
Role of Arbuscular Mycorrhizal Fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses

 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
1.1. Economic Importance of Soil Microbes
1.2. Molecular Mechanisms of Plant-Microbe Interactions
2. Arbuscular Mycorrhizal Fungi Background
2.1. Sustainability of Agriculture and AM Fungi
2.2. AM Fungi in Crops Defense Mechanisms
2.3. AM Fungi Play an Essential Role in Increasing Crops’ Productivity Maintenance of Soil Health
2.4. Role of AMF in the Ecosystem and Mitigation of Environmental Stresses
2.4.1. Wasteland Salvage
2.4.2. Mitigation of Drought Stress
2.4.3. Bioremediation
2.4.4. AMF-Induced Changes in the Host to Deal with HMs
2.4.5. Temperature (High and Low)
2.4.6. Salt Strain
2.4.7. AMF and Combined Abiotic Stresses
3. Importance of AMF in Soil Fertility
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kastner, T.; Chaudhary, A.; Gingrich, S.; Marques, A.; Persson, U.M.; Bidoglio, G.; Le Provost, G.; Schwarzmüller, F. Global agricultural trade and land system sustainability: Implications for ecosystem carbon storage, biodiversity, and human nutrition. One Earth 2021, 4, 1425–1443. [Google Scholar] [CrossRef]
- Byerlee, D.; Falcon, W.P.; Naylor, R. The Tropical Oil Crop Revolution: Food, Feed, Fuel, and Forests; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Finucane, M.L.; Acosta, J.; Wicker, A.; Whipkey, K. Short-term solutions to a long-term challenge: Rethinking disaster recovery planning to reduce vulnerabilities and inequities. Int. J. Environ. Res. Public Health 2020, 17, 482. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Khanra, S.; Haldar, G.; Bhowmick, T.K.; Gayen, K. Diverse cyanobacteria resource from north east region of India for valuable biomolecules: Phycobiliprotein, carotenoid, carbohydrate and lipid. Curr. Biochem. Eng. 2019, 5, 21–33. [Google Scholar] [CrossRef]
- Sahoo, G.; Wani, A.M.; Swamy, S.L.; Roul, P.K.; Dash, A.C.; Sharma, A. Livelihood strategy and sustainability aspects in industrialization as a source of employment in rural areas. In Social Morphology, Human Welfare, and Sustainability; Springer: Berlin/Heidelberg, Germany, 2022; pp. 643–670. [Google Scholar]
- Brevik, E.C.; Slaughter, L.; Singh, B.R.; Steffan, J.J.; Collier, D.; Barnhart, P.; Pereira, P. Soil and human health: Current status and future needs. Air Soil Water Res. 2020, 13, 1178622120934441. [Google Scholar] [CrossRef]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef]
- Taghvaei, M.; Kordestani, M.D.; Saleh, M.; Mastinu, A. The Reinforcement of Early Growth, Extract, and Oil of Silybum marianum L. by Polymer Organic Cover and Bacteria Inoculation under Water Deficit. Soil Syst. 2023, 7, 61. [Google Scholar] [CrossRef]
- Motamedi, M.; Zahedi, M.; Karimmojeni, H.; Motamedi, H.; Mastinu, A. Effect of rhizosphere bacteria on antioxidant enzymes and some biochemical characteristics of Medicago sativa L. subjected to herbicide stress. Acta Physiol. Plant. 2022, 44, 84. [Google Scholar] [CrossRef]
- Chanclud, E.; Morel, J.B. Plant hormones: A fungal point of view. Mol Plant Pathol 2016, 17, 1289–1297. [Google Scholar] [CrossRef]
- Luty, L.; Musiał, K.; Zioło, M. The role of selected ecosystem services in different farming systems in Poland regarding the differentiation of agricultural land structure. Sustainability 2021, 13, 6673. [Google Scholar] [CrossRef]
- Fernández, J.A.; Ayastuy, M.E.; Belladonna, D.P.; Comezaña, M.M.; Contreras, J.; de Maria Mourão, I.; Orden, L.; Rodríguez, R.A. Current Trends in Organic Vegetable Crop Production: Practices and Techniques. Horticulturae 2022, 8, 893. [Google Scholar] [CrossRef]
- Ngosong, C.; Tatah, B.N.; Olougou, M.N.E.; Suh, C.; Nkongho, R.N.; Ngone, M.A.; Achiri, D.T.; Tchakounté, G.V.T.; Ruppel, S. Inoculating plant growth-promoting bacteria and arbuscular mycorrhiza fungi modulates rhizosphere acid phosphatase and nodulation activities and enhance the productivity of soybean (Glycine max). Front. Plant Sci. 2022, 13, 934339. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, L.; Zhou, J.; Rengel, Z.; George, T.S.; Feng, G. Exploring the secrets of hyphosphere of arbuscular mycorrhizal fungi: Processes and ecological functions. Plant Soil 2022, 418, 1–22. [Google Scholar] [CrossRef]
- Wild, B.; Gerrits, R.; Bonneville, S. The contribution of living organisms to rock weathering in the critical zone. npj Mater. Degrad. 2022, 6, 98. [Google Scholar] [CrossRef]
- Bhattacharjee, A.; Qafoku, O.; Richardson, J.A.; Anderson, L.N.; Schwarz, K.; Bramer, L.M.; Lomas, G.X.; Orton, D.J.; Zhu, Z.; Engelhard, M.H. A Mineral-Doped Micromodel Platform Demonstrates Fungal Bridging of Carbon Hot Spots and Hyphal Transport of Mineral-Derived Nutrients. Msystems 2022, 7, e0091322. [Google Scholar] [CrossRef]
- Tiwari, J.; Ma, Y.; Bauddh, K. Arbuscular mycorrhizal fungi: An ecological accelerator of phytoremediation of metal contaminated soils. Arch. Agron. Soil Sci. 2022, 68, 283–296. [Google Scholar] [CrossRef]
- Dong, H.; Huang, L.; Zhao, L.; Zeng, Q.; Liu, X.; Sheng, Y.; Shi, L.; Wu, G.; Jiang, H.; Li, F. A critical review of mineral–microbe interaction and co-evolution: Mechanisms and applications. Natl. Sci. Rev. 2022, 9, nwac128. [Google Scholar] [CrossRef]
- O’Callaghan, M.; Ballard, R.A.; Wright, D. Soil microbial inoculants for sustainable agriculture: Limitations and opportunities. Soil Use Manag. 2022, 38, 1340–1369. [Google Scholar] [CrossRef]
- Omomowo, O.I.; Babalola, O.O. Bacterial and fungal endophytes: Tiny giants with immense beneficial potential for plant growth and sustainable agricultural productivity. Microorganisms 2019, 7, 481. [Google Scholar] [CrossRef]
