
Functional Characterization of TkSRPP Promoter in Response to Hormones and Wounding Stress in Transgenic Tobacco
 ,
,
Abstract
:1. Introduction
2. Results
2.1. The 5′ Flanking Sequence of TkSRPP Promoter Cloning and Sequence Analysis
2.2. cis-Element Analysis of TkSRPP Promoter
2.3. Activity Analysis of Various SRPP Promoters
2.4. Genetic Transformation of Different TkSRPP Promoters
2.5. Expression Analysis of SRPP Promoter in Tobacco Different Tissues
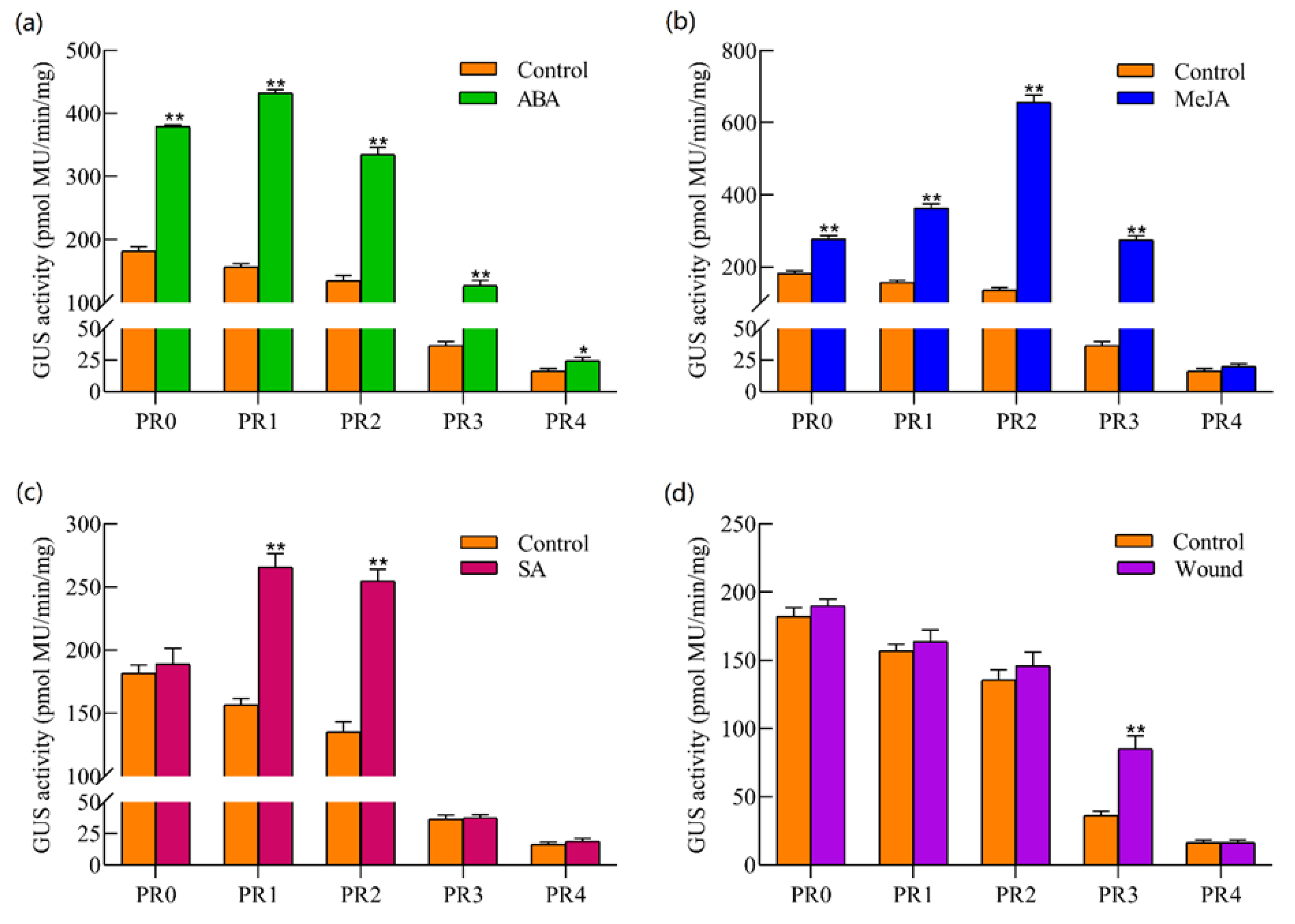
2.6. GUS Activity Analysis of TkSRPP Promoters under Hormones and Wounding Stress
2.7. Expression Analysis of TkSRPP Gene under Hormones and Wounding Stress
3. Discussion
3.1. The Key Region Function of SRPP Promoter
3.2. SRPP Is a Typical Stress-Induced Gene in Plants
3.3. The TkSRPP Gene Is Expected to Promote Rubber Biosynthesis under Hormones and Wounding Stress
4. Materials and Methods
4.1. Plant Materials, Growth Conditions
4.2. Cloning of the HbSRPP Genomic Sequence
4.3. Analysis of cis-Regulatory Elements in the TkSRPP Promoter
4.4. Construction of the TkSRPP Promoter::GUS Plasmids
4.5. Histochemical Straining Assays
4.6. Transgenic Plant Generation
4.7. Tissue Specificity of TkSRPP Promoter
4.8. Abiotic Stress Treatments
4.9. GUS Fluorimetric Assays
4.10. qRT-PCR
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, L.; Nie, Z.; Kang, G.; Duan, C.; Li, Y.; Zeng, R. In-depth proteome analysis of the rubber particle of Hevea brasiliensis (para rubber tree). Plant Mol. Biol. 2013, 82, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Feeney, M.; Ahmadi, M.; Lonoce, C.; Sajari, R.; Di Cola, A.; Frigerio, L. Subcellular localization and interactions among rubber particle proteins from Hevea brasiliensis. J. Exp. Bot. 2017, 68, 5045–5055. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, J.; Wu, Y.; You, Y.; He, J.; Zhang, J.; Zhang, L.; Dong, Y. Micromorphological characterization and label-free quantitation of small rubber particle protein in natural rubber latex. Anal. Biochem. 2016, 499, 34–42. [Google Scholar] [CrossRef]
- Collins-Silva, J.; Nural, A.T.; Skaggs, A.; Scott, D.; Hathwaik, U.; Woolsey, R.; Schegg, K.; McMahan, C.; Whalen, M.; Cornish, K.; et al. Altered levels of the Taraxacum kok-saghyz (Russian dandelion) small rubber particle protein, TkSRPP3, result in qualitative and quantitative changes in rubber metabolism. Phytochemistry 2012, 79, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Cherian, S.; Ryu, S.B.; Cornish, K. Natural rubber biosynthesis in plants, the rubber transferase complex, and metabolic engineering progress and prospects. Plant Biotechnol. J. 2019, 17, 2041–2061. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.J.; Ryu, S.B.; Kwak, Y.S.; Kang, H. A novel cDNA from Parthenium argentatum Gray enhances the rubber biosynthetic activity in vitro. J. Exp. Bot. 2004, 55, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Hillebrand, A.; Post, J.J.; Wurbs, D.; Wahler, D.; Lenders, M.; Krzyzanek, V.; Pruefer, D.; Gronover, C.S. Down-Regulation of Small Rubber Particle Protein Expression Affects Integrity of Rubber Particles and Rubber Content in Taraxacum brevicorniculatum. PLoS ONE 2012, 7, e41874. