-8 | 1 | 2 | 1 | 800 | |
| Mod-9 | 2 | 2 | 1 | 800 | |
| Mod-10 | 1 | 1 | 1 | 800 | |
| T. aestivum “Thatcher” | Sus-1 | 4 | 4 | 4 | 800 |
| Sus-2 | 4 | 3 | 4 | 800 | |
| Sus-3 | 4 | 4 | 4 | 800 | |
| Sus-4 | 4 | 4 | 4 | 800 | |
| Sus-5 | 4 | 4 | 4 | 800 | |
| Sus-6 | 3 | 4 | 3 | 800 | |
| Sus-7 | 4 | 4 | 4 | 800 | |
| Sus-8 | 4 | 4 | 4 | 800 | |
| Sus-9 | 3 | 4 | 4 | 800 | |
| Sus-10 | 4 | 4 | 4 | 800 | |
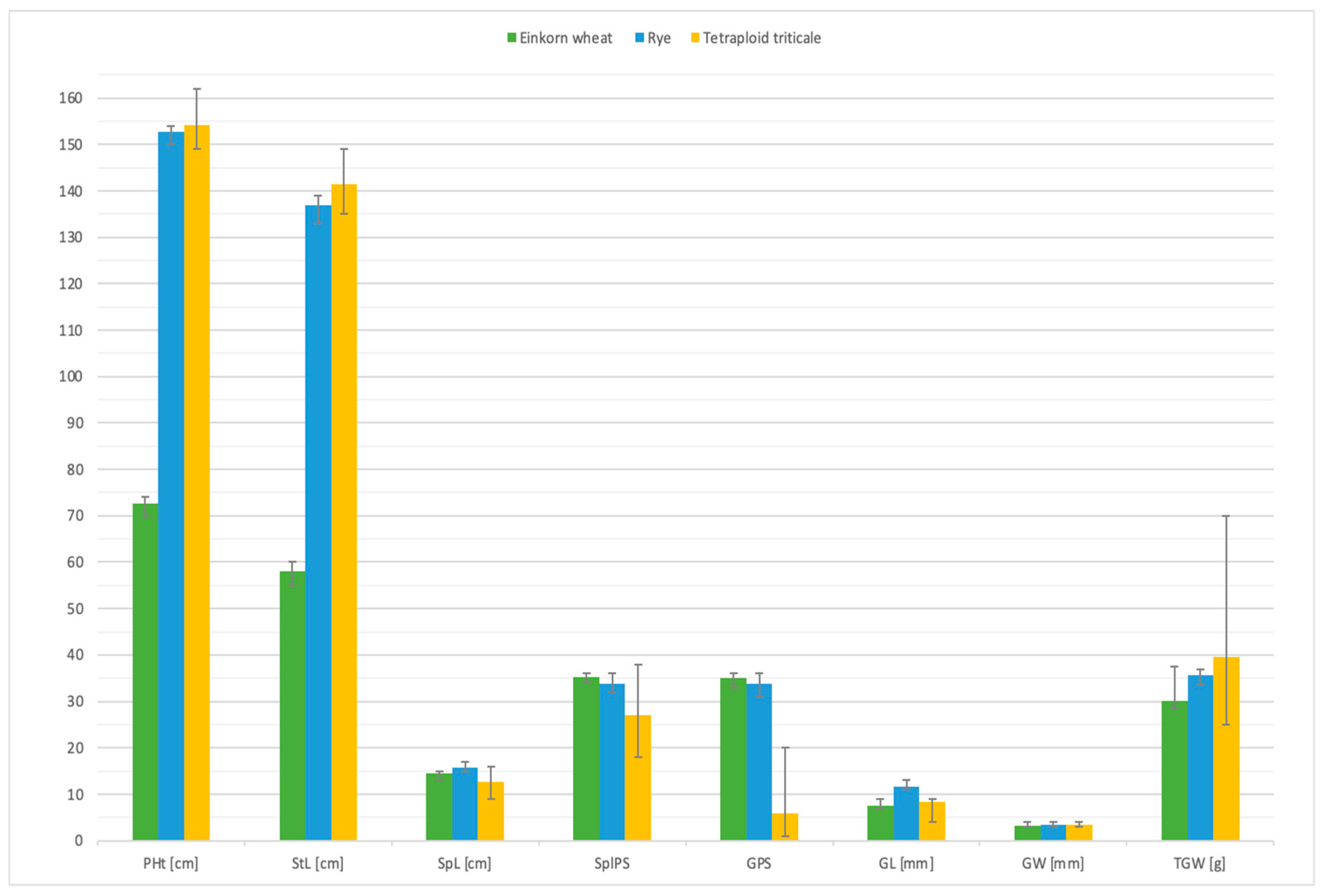
| Genotypes | Summary Values | PHt [cm] | StL [cm] | SpL [cm] | SplPS | GPS | GL [mm] | GW [mm] | TGW [g] |
|---|---|---|---|---|---|---|---|---|---|
| Einkorn wheat Triticum monococcum AmAPI 191383 | Mean | 72.5333 | 58.1 | 14.4333 | 35.2667 | 35.1 | 7.6167 | 3.2333 | 30.2 |
| Range | 70–74 | 55–60 | 13–15 | 34–36 | 33–36 | 7–9 | 3–4 | 28.4–37.6 | |
| Variance | 1.2023 | 2.0237 | 0.4192 | 0.9446 | 1.0407 | 0.4438 | 0.1819 | 5.7464 | |
| Standard deviation | 1.0965 | 1.4226 | 0.6475 | 0.9719 | 1.0201 | 0.6662 | 0.4265 | 2.3972 | |
| Standard error | 0.1416 | 0.1837 | 0.0836 | 0.1255 | 0.1317 | 0.086 | 0.0551 | 0.3095 | |
| Rye Secale cereale RR Dankowskie Hadron | Mean | 152.7833 | 136.95 | 15.8333 | 33.8333 | 33.7167 | 11.5333 | 3.3667 | 35.7533 |
| Range | 150–154 | 133–139 | 15–17 | 32–36 | 31–36 | 11–13 | 3–4 | 33.6–36.9 | |
| Variance | 0.9862 | 2.2517 | 0.75514 | 1.9379 | 2.0031 | 0.4565 | 0.2362 | 0.732 | |
| Standard deviation | 0.9931 | 1.5006 | 0.8668 | 1.9321 | 1.4153 | 0.6756 | 0.486 | 0.8556 | |
| Standard error | 0.4282 | 0.1937 | 0.1119 | 0.1797 | 0.1827 | 0.0872 | 0.0627 | 0.1105 | |
| Amphiploids AmAmRR S4 plants | Mean | 154.1833 | 141.45 | 12.75 | 26.9667 | 5.8833 | 8.2833 | 3.3167 | 39.4967 |
| Range | 149–162 | 135–149 | 9–16 | 18–38 | 1–20 | 4–9 | 3–4 | 25.1–70.2 | |
| Variance | 12.254 | 15.8449 | 3.2754 | 19.1175 | 20.4438 | 1.2573 | 0.2201 | 54.219 | |
| Standard deviation | 3.5006 | 3.9806 | 1.8098 | 4.3724 | 4.5215 | 1.1213 | 0.4691 | 7.3634 | |