- Sangwan, S.; Prasanna, R. Mycorrhizae helper bacteria: Unlocking their potential as bioenhancers of plant–arbuscular mycorrhizal fungal associations. Microb. Ecol. 2022, 84, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Lai, S.; Chen, M.; Long, X.; Fu, X.; Yang, H. Effects of grass cultivation on soil arbuscular mycorrhizal fungi community in a tangerine orchard. Rhizosphere 2022, 24, 100583. [Google Scholar] [CrossRef]
- Tsiknia, M.; Tsikou, D.; Papadopoulou, K.K.; Ehaliotis, C. Multi-species relationships in legume roots: From pairwise legume-symbiont interactions to the plant–microbiome–soil continuum. FEMS Microbiol. Ecol. 2021, 97, fiaa222. [Google Scholar] [CrossRef]
- Garg, N.; Cheema, A. Relative roles of Arbuscular Mycorrhizae in establishing a correlation between soil properties, carbohydrate utilization and yield in Cicer arietinum L. under as stress. Ecotoxicol. Environ. Saf. 2021, 207, 111196. [Google Scholar] [CrossRef]
- Andrade, S.; Silveira, A.; Mazzafera, P. Arbuscular mycorrhiza alters metal uptake and the physiological response of Coffea arabica seedlings to increasing Zn and Cu concentrations in soil. Sci. Total Environ. 2010, 408, 5381–5391. [Google Scholar] [CrossRef]
- Oruru, M.B.; Njeru, E.M. Upscaling arbuscular mycorrhizal symbiosis and related agroecosystems services in smallholder farming systems. BioMed Res. Int. 2016, 2016, 4376240. [Google Scholar] [CrossRef]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Diagne, N.; Ndour, M.; Djighaly, P.I.; Ngom, D.; Ngom, M.C.N.; Ndong, G.; Svistoonoff, S.; Cherif-Silini, H. Effect of plant growth promoting rhizobacteria (PGPR) and arbuscular mycorrhizal fungi (AMF) on salt stress tolerance of Casuarina obesa (Miq.). Front. Sustain. Food Syst. 2020, 4, 601004. [Google Scholar] [CrossRef]
- Emmanuel, O.C.; Babalola, O.O. Productivity and quality of horticultural crops through co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiol. Res. 2020, 239, 126569. [Google Scholar] [CrossRef]
- Wahid, F.; Fahad, S.; Danish, S.; Adnan, M.; Yue, Z.; Saud, S.; Siddiqui, M.H.; Brtnicky, M.; Hammerschmiedt, T.; Datta, R. Sustainable management with mycorrhizae and phosphate solubilizing bacteria for enhanced phosphorus uptake in calcareous soils. Agriculture 2020, 10, 334. [Google Scholar] [CrossRef]
- Wang, F.; Adams, C.A.; Yang, W.; Sun, Y.; Shi, Z. Benefits of arbuscular mycorrhizal fungi in reducing organic contaminant residues in crops: Implications for cleaner agricultural production. Crit. Rev. Environ. Sci. Technol. 2020, 50, 1580–1612. [Google Scholar] [CrossRef]
- Mishra, V.; Ellouze, W.; Howard, R. Utility of arbuscular mycorrhizal fungi for improved production and disease mitigation in organic and hydroponic greenhouse crops. J. Hortic 2018, 5, 237. [Google Scholar] [CrossRef]
- Buczkowska, H.B.; Sałata, A. Inoculation with arbuscular mycorrhizal fungi (AMF) and plant irrigation with yield–forming factors in organic sweet pepper (Capsicum annuum L.) cultivation. Acta Sci. Pol. Hortorum Cultus 2020, 19, 125–138. [Google Scholar] [CrossRef]
- Volpato, S.; Masoero, G.; Giovannetti, G.; Nuti, M. Arbuscular mycorrhizal biofertilizers sources in the potato (Solanum tuberosum) plant show interactions with cultivars on yield and litter-bags Spectral Features. J. Agron. Res. 2020, 2, 10–17. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O.O.; Cheseto, X.; Torto, B. Effects of rhizobia and arbuscular mycorrhizal fungi on yield, size distribution and fatty acid of soybean seeds grown under drought stress. Microbiol. Res. 2021, 242, 126640. [Google Scholar] [CrossRef]
- Jamal, A.; Ayub, N.; Usman, M.; Khan, A.G. Arbuscular mycorrhizal fungi enhance zinc and nickel uptake from contaminated soil by soybean and lentil. Int. J. Phytoremediation 2002, 4, 205–221. [Google Scholar] [CrossRef]
- McKean, T. Effects of Soilless Substrate Systems and Environmental Conditions on Yield, Total Soluble Solids, and Titratable Acidity of Greenhouse Strawberry (Fragaria × ananassa). Master’s Thesis, The Ohio State University, Columbus, OH, USA, 2019. [Google Scholar]
- Quan, L.; Duan, K.; Wei, Z.; Li, W.; Chen, Y.; Duan, W.; Qin, C.; Shen, Z.; Xia, Y. Beneficial effects of arbuscular mycorrhizae on Cu detoxification in Mimosa pudica L. grown in Cu-polluted soils. Environ. Sci. Pollut. Res. 2022, 30, 25755–25763. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Quiroz, A.; Herrera, H.; Soto, J.; Campos-Vargas, R.; Ortiz, J.; Arriagada, C. Rhizosphere fungi regulate the expression of metal tolerance genes in Solanum lycopersicum L. (Solanaceae) growing in a metal (loid)-contaminated soil. Rhizosphere 2022, 24, 100599. [Google Scholar] [CrossRef]
- Boyno, G.; Demir, S.; Danesh, Y.R. Effects of some biological agents on the growth and biochemical parameters of tomato plants infected with Alternaria solani (Ellis & Martin) Sorauer. Eur. J. Plant Pathol. 2022, 162, 19–29. [Google Scholar]
- Fracasso, A.; Telò, L.; Lanfranco, L.; Bonfante, P.; Amaducci, S. Physiological beneficial effect of Rhizophagus intraradices inoculation on tomato plant yield under water deficit conditions. Agronomy 2020, 10, 71. [Google Scholar] [CrossRef]
- Sarker, M.; Ali, M.; Islam, M.; Huda, M.; Pun, I.; Podder, R.; Tomczak, A.; Hossain, A. Use of different eco-friendly management approaches for controlling root-knot nematode (Meloidogyne incognita L.) in tomato (Solanum lycopersicum L.). Thai J. Agric. Sci. 2021, 54, 89–103. [Google Scholar]
- Wang, J.; Zhai, L.; Ma, J.; Zhang, J.; Wang, G.G.; Liu, X.; Zhang, S.; Song, J.; Wu, Y. Comparative physiological mechanisms of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects on leaves and roots of Zelkova serrata. Mycorrhiza 2020, 30, 341–355. [Google Scholar] [CrossRef]
- Marro, N.; Caccia, M.; Doucet, M.E.; Cabello, M.; Becerra, A.; Lax, P. Mycorrhizas reduce tomato root penetration by false root-knot nematode Nacobbus aberrans. Appl. Soil Ecol. 2018, 124, 262–265. [Google Scholar] [CrossRef]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef]