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.Y.; Seo, Y.S.; Lee, H.; Kim, W.T. Constitutive expression of CaSRP1, a hot pepper small rubber particle protein homolog, resulted in fast growth and improved drought tolerance in transgenic Arabidopsis plants. Planta 2010, 232, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Park, K.Y.; Seo, Y.S.; Kim, W.T. Arabidopsis Small Rubber Particle Protein Homolog SRPs Play Dual Roles as Positive Factors for Tissue Growth and Development and in Drought Stress Responses. Plant Physiol. 2016, 170, 2494–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amerik, A.Y.; Martirosyan, Y.T.; Gachok, I.V. Regulation of Natural Rubber Biosynthesis by Proteins Associated with Rubber Particles. Russ. J. Bioorganic Chem. 2018, 44, 140–149. [Google Scholar] [CrossRef]
- Men, X.; Wang, F.; Chen, G.Q.; Zhang, H.B.; Xian, M. Biosynthesis of Natural Rubber: Current State and Perspectives. Int. J. Mol. Sci. 2018, 20, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tata, S.K.; Choi, J.Y.; Jung, J.-Y.; Lim, K.Y.; Shin, J.S.; Ryu, S.B. Laticifer tissue-specific activation of the Hevea SRPP promoter in Taraxacum brevicorniculatum and its regulation by light, tapping and cold stress. Ind. Crops Prod. 2012, 40, 219–224. [Google Scholar] [CrossRef]
- Guo, D.; Li, H.-L.; Peng, S.-Q. Molecular and functional characterization of the HbSRPP promoter in response to hormones and abiotic stresses. Transgenic Res. 2013, 23, 331. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.M.; Guo, D.; Yang, S.; Shi, M.; Chao, J.; Li, H.; Peng, S.; Tian, W.M. Jasmonate signalling in regulation of rubber biosynthesis in laticifer cells of rubber tree (Hevea brasiliensis Muell. Arg.). J. Exp. Bot. 2018, 69, 3559–3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Zhai, J.; Wang, Q.; Yuan, H.; Huang, X. Function of Hevea brasiliensis NAC1 in dehydration-induced laticifer differentiation and latex biosynthesis. Planta 2017, 245, 31–44. [Google Scholar] [CrossRef]
- Li, H.L.; Wei, L.R.; Guo, D.; Wang, Y.; Zhu, J.H.; Chen, X.T.; Peng, S.Q. HbMADS4, a MADS-box Transcription Factor from Hevea brasiliensis, Negatively Regulates HbSRPP. Front. Plant Sci. 2016, 7, 1709. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.; Xu, X.; Ruan, J.; Liu, S.; Wu, S.; Shao, X.; Wang, X.; Gan, L.; Qin, B.; Yang, Y.; et al. Genome analysis of Taraxacum kok-saghyz Rodin provides new insights into rubber biosynthesis. Natl. Sci. Rev. 2018, 5, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.W.; Yan, J.; Lei, J.; Li, J.; Zhu, J.; Zhang, H. De novo Transcriptome Sequencing of MeJA-Induced Taraxacum koksaghyz Rodin to Identify Genes Related to Rubber Formation. Sci. Rep. 2017, 7, 15697. [Google Scholar] [CrossRef] [Green Version]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Li, A.; Chen, L.; Ren, H.; Wang, X.; Zhang, H.; Huang, R.F. Analysis of the essential DNA region for OsEBP-89 promoter in response to methyl jasmonic acid. Sci. China Ser. C Life Sci. 2008, 51, 280–285. [Google Scholar] [CrossRef]