| Standard error | 0.4519 | 0.5139 | 0.2336 | 0.5645 | 0.5837 | 0.1448 | 0.0606 | 0.9506 | |
| Analysis of variance (ANOVA)—treatment between groups | |||||||||
| F–value | 27,229.76 | 19,658.9 | 96.49 | 161.04 | 2082.24 | 366.34 | 1.28 | 64.89 | |
| p–value | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.285598 | <0.0001 | |
| Tukey’s Highest Significant Difference (HSD) between any two sample means at designated level | |||||||||
| AmAm vs. RR | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.05 | p < 0.05 | p < 0.01 | n/a | p < 0.01 | |
| AmAm vs. AmAmRR | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 | n/a | p < 0.01 | |
| RR vs. AmAmRR | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 | n/a | p < 0.01 | |
| Significance level | HSD [0.05] | 0.95 | 1.12 | 0.53 | 1.17 | 1.21 | 0.37 | n/a | 1.94 |
| HSD [0.01] | 1.18 | 1.39 | 0.66 | 1.46 | 1.51 | 0.46 | n/a | 2.42 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiatek, M.T.; Noweiska, A.; Bobrowska, R.; Czapiewska, A.; Aygün, M.; Munyamahoro, F.d.; Mikołajczyk, S.; Tomkowiak, A.; Kurasiak-Popowska, D.; Poślednik, P. Novel Tetraploid Triticale (Einkorn Wheat × Rye)—A Source of Stem Rust Resistance. Plants 2023, 12, 278. https://doi.org/10.3390/plants12020278
Kwiatek MT, Noweiska A, Bobrowska R, Czapiewska A, Aygün M, Munyamahoro Fd, Mikołajczyk S, Tomkowiak A, Kurasiak-Popowska D, Poślednik P. Novel Tetraploid Triticale (Einkorn Wheat × Rye)—A Source of Stem Rust Resistance. Plants. 2023; 12(2):278. https://doi.org/10.3390/plants12020278
Chicago/Turabian StyleKwiatek, Michał T., Aleksandra Noweiska, Roksana Bobrowska, Adrianna Czapiewska, Mert Aygün, Francois d’Assise Munyamahoro, Sylwia Mikołajczyk, Agnieszka Tomkowiak, Danuta Kurasiak-Popowska, and Paweł Poślednik. 2023. "Novel Tetraploid Triticale (Einkorn Wheat × Rye)—A Source of Stem Rust Resistance" Plants 12, no. 2: 278. https://doi.org/10.3390/plants12020278