- Palumbo, G.; Carfagna, S.; Stoleru, V.; Torino, V.; Romano, P.; Letizia, F.; Di Martino, C. Environmental sustainability fruit quality and production in mycorrhizal tomato plants without P fertilizing. Agron. Res. 2020, 18, 2535–2549. [Google Scholar]
- Fayaz, F.; Zahedi, M. Beneficial effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) nutritional status and tolerance indices under soil salinity stress. J. Plant Nutr. 2021, 45, 185–201. [Google Scholar] [CrossRef]
- Alaux, P.-L. Does the Arbuscular Mycorrhizal Fungus Rhizophagus Irregularis Mitigate Late Blight in Potato Plants? Presses Universitaires de Louvain: Louvain, Belgium, 2020. [Google Scholar]
- Ivanov, S.; Austin, J.; Berg, R.H.; Harrison, M.J. Extensive membrane systems at the host–arbuscular mycorrhizal fungus interface. Nat. Plants 2019, 5, 194–203. [Google Scholar] [CrossRef]
- Carella, P.; Gogleva, A.; Tomaselli, M.; Alfs, C.; Schornack, S. Phytophthora palmivora establishes tissue-specific intracellular infection structures in the earliest divergent land plant lineage. Proc. Natl. Acad. Sci. USA 2018, 115, E3846–E3855. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, J.; Xie, K.; Tian, Y.; Yan, A.; Liu, J.; Huang, Y.; Wang, S.; Zhu, Y.; Chen, A. A mycorrhiza-specific H+-ATPase is essential for arbuscule development and symbiotic phosphate and nitrogen uptake. Plant Cell Environ. 2020, 43, 1069–1083. [Google Scholar] [CrossRef]
- Mbodj, D.; Effa-Effa, B.; Kane, A.; Manneh, B.; Gantet, P.; Laplaze, L.; Diedhiou, A.; Grondin, A. Arbuscular mycorrhizal symbiosis in rice: Establishment, environmental control and impact on plant growth and resistance to abiotic stresses. Rhizosphere 2018, 8, 12–26. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. Reclamation of arid and semi-arid soils: The role of plant growth-promoting archaea and bacteria. Curr. Plant Biol. 2021, 25, 100173. [Google Scholar] [CrossRef]
- Kumar, K.; Karthika, K. Abiotic and biotic factors influencing soil health and/or soil degradation. In Soil Health; Springer: Berlin/Heidelberg, Germany, 2020; pp. 145–161. [Google Scholar]
- Dong, S.; Shang, Z.; Gao, J.; Boone, R.B. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 287, 106684. [Google Scholar] [CrossRef]
- Singla, A.; Sharma, T. Role of Plant Physiology in the Era of Second Green Revolution. Int. J. Plant Biotechnol. 2021, 7, 18–36. [Google Scholar]
- Sarker, M.N.I.; Wu, M.; Alam, G.M.; Islam, M.S. Role of climate smart agriculture in promoting sustainable agriculture: A systematic literature review. Int. J. Agric. Resour. Gov. Ecol. 2019, 15, 323–337. [Google Scholar] [CrossRef]
- Dhakal, B.; Kattel, R.R. Effects of global changes on ecosystems services of multiple natural resources in mountain agricultural landscapes. Sci. Total Environ. 2019, 676, 665–682. [Google Scholar] [CrossRef]
- Liu, C.; Xu, Y.; Lu, X.; Han, J. Trade-offs and driving forces of land use functions in ecologically fragile areas of northern Hebei Province: Spatiotemporal analysis. Land Use Policy 2021, 104, 105387. [Google Scholar] [CrossRef]
- Luiz, J.M. Economics, transitions, and traps in emerging markets. In The Oxford Handbook of Management in Emerging Markets; Oxford University Press: Oxford UK, 2019; p. 77. [Google Scholar]
- Aldy, J.E.; Stavins, R.N. The promise and problems of pricing carbon: Theory and experience. J. Environ. Dev. 2012, 21, 152–180. [Google Scholar] [CrossRef]
- Sandalow, D.; Aines, R.; Friedmann, J.; McCormick, C.; Sanchez, D.L. Biomass Carbon Removal and Storage (BiRCS) Roadmap; Lawrence Livermore National Lab. (LLNL): Livermore, CA, USA, 2021. [Google Scholar]
- Elkin-Koren, N.; Netanel, N.W. Transplanting fair use across the globe: A case study testing the credibility of US opposition. Hastings Law J. 2020, 72, 1121. [Google Scholar]
- De Pinto, A.; Cenacchi, N.; Kwon, H.-Y.; Koo, J.; Dunston, S. Climate smart agriculture and global food-crop production. PLoS ONE 2020, 15, e0231764. [Google Scholar] [CrossRef]
- Ejersa, M.T. The role of arbuscular mycorrhizal fungi on agricultural crop productivity and ecosystem service: A review. Int. J. Agrofor. Silvic. 2021, 9, 1–9. [Google Scholar]
- Messerli, P.; Murniningtyas, E.; Eloundou-Enyegue, P.; Foli, E.G.; Furman, E.; Glassman, A.; Hernández Licona, G.; Kim, E.M.; Lutz, W.; Moatti, J.-P. Global Sustainable Development Report 2019: The Future Is Now–Science for Achieving Sustainable Development; United Nations: New York, NY, USA, 2019. [Google Scholar]
- Mansour, K.M.M. Management of Natural Resources in Function of Developing Agricultural Production System in LIBYA. Ph.D. Thesis, University of Business Academy in Novi Sad, Novi Sad, Serbia, 2020. [Google Scholar]
- Rey, F.; Rivière, P.; Flipon, E.; de Buck, A.; Feher, J.; Constanzo, A.; Lazzaro, M. Frugal, multi-actor and decentralised cultivar evaluation models for organic agriculture: Methods, tools and guidelines. Breed. Seed Sect. Innov. Org. Food Syst. 2021, 117. [Google Scholar]
- Artmann, M. Assessment of soil sealing management responses, strategies, and targets toward ecologically sustainable urban land use management. Ambio 2014, 43, 530–541. [Google Scholar] [CrossRef]
- Favero-Longo, S.E.; Viles, H.A. A review of the nature, role and control of lithobionts on stone cultural heritage: Weighing-up and managing biodeterioration and bioprotection. World J. Microbiol. Biotechnol. 2020, 36, 100. [Google Scholar] [CrossRef] [PubMed]
- Bhantana, P.; Rana, M.S.; Sun, X.-c.; Moussa, M.G.; Saleem, M.H.; Syaifudin, M.; Shah, A.; Poudel, A.; Pun, A.B.; Bhat, M.A. Arbuscular mycorrhizal fungi and its major role in plant growth, zinc nutrition, phosphorous regulation and phytoremediation. Symbiosis 2021, 84, 19–37. [Google Scholar] [CrossRef]
- Swain, H.; Naik, S.K.; Mukherjee, A.K. Utilization of Arbuscular Mycorrhizal Fungi to Boom the Efficiency and Product Nature of Horticultural Crops. In Advances in Agricultural and Industrial Microbiology; Springer: Berlin/Heidelberg, Germany, 2022; pp. 119–130. [Google Scholar]