- Wang, C.; Gao, G.; Cao, S.; Xie, Q.; Qi, H. Isolation and functional validation of the CmLOX08 promoter associated with signalling molecule and abiotic stress responses in oriental melon, Cucumis melo var. makuwa Makino. BMC Plant Biol. 2019, 19, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, G.-J.; Xue, X.-Y.; Mao, Y.-B.; Wang, L.-J.; Chen, X.-Y. Arabidopsis MYC2 Interacts with DELLA Proteins in Regulating Sesquiterpene Synthase Gene Expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [Green Version]
- Freitas, E.O.; Melo, B.P.; Lourenco-Tessutti, I.T.; Arraes, F.B.M.; Amorim, R.M.; Lisei-de-Sa, M.E.; Costa, J.A.; Leite, A.G.B.; Faheem, M.; Ferreira, M.A.; et al. Identification and characterization of the GmRD26 soybean promoter in response to abiotic stresses: Potential tool for biotechnological application. BMC Biotechnol. 2019, 19, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, G.J.; Yan, X.F.; Wei, Z.G.; Xu, Z.R. MeJA-inducible expression of the heterologous JAZ2 promoter from Arabidopsis in Populus trichocarpa protoplasts. J. Plant Dis. Prot. 2011, 118, 69–74. [Google Scholar] [CrossRef]
- Wang, Q.; Yuan, F.; Pan, Q.; Li, M.; Wang, G.; Zhao, J.; Tang, K. Isolation and functional analysis of the Catharanthus roseus deacetylvindoline-4-O-acetyltransferase gene promoter. Plant Cell Rep. 2010, 29, 185–192. [Google Scholar] [CrossRef]
- Goldsbrough, A.P.; Albrecht, H.; Stratford, R. Salicylic acid-inducible binding of a tobacco nuclear protein to a 10 bp sequence which is highly conserved amongst stress-inducible genes. Plant J. 1993, 3, 563–571. [Google Scholar] [CrossRef]
- Wen, Z.; Yao, L.; Singer, S.D.; Muhammad, H.; Li, Z.; Wang, X. Constitutive heterologous overexpression of a TIR-NB-ARC-LRR gene encoding a putative disease resistance protein from wild Chinese Vitis pseudoreticulata in Arabidopsis and tobacco enhances resistance to phytopathogenic fungi and bacteria. Plant Physiol. Biochem. 2017, 112, 346–361. [Google Scholar] [CrossRef]
- Liu, H.; Lyu, H.M.; Zhu, K.; Van de Peer, Y.; Cheng, Z.M. The emergence and evolution of intron-poor and intronless genes in intron-rich plant gene families. Plant J. 2020, 105, 1072–1082. [Google Scholar] [CrossRef]
- Laibach, N.; Schmidl, S.; Muller, B.; Bergmann, M.; Prufer, D.; Schulze Gronover, C. Small rubber particle proteins from Taraxacum brevicorniculatum promote stress tolerance and influence the size and distribution of lipid droplets and artificial poly(cis-1,4-isoprene) bodies. Plant J. 2018, 93, 1045–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Z.; Wang, D.; Sun, Y.; Yang, Q.; Meng, X.; Wang, L.; Feng, W.; Li, L.; Wurtele, E.S.; Wang, X. Comparative Proteomics of Rubber Latex Revealed Multiple Protein Species of REF/SRPP Family Respond Diversely to Ethylene Stimulation among Different Rubber Tree Clones. Int. J. Mol. Sci. 2017, 18, 958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasternack, C.; Strnad, M. Jasmonates are signals in the biosynthesis of secondary metabolites—Pathways, transcription factors and applied aspects—A brief review. New Biotechnol. 