- Bagyaraj, D.J.; Sridhar, K.R.; Revanna, A. Arbuscular Mycorrhizal Fungi Influence Crop Productivity, Plant Diversity, and Ecosystem Services. In Fungal Diversity, Ecology and Control Management; Springer: Singapore, 2022; pp. 345–362. [Google Scholar]
- Ma, J.; Ullah, S.; Niu, A.; Liao, Z.; Qin, Q.; Xu, S.; Lin, C. Heavy metal pollution increases CH4 and decreases CO2 emissions due to soil microbial changes in a mangrove wetland: Microcosm experiment and field examination. Chemosphere 2021, 269, 128735. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, B.; Heinonsalo, J.; Simon, J. Mechanisms of carbon sequestration in highly organic ecosystems–importance of chemical ecology. ChemistryOpen 2020, 9, 464–469. [Google Scholar] [CrossRef]
- Ye, J.; An, N.; Chen, H.; Ying, Z.; Zhang, S.; Zhao, J. Performance and mechanism of carbon dioxide fixation by a newly isolated chemoautotrophic strain Paracoccus denitrificans PJ-1. Chemosphere 2020, 252, 126473. [Google Scholar] [CrossRef]
- Taub, D. Effects of rising atmospheric concentrations of carbon dioxide on plants. Nat. Educ. Knowl. 2010, 3, 21. [Google Scholar]
- Wielopolski, L. Geological carbon sequestration: A new approach for near-surface assurance monitoring. Carbon Capture Storage CO2 Manag. Technol. 2014, 2013, 105–120. [Google Scholar] [CrossRef]
- Tan, Z.; Liu, S.; Sohl, T.L.; Wu, Y.; Young, C.J. Ecosystem carbon stocks and sequestration potential of federal lands across the conterminous United States. Proc. Natl. Acad. Sci. USA 2015, 112, 12723–12728. [Google Scholar] [CrossRef]
- Ren, G.; Zhang, H.; Lin, X.; Zhu, J.; Jia, Z. Response of leaf endophytic bacterial community to elevated CO2 at different growth stages of rice plant. Front. Microbiol. 2015, 6, 855. [Google Scholar] [CrossRef]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Law, T.F.; De Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, C.D. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef]
- Pattnaik, S.; Mohapatra, B.; Gupta, A. Plant growth-promoting microbe mediated uptake of essential nutrients (Fe, P, K) for crop stress management: Microbe–soil–plant continuum. Front. Agron. 2021, 3, 689972. [Google Scholar] [CrossRef]
- Aghdam, S.A.; Brown, A.M.V. Deep learning approaches for natural product discovery from plant endophytic microbiomes. Environ. Microbiome 2021, 16, 6. [Google Scholar] [CrossRef]
- Rozpądek, P.; Wężowicz, K.; Nosek, M.; Ważny, R.; Tokarz, K.; Lembicz, M.; Miszalski, Z.; Turnau, K. The fungal endophyte Epichloë typhina improves photosynthesis efficiency of its host orchard grass (Dactylis glomerata). Planta 2015, 242, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Cregger, M.A.; Carper, D.L.; Christel, S.; Doktycz, M.J.; Labbé, J.; Michener, J.K.; Dove, N.C.; Johnston, E.R.; Moore, J.A.; Vélez, J.M. Plant–microbe interactions: From genes to ecosystems using Populus as a model system. Phytobiomes J. 2021, 5, 29–38. [Google Scholar] [CrossRef]
- Williams, A.; Pétriacq, P.; Beerling, D.J.; Cotton, T.A.; Ton, J. Impacts of atmospheric CO2 and soil nutritional value on plant responses to rhizosphere colonization by soil bacteria. Front. Plant Sci. 2018, 9, 1493. [Google Scholar] [CrossRef]
- Hayat, S.; Faraz, A.; Faizan, M. Root exudates: Composition and impact on plant–microbe interaction. In Biofilms in Plant and Soil Health; Wiley: Toronto, ON, Canada, 2017; pp. 179–193. [Google Scholar]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The chemistry of plant–microbe interactions in the rhizosphere and the potential for metabolomics to reveal signaling related to defense priming and induced systemic resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef]
- Habib, S.H.; Kausar, H.; Saud, H.M. Plant growth-promoting rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzymes. BioMed Res. Int. 2016, 2016, 6284547. [Google Scholar] [CrossRef]
- Mastan, A.; Bharadwaj, R.; Kushwaha, R.K.; Vivek Babu, C.S. Functional fungal endophytes in Coleus forskohlii regulate labdane diterpene biosynthesis for elevated forskolin accumulation in roots. Microb. Ecol. 2019, 78, 914–926. [Google Scholar] [CrossRef]
- Khanna, K.; Kohli, S.K.; Ohri, P.; Bhardwaj, R.; Al-Huqail, A.A.; Siddiqui, M.H.; Alosaimi, G.S.; Ahmad, P. Microbial fortification improved photosynthetic efficiency and secondary metabolism in Lycopersicon esculentum plants under Cd stress. Biomolecules 2019, 9, 581. [Google Scholar] [CrossRef]
- Wu, K.; Li, J.; Luo, J.; Liu, Y.; Song, Y.; Liu, N.; Rafiq, M.T.; Li, T. Effects of elevated CO2 and endophytic bacterium on photosynthetic characteristics and cadmium accumulation in Sedum alfredii. Sci. Total Environ. 2018, 643, 357–366. [Google Scholar] [CrossRef]
- Shukla, N.; Awasthi, R.; Rawat, L.; Kumar, J. Biochemical and physiological responses of rice (Oryza sativa L.) as influenced by Trichoderma harzianum under drought stress. Plant Physiol. Biochem. 2012, 54, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Rho, H.; Van Epps, V.; Wegley, N.; Doty, S.L.; Kim, S.-H. Salicaceae endophytes modulate stomatal behavior and increase water use efficiency in rice. Front. Plant Sci. 2018, 9, 188. [Google Scholar] [CrossRef]
- Jansson, C.; Faiola, C.; Wingler, A.; Zhu, X.-G.; Kravchenko, A.; De Graaff, M.-A.; Ogden, A.J.; Handakumbura, P.P.; Werner, C.; Beckles, D.M. Crops for carbon farming. Front. Plant Sci. 2021, 12, 636709. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Kracher, B.; Hiruma, K.; Münch, P.; Garrido-Oter, R.; Thon, M.; Weimann, A.; Damm, U.; Dallery, J.; Hainaut, M. Survival trade-offs in plant roots during colonization by closely related beneficial and pathogenic fungi. Nat. Commun. 2016, 7, 11362. [Google Scholar] [CrossRef] [PubMed]