2017, 48, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Laosombut, T.; Arreewichit, P.; Nirapathpongporn, K.; Traiperm, P.; Kongsawadworakul, P.; Viboonjun, U.; Narangajavana, J. Differential Expression of Methyl Jasmonate-Responsive Genes Correlates with Laticifer Vessel Proliferation in Phloem Tissue of Rubber Tree (Hevea brasiliensis). J. Plant Growth Regul. 2016, 35, 1049–1063. [Google Scholar] [CrossRef]
- Liu, J.P.; Hu, J.; Liu, Y.H.; Yang, C.P.; Zhuang, Y.F.; Guo, X.L.; Li, Y.J.; Zhang, L. Transcriptome analysis of Hevea brasiliensis in response to exogenous methyl jasmonate provides novel insights into regulation of jasmonate-elicited rubber biosynthesis. Physiol. Mol. Biol. Plants 2018, 24, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Koo, A.J.; Gao, X.; Jones, A.D.; Howe, G.A. A rapid wound signal activates the systemic synthesis of bioactive jasmonates in Arabidopsis. Plant J. 2009, 59, 974–986. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, F.; Chen, L.; Pan, Y.; Sun, L.; Bao, N.; Zhang, T.; Cui, C.X.; Qiu, Z.; Zhang, Y.; et al. Jasmonate-mediated wound signalling promotes plant regeneration. Nat. Plants 2019, 5, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Rymen, B.; Sugimoto, K. How do plants transduce wound signals to induce tissue repair and organ regeneration? Curr. Opin. Plant Biol. 2020, 57, 72–77. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes Under Abiotic Stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Hu, Y.; Wang, H.; Guo, Q.; Chen, Y.; Howe, G.A.; Yu, D. Molecular Mechanism Underlying the Synergetic Effect of Jasmonate on Abscisic Acid Signaling during Seed Germination in Arabidopsis. Plant Cell 2020, 32, 3846–3865. [Google Scholar] [CrossRef]
- Allu, A.D.; Brotman, Y.; Xue, G.P.; Balazadeh, S. Transcription factor ANAC032 modulates JA/SA signalling in response to Pseudomonas syringae infection. EMBO Rep. 2016, 17, 1578–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallois, P.; Marinho, P. Leaf disc transformation using agrobacterium tumefaciens—Expression of heterologous genes in tobacco. Methods Mol. Biol. 1995, 49, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Atsumi, G. CRISPR/Cas9-mediated knockout of the RDR6 gene in Nicotiana benthamiana for efficient transient expression of recombinant proteins. Planta 2019, 250, 463–473. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cis-Elements | Number | Function | Motif |
|---|---|---|---|
| CAAT-box | 38 | common cis-acting element in eukaryotic genes | CAAT |
| TATA-box | 28 | core promoter element around −30 of transcription start | TATA |
| ARE | 5 | cis-acting regulatory element essential for the anaerobic induction | AAACCA |
| MBS | 3 | MYB binding site involved in drought-inducibility | CAACTG |