- Gul Jan, F.; Hamayun, M.; Hussain, A.; Jan, G.; Iqbal, A.; Khan, A.; Lee, I.-J. An endophytic isolate of the fungus Yarrowia lipolytica produces metabolites that ameliorate the negative impact of salt stress on the physiology of maize. BMC Microbiol. 2019, 19, 3. [Google Scholar] [CrossRef]
- Hussain, T.; Li, J.; Feng, X.; Asrar, H.; Gul, B.; Liu, X. Salinity induced alterations in photosynthetic and oxidative regulation are ameliorated as a function of salt secretion. J. Plant Res. 2021, 134, 779–796. [Google Scholar] [CrossRef]
- Inada, H.; Kondo, T.; Akhtar, N.; Hoshino, D.; Yamaguchi, M.; Izuta, T. Relationship between cultivar difference in the sensitivity of net photosynthesis to ozone and reactive oxygen species scavenging system in Japanese winter wheat (Triticum aestivum). Physiol. Plant. 2012, 146, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.; Khadka, R.; Doni, F.; Uphoff, N. Benefits to plant health and productivity from enhancing plant microbial symbionts. Front. Plant Sci. 2021, 11, 610065. [Google Scholar] [CrossRef]
- Nie, M.; Bell, C.; Wallenstein, M.D.; Pendall, E. Increased plant productivity and decreased microbial respiratory C loss by plant growth-promoting rhizobacteria under elevated CO2. Sci. Rep. 2015, 5, 9212. [Google Scholar] [CrossRef] [PubMed]
- Khalloufi, M.; Martínez-Andújar, C.; Lachaâl, M.; Karray-Bouraoui, N.; Pérez-Alfocea, F.; Albacete, A. The interaction between foliar GA3 application and arbuscular mycorrhizal fungi inoculation improves growth in salinized tomato (Solanum lycopersicum L.) plants by modifying the hormonal balance. J. Plant Physiol. 2017, 214, 134–144. [Google Scholar] [CrossRef]
- Lin, J.; Wang, Y.; Sun, S.; Mu, C.; Yan, X. Effects of arbuscular mycorrhizal fungi on the growth, photosynthesis and photosynthetic pigments of Leymus chinensis seedlings under salt-alkali stress and nitrogen deposition. Sci. Total Environ. 2017, 576, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Abd_Allah, E.; Alqarawi, A.; Al Huqail, A.A.; Egamberdieva, D.; Wirth, S. Alleviation of cadmium stress in Solanum lycopersicum L. by arbuscular mycorrhizal fungi via induction of acquired systemic tolerance. Saudi J. Biol. Sci. 2016, 23, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Shi-Chu, L.; Yong, J.; Ma-Bo, L.; Wen-Xu, Z.; Nan, X.; Hui-hui, Z. Improving plant growth and alleviating photosynthetic inhibition from salt stress using AMF in alfalfa seedlings. J. Plant Interact. 2019, 14, 482–491. [Google Scholar] [CrossRef]
- Shahvali, R.; Shiran, B.; Ravash, R.; Fallahi, H.; Đeri, B.B. Effect of symbiosis with arbuscular mycorrhizal fungi on salt stress tolerance in GF677 (peach× almond) rootstock. Sci. Hortic. 2020, 272, 109535. [Google Scholar] [CrossRef]
- Parihar, M.; Rakshit, A.; Rana, K.; Tiwari, G.; Jatav, S.S. The effect of arbuscular mycorrhizal fungi inoculation in mitigating salt stress of pea (Pisum sativum L.). Commun. Soil Sci. Plant Anal. 2020, 51, 1545–1559. [Google Scholar] [CrossRef]
- Li, Z.; Wu, N.; Meng, S.; Wu, F.; Liu, T. Arbuscular mycorrhizal fungi (AMF) enhance the tolerance of Euonymus maackii Rupr. at a moderate level of salinity. PLoS ONE 2020, 15, e0231497. [Google Scholar] [CrossRef]
- Bijalwan, P.; Jeddi, K.; Saini, I.; Sharma, M.; Kaushik, P.; Hessini, K. Mitigation of saline conditions in watermelon with mycorrhiza and silicon application. Saudi J. Biol. Sci. 2021, 28, 3678–3684. [Google Scholar] [CrossRef]
- Klinsukon, C.; Lumyong, S.; Kuyper, T.W.; Boonlue, S. Colonization by arbuscular mycorrhizal fungi improves salinity tolerance of eucalyptus (Eucalyptus camaldulensis) seedlings. Sci. Rep. 2021, 11, 4362. [Google Scholar] [CrossRef]
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved photosynthetic efficacy of maize (Zea mays) plants with arbuscular mycorrhizal fungi (AMF) under high temperature stress. J. Photochem. Photobiol. B Biol. 2018, 180, 149–154. [Google Scholar] [CrossRef]
- Cabral, C.; Ravnskov, S.; Tringovska, I.; Wollenweber, B. Arbuscular mycorrhizal fungi modify nutrient allocation and composition in wheat (Triticum aestivum L.) subjected to heat-stress. Plant Soil 2016, 408, 385–399. [Google Scholar] [CrossRef]
- Liu, N.; Chen, X.; Song, F.; Liu, F.; Liu, S.; Zhu, X. Effects of arbuscular mycorrhiza on growth and nutrition of maize plants under low temperature stress. Philipp. Agric. Sci. 2016, 99, 246–252. [Google Scholar]
- Zhu, X.; Song, F.; Liu, F. Altered amino acid profile of arbuscular mycorrhizal maize plants under low temperature stress. J. Plant Nutr. Soil Sci. 2016, 179, 186–189. [Google Scholar] [CrossRef]
- Chu, X.; Fu, J.; Sun, Y.; Xu, Y.; Miao, Y.; Xu, Y.; Hu, T. Effect of arbuscular mycorrhizal fungi inoculation on cold stress-induced oxidative damage in leaves of Elymus nutans Griseb. S. Afr. J. Bot. 2016, 104, 21–29. [Google Scholar] [CrossRef]
- Hajiboland, R.; Joudmand, A.; Aliasgharzad, N.; Tolrá, R.; Poschenrieder, C. Arbuscular mycorrhizal fungi alleviate low-temperature stress and increase freezing resistance as a substitute for acclimation treatment in barley. Crop Pasture Sci. 2019, 70, 218–233. [Google Scholar] [CrossRef]
- Ma, J.; Janoušková, M.; Ye, L.; Bai, L.; Dong, R.; Yan, Y.; Yu, X.; Zou, Z.; Li, Y.; He, C. Role of arbuscular mycorrhiza in alleviating the effect of cold on the photosynthesis of cucumber seedlings. Photosynthetica 2019, 57, 86–95. [Google Scholar] [CrossRef]
- Nogales, A.; Ribeiro, H.; Nogales-Bueno, J.; Hansen, L.D.; Gonçalves, E.F.; Coito, J.L.; Rato, A.E.; Peixe, A.; Viegas, W.; Cardoso, H. Response of Mycorrhizal’Touriga Nacional ‘Variety Grapevines to High Temperatures Measured by Calorespirometry and Near-Infrared Spectroscopy. Plants 2020, 9, 1499. [Google Scholar] [CrossRef]
- Reva, M.; Cano, C.; Herrera, M.-A.; Bago, A. Arbuscular mycorrhizal inoculation enhances endurance to severe heat stress in three horticultural crops. HortScience 2021, 56, 396–406. [Google Scholar] [CrossRef]
- Mirshad, P.; Puthur, J.T. Arbuscular mycorrhizal association enhances drought tolerance potential of promising bioenergy grass (Saccharum arundinaceum retz.). Environ. Monit. Assess. 2016, 188, 425. [Google Scholar] [CrossRef]
- Rani, B. Effect of Arbuscular Mycorrhiza Fungi on Biochemical Parameters in Wheat (Triticum aestivum L.) Under Drought Conditions [With CD Copy]; Biochemistry, CCSHAU: Hisar, India, 2016. [Google Scholar]
- Yooyongwech, S.; Samphumphuang, T.; Tisarum, R.; Theerawitaya, C.; Cha-um, S. Arbuscular mycorrhizal fungi (AMF) improved water deficit tolerance in two different sweet potato genotypes involves osmotic adjustments via soluble sugar and free proline. Sci. Hortic. 2016, 198, 107–117. [Google Scholar] [CrossRef]
- Tallapragada, P.; Dikshit, R.; Seshagiri, S. Influence of Rhizophagus spp. and Burkholderia seminalis on the growth of tomato (Lycopersicon esculatum) and bell pepper (Capsicum annuum) under drought stress. Commun. Soil Sci. Plant Anal. 2016, 47, 1975–1984. [Google Scholar] [CrossRef]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the impact of arbuscular mycorrhizal symbiosis on tomato tolerance to water stress. Plant Physiol. 2016, 171, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Pandey, S. Role of arbuscular mycorrhizal fungi on plant growth and reclamation of barren soil with wheat (Triticum aestivum L.) crop. Int. J. Soil Sci 2016, 12, 25–31. [Google Scholar] [CrossRef]
- Pedranzani, H.; Rodríguez-Rivera, M.; Gutiérrez, M.; Porcel, R.; Hause, B.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis regulates physiology and performance of Digitaria eriantha plants subjected to abiotic stresses by modulating antioxidant and jasmonate levels. Mycorrhiza 2016, 26, 141–152. [Google Scholar] [CrossRef]
- Goicoechea, N.; Antolín, M.C. Increased nutritional value in food crops. Microb. Biotechnol. 2017, 10, 1004–1007. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.; Lou, X.; Tang, M. Mycorrhizal and non-mycorrhizal Medicago truncatula roots exhibit differentially regulated NADPH oxidase and antioxidant response under Pb stress. Environ. Exp. Bot. 2019, 164, 10–19. [Google Scholar] [CrossRef]
- Zhang, F.; Jia-Dong, H.; Qiu-Dan, N.; Qiang-Sheng, W.; Ying-Ning, Z. Enhancement of drought tolerance in trifoliate orange by mycorrhiza: Changes in root sucrose and proline metabolisms. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 270–276. [Google Scholar] [CrossRef]
- Wu, H.-H.; Zou, Y.-N.; Rahman, M.M.; Ni, Q.-D.; Wu, Q.-S. Mycorrhizas alter sucrose and proline metabolism in trifoliate orange exposed to drought stress. Sci. Rep. 2017, 7, 42389. [Google Scholar] [CrossRef]
- Aalipour, H.; Nikbakht, A.; Etemadi, N.; Rejali, F.; Soleimani, M. Biochemical response and interactions between arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria during establishment and stimulating growth of Arizona cypress (Cupressus arizonica G.) under drought stress. Sci. Hortic. 2020, 261, 108923. [Google Scholar] [CrossRef]
- Hu, Y.; Xie, W.; Chen, B. Arbuscular mycorrhiza improved drought tolerance of maize seedlings by altering photosystem II efficiency and the levels of key metabolites. Chem. Biol. Technol. Agric. 2020, 7, 20. [Google Scholar] [CrossRef]
- Boutasknit, A.; Baslam, M.; Ait-El-Mokhtar, M.; Anli, M.; Ben-Laouane, R.; Douira, A.; El Modafar, C.; Mitsui, T.; Wahbi, S.; Meddich, A. Arbuscular mycorrhizal fungi mediate drought tolerance and recovery in two contrasting carob (Ceratonia siliqua L.) ecotypes by regulating stomatal, water relations, and (in) organic adjustments. Plants 2020, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Meng, P.; Feng, H.; Wang, C. Effects of arbuscular mycorrhizal fungi on growth and physiological performance of Catalpa bungei CA Mey. under drought stress. Forests 2020, 11, 1117. [Google Scholar] [CrossRef]
- Liao, X.; Chen, J.; Guan, R.; Liu, J.; Sun, Q. Two arbuscular mycorrhizal fungi alleviates drought stress and improves plant growth in Cinnamomum migao seedlings. Mycobiology 2021, 49, 396–405. [Google Scholar] [CrossRef]
- Gholinezhad, E.; Darvishzadeh, R. Influence of arbuscular mycorrhiza fungi and drought stress on fatty acids profile of sesame (Sesamum indicum L.). Field Crops Res. 2021, 262, 108035. [Google Scholar] [CrossRef]
- Jiang, Q.-Y.; Zhuo, F.; Long, S.-H.; Zhao, H.-D.; Yang, D.-J.; Ye, Z.-H.; Li, S.-S.; Jing, Y.-X. Can arbuscular mycorrhizal fungi reduce Cd uptake and alleviate Cd toxicity of Lonicera japonica grown in Cd-added soils? Sci. Rep. 2016, 6, 21805. [Google Scholar] [CrossRef]
- Garg, N.; Singh, S. Arbuscular mycorrhiza Rhizophagus irregularis and silicon modulate growth, proline biosynthesis and yield in Cajanus cajan L. Millsp.(pigeonpea) genotypes under cadmium and zinc stress. J. Plant Growth Regul. 2018, 37, 46–63. [Google Scholar] [CrossRef]
- Liu, L.; Li, J.; Yue, F.; Yan, X.; Wang, F.; Bloszies, S.; Wang, Y. Effects of arbuscular mycorrhizal inoculation and biochar amendment on maize growth, cadmium uptake and soil cadmium speciation in Cd-contaminated soil. Chemosphere 2018, 194, 495–503. [Google Scholar] [CrossRef]
- Abdelhameed, R.E.; Metwally, R.A. Alleviation of cadmium stress by arbuscular mycorrhizal symbiosis. Int. J. Phytoremediation 2019, 21, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Ai, S.; Chen, K.; Wang, X. Arbuscular mycorrhiza augments cadmium tolerance in soybean by altering accumulation and partitioning of nutrient elements, and related gene expression. Ecotoxicol. Environ. Saf. 2019, 171, 231–239. [Google Scholar] [CrossRef]
- Sayın, F.; Khalvati, M.; Erdinçler, A. Effects of sewage sludge application and arbuscular mycorrhizal fungi (G. mosseae and G. intraradices) interactions on the heavy metal phytoremediation in chrome mine tailings. Front. Plant Sci. 2019, 112, 217–224. [Google Scholar]