| O2-site | 3 | cis-acting regulatory element involved in zein metabolism regulation | GTAC |
| GT1-motif | 2 | light responsive element | GGTTAA |
| TCA-element | 4 | cis-acting element involved in salicylic acid responsiveness | CCATCTTTTT |
| I-box | 1 | part of a light responsive element | GATAA |
| chs-CMA1a | 1 | part of a light responsive element | TCACTTGA |
| Box 4 | 1 | part of a conserved DNA module involved in light responsiveness | ATTAAT |
| CAT-box | 1 | cis-acting regulatory element related to meristem expression | GCCACT |
| CGTCA-motif | 1 | cis-acting regulatory element involved in the MeJA-responsiveness | CGTCA |
| G-box | 2 | cis-acting regulatory element involved in light responsiveness | CACATGG |
| MRE | 1 | MYB binding site involved in light responsiveness | AACCTAA |
| TGACG-motif | 1 | cis-acting regulatory element involved in the MeJA-responsiveness | TGACG |
| WUN-motif | 1 | wound-responsive element | TCATTACGAA |
| Gap-box | 1 | part of a light responsive element | CAAATGAA(A/G)A |
| GA-motif | 1 | part of a light responsive element | ATAGATAA |
| GATA-motif | 1 | part of a light responsive element | AAGATAAGATT |
| P-box | 1 | gibberellin-responsive element | CCTTTTG |
| TCT-motif | 1 | part of a light responsive element | TCTTAC |
| ABRE | 1 | Abscisic acid response cis-acting element | ACTG |
| TGA-element | 1 | auxin-responsive element | AACGAC |
| 3-AF1 4-binding site | 1 | light responsive element | TAAGAGAGGAA |
| W-box | 1 | WRKY binding wite involved in salicylic acid (SA)-induced responsiveness | TTGAC |
| TCA | 1 | cis-acting element involved in salicylic acid responsiveness | TCATCTTCAT |
| GATABOX | 48 | Required for light regulation and tissue-specific expression | GATA |
| EBOXBNNAPA | 24 | Abscisic acid response cis-acting element | CANNTG |
| MYBCORE | 9 | ABA response element | CNGTTR |
| CCAATBOX1 | 5 | Work with HSE to increase the activity of the promoter | CCAAT |
| WBOXATNPR1 | 4 | Important response component of SA | TTGAC |
| MYB | 5 | Unknown | CAACCA |
| MYC | 4 | Unknown | CATTTG |
| MYB-like | 1 | Unknown | TAACCA |
| Myb | 4 | Unknown | CAACTG |
| Myb-binding site | 1 | Unknown | CAACAG |
| STRE | 4 | Unknown | AGGGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, G.; Fan, M.; Wang, H.; Leng, Y.; Sun, J.; Huang, J.; Zhang, H.; Yan, J. Functional Characterization of TkSRPP Promoter in Response to Hormones and Wounding Stress in Transgenic Tobacco. Plants 2023, 12, 252. https://doi.org/10.3390/plants12020252
Dong G, Fan M, Wang H, Leng Y, Sun J, Huang J, Zhang H, Yan J. Functional Characterization of TkSRPP Promoter in Response to Hormones and Wounding Stress in Transgenic Tobacco. Plants. 2023; 12(2):252. https://doi.org/10.3390/plants12020252
Chicago/Turabian StyleDong, Gaoquan, Mengwei Fan, Hainan Wang, Yadong Leng, Junting Sun, Jun Huang, Hao Zhang, and Jie Yan. 2023. "Functional Characterization of TkSRPP Promoter in Response to Hormones and Wounding Stress in Transgenic Tobacco" Plants 12, no. 2: 252. https://doi.org/10.3390/plants12020252