- Singh, G.; Pankaj, U.; Chand, S.; Verma, R.K. Arbuscular mycorrhizal fungi-assisted phytoextraction of toxic metals by Zea mays L. from tannery sludge. Soil Sediment Contam. Int. J. 2019, 28, 729–746. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, Y.; Xiao, Z.; Ban, Y.; Belvett, N. Positive effects of Funneliformis mosseae inoculation on reed seedlings under water and TiO2 nanoparticles stresses. World J. Microbiol. Biotechnol. 2019, 35, 81. [Google Scholar] [CrossRef] [PubMed]
- Zhan, F.; Li, B.; Jiang, M.; Li, T.; He, Y.; Li, Y.; Wang, Y. Effects of arbuscular mycorrhizal fungi on the growth and heavy metal accumulation of bermudagrass [Cynodon dactylon (L.) Pers.] grown in a lead–zinc mine wasteland. Int. J. Phytoremediation 2019, 21, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, M.; Li, Y.; Che, Y.; Xiao, Y. Effects of arbuscular mycorrhizal fungi, biochar and cadmium on the yield and element uptake of Medicago sativa. Sci. Total Environ. 2019, 655, 1150–1158. [Google Scholar] [CrossRef]
- Wu, J.T.; Wang, L.; Zhao, L.; Huang, X.C.; Ma, F. Arbuscular mycorrhizal fungi effect growth and photosynthesis of Phragmites australis (Cav.) Trin ex. Steudel under copper stress. Plant Biol. 2020, 22, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Crossay, T.; Cavaloc, Y.; Majorel, C.; Redecker, D.; Medevielle, V.; Amir, H. Combinations of different arbuscular mycorrhizal fungi improve fitness and metal tolerance of sorghum in ultramafic soil. Rhizosphere 2020, 14, 100204. [Google Scholar] [CrossRef]
- Wang, X.-X.; Wang, X.; Sun, Y.; Cheng, Y.; Liu, S.; Chen, X.; Feng, G.; Kuyper, T.W. Arbuscular mycorrhizal fungi negatively affect nitrogen acquisition and grain yield of maize in a N deficient soil. Front. Microbiol. 2018, 9, 418. [Google Scholar] [CrossRef] [PubMed]
- Manjunatha, H.; Nirmalnath, P.J.; Chandranath, H.; Ammanna, S.; Jagadeesh, K. Field evalualtion of native arbuscular mycorrhizal fungi in the management of Striga in sugarcane (Saccharum officinarum L.). J. Pharmacogn. Phytochem. 2018, 7, 2496–2500. [Google Scholar]
- Liu, Y.; Feng, X.; Gao, P.; Li, Y.; Christensen, M.J.; Duan, T. Arbuscular mycorrhiza fungi increased the susceptibility of Astragalus adsurgens to powdery mildew caused by Erysiphe pisi. Mycology 2018, 9, 223–232. [Google Scholar] [CrossRef]
- Kumari, S.M.P.; Srimeena, N. Arbuscular mycorrhizal fungi (AMF) induced defense factors against the damping-off disease pathogen, Pythium aphanidermatum in chilli (Capsicum annum). Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2243–2248. [Google Scholar] [CrossRef]
- Kumari, S.M.P.; Prabina, B.J. Protection of tomato, Lycopersicon esculentum from wilt pathogen, Fusarium oxysporum f. sp. lycopersici by arbuscular mycorrhizal fungi, Glomus sp. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1368–1378. [Google Scholar] [CrossRef]
- Spagnoletti, F.N.; Cornero, M.; Chiocchio, V.; Lavado, R.S.; Roberts, I.N. Arbuscular mycorrhiza protects soybean plants against Macrophomina phaseolina even under nitrogen fertilization. Eur. J. Plant Pathol. 2020, 156, 839–849. [Google Scholar] [CrossRef]
- Aylward, J.; Roets, F.; Dreyer, L.L.; Wingfield, M.J. Unseen fungal biodiversity and complex inter-organismal interactions in Protea flower heads. Fungal Biol. Rev. 2023, 45, 100317. [Google Scholar] [CrossRef]
- Frąc, M.; Jędryczka, M.; Hannula, E.S. Soil fungal biodiversity for plant and soil health, volume II. Front. Microbiol. 2023, 14, 1170312. [Google Scholar] [CrossRef]
- Vandegrift, R.; Newman, D.; Dentinger, B.; Batallas-Molina, R.; Dueñas, N.; Flores, J.; Goyes, P.; Jenkinson, T.; McAlpine, J.; Navas, D. Richer than Gold: The fungal biodiversity of Reserva Los Cedros, a threatened Andean cloud forest. Bot. Stud. 2023, 64, 17. [Google Scholar] [CrossRef]
- Panstruga, R.; Antonin, W.; Lichius, A. Looking outside the box: A comparative cross-kingdom view on the cell biology of the three major lineages of eukaryotic multicellular life. Cell. Mol. Life Sci. 2023, 80, 198. [Google Scholar] [CrossRef]
- Tang, S.-K.; Zhi, X.-Y.; Zhang, Y.; Makarova, K.S.; Liu, B.-B.; Zheng, G.-S.; Zhang, Z.-P.; Zheng, H.-J.; Wolf, Y.I.; Zhao, Y.-R. Cellular differentiation into hyphae and spores in halophilic archaea. Nat. Commun. 2023, 14, 1827. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, X.; Wang, E. Mycorrhizal symbiosis in plant growth and stress adaptation: From genes to ecosystems. Annu. Rev. Plant Biol. 2023, 74, 569–607. [Google Scholar] [CrossRef]
- Tajuddin, R.Y.; Asfar, M.; Dirpan, A. An overview-is mycelium composite foam can be used as a food packaging. AIP Conf. Proc. 2023, 2596, 040010. [Google Scholar]
- Sun, X.; Shi, J.; Ding, G. Combined effects of arbuscular mycorrhiza and drought stress on plant growth and mortality of forage sorghum. Appl. Soil Ecol. 2017, 119, 384–391. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef]
- Redecker, D.; Schüßler, A.; Stockinger, H.; Stürmer, S.L.; Morton, J.B.; Walker, C. An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 2013, 23, 515–531. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Bago, B.; Pfeffer, P.; Shachar-Hill, Y. Carbon transport and metabolism in arbuscular mycorrhiza. Plant Physiol 2000, 124, 949–957. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Gianinazzi, S.; Gollotte, A.; Binet, M.-N.; van Tuinen, D.; Redecker, D.; Wipf, D. Agroecology: The key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 2010, 20, 519–530. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant growth-promoting rhizobacteria for sustainable agricultural production. Microorganisms 2023, 11, 1088. [Google Scholar] [CrossRef] [PubMed]
- Astapati, A.D.; Nath, S. The complex interplay between plant-microbe and virus interactions in sustainable agriculture: Harnessing phytomicrobiomes for enhanced soil health, designer plants, resource use efficiency, and food security. Crop Des. 2023, 2, 100028. [Google Scholar] [CrossRef]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef]
- Li, W.; Yao, J.; He, C.; Ren, Y.; Zhao, L.; He, X. The synergy of dark septate endophytes and organic residue on Isatis indigotica growth and active ingredients accumulation under drought stress. Ind. Crops Prod. 2023, 203, 117147. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, J.; Cui, L.; Tang, Z.; Ci, D.; Zou, X.; Zhang, X.; Yu, X.; Wang, Y.; Si, T. The multifaceted roles of Arbuscular Mycorrhizal Fungi in peanut responses to salt, drought, and cold stress. BMC Plant Biol. 2023, 23, 36. [Google Scholar] [CrossRef]
- Díaz-Urbano, M.; Goicoechea, N.; Velasco, P.; Poveda, J. Development of agricultural bio-inoculants based on mycorrhizal fungi and endophytic filamentous fungi: Co-inoculants for improve plant-physiological responses in sustainable agriculture. Biol. Control 2023, 182, 105223. [Google Scholar] [CrossRef]
- Li, G.; Tang, X.; Hou, Q.; Li, T.; Xie, H.; Lu, Z.; Zhang, T.; Liao, Y.; Wen, X. Response of soil organic carbon fractions to legume incorporation into cropping system and the factors affecting it: A global meta-analysis. Agric. Ecosyst. Environ. 2023, 342, 108231. [Google Scholar] [CrossRef]
- Medina-Sauza, R.M.; Álvarez-Jiménez, M.; Delhal, A.; Reverchon, F.; Blouin, M.; Guerrero-Analco, J.A.; Cerdán, C.R.; Guevara, R.; Villain, L.; Barois, I. Earthworms building up soil microbiota, a review. Front. Environ. Sci. 2019, 7, 81. [Google Scholar] [CrossRef]
- Janowski, D.; Leski, T. Factors in the distribution of mycorrhizal and soil fungi. Diversity 2022, 14, 1122. [Google Scholar] [CrossRef]
- Ray, P.; Lakshmanan, V.; Labbé, J.L.; Craven, K.D. Microbe to microbiome: A paradigm shift in the application of microorganisms for sustainable agriculture. Front. Microbiol. 2020, 11, 622926. [Google Scholar] [CrossRef]
- Bhupenchandra, I.; Chongtham, S.K.; Devi, E.L.; Choudhary, A.K.; Salam, M.D.; Sahoo, M.R.; Bhutia, T.L.; Devi, S.H.; Thounaojam, A.S.; Behera, C. Role of biostimulants in mitigating the effects of climate change on crop performance. Front. Plant Sci. 2022, 13, 967665. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Jajoo, A. Arbuscular mycorrhizal fungi protects maize plants from high temperature stress by regulating photosystem II heterogeneity. Ind. Crops Prod. 2020, 143, 111934. [Google Scholar] [CrossRef]
- Etikala, B.; Adimalla, N.; Madhav, S.; Somagouni, S.G.; Keshava Kiran Kumar, P. Salinity Problems in Groundwater and Management Strategies in Arid and Semiarid Regions. In Groundwater Geochemistry: Pollution and Remediation Methods; Wiley: Toronto, ON, Canada, 2021; pp. 42–56. [Google Scholar]
- Lin, C.; Wang, Y.; Liu, M.; Li, Q.; Xiao, W.; Song, X. Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia lanceolata). Sci. Rep. 2020, 10, 12260. [Google Scholar] [CrossRef]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Baron, N.C.; Rigobelo, E.C. Endophytic fungi: A tool for plant growth promotion and sustainable agriculture. Mycology 2022, 13, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, M.; Kumar, V.; Vyas, P.; Dhaliwal, H.S.; Saxena, A.K. Microbial biofertilizers: Bioresources and eco-friendly technologies for agricultural and environmental sustainability. Biocatal. Agric. Biotechnol. 2020, 23, 101487. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Edlinger, A.; Garland, G.; Hartman, K.; Banerjee, S.; Degrune, F.; García-Palacios, P.; Hallin, S.; Valzano-Held, A.; Herzog, C.; Jansa, J. Agricultural management and pesticide use reduce the functioning of beneficial plant symbionts. Nat. Ecol. Evol. 2022, 6, 1145–1154. [Google Scholar] [CrossRef]
- Babikova, Z.; Gilbert, L.; Bruce, T.; Dewhirst, S.Y.; Pickett, J.A.; Johnson, D. Arbuscular mycorrhizal fungi and aphids interact by changing host plant quality and volatile emission. Funct. Ecol. 2014, 28, 375–385. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef]
- Elhamouly, N.A.; Hewedy, O.A.; Zaitoon, A.; Miraples, A.; Elshorbagy, O.T.; Hussien, S.; El-Tahan, A.; Peng, D. The hidden power of secondary metabolites in plant-fungi interactions and sustainable phytoremediation. Front. Plant Sci. 2022, 13, 1044896. [Google Scholar] [CrossRef]
- Filion, M.; St-Arnaud, M.; Jabaji-Hare, S. Quantification of Fusarium solani f. sp. phaseoli in mycorrhizal bean plants and surrounding mycorrhizosphere soil using real-time polymerase chain reaction and direct isolations on selective media. Phytopathology 2003, 93, 229–235. [Google Scholar] [CrossRef]
- Nanjundappa, A.; Bagyaraj, D.J.; Saxena, A.K.; Kumar, M.; Chakdar, H. Interaction between arbuscular mycorrhizal fungi and Bacillus spp. in soil enhancing growth of crop plants. Fungal Biol. Biotechnol. 2019, 6, 23. [Google Scholar] [CrossRef]
- George, N.P.; Ray, J.G. The inevitability of arbuscular mycorrhiza for sustainability in organic agriculture—A critical review. Front. Sustain. Food Syst. 2023, 7, 1124688. [Google Scholar] [CrossRef]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial services of arbuscular mycorrhizal fungi–from ecology to application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef]
- Khaliq, A.; Perveen, S.; Alamer, K.H.; Zia Ul Haq, M.; Rafique, Z.; Alsudays, I.M.; Althobaiti, A.T.; Saleh, M.A.; Hussain, S.; Attia, H. Arbuscular Mycorrhizal Fungi Symbiosis to Enhance Plant–Soil Interaction. Sustainability 2022, 14, 7840. [Google Scholar] [CrossRef]
- Loo, W.T.; Chua, K.-O.; Mazumdar, P.; Cheng, A.; Osman, N.; Harikrishna, J.A. Arbuscular Mycorrhizal Symbiosis: A Strategy for Mitigating the Impacts of Climate Change on Tropical Legume Crops. Plants 2022, 11, 2875. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Chen, G.; Zhao, J.; Sinha, S.; Zhang, W. AMF-Placer 2.0: Open Source Timing-driven Analytical Mixed-size Placer for Large-scale Heterogeneous FPGA. arXiv 2022, arXiv:2210.08682. [Google Scholar]
- Islam, M.A. Effects of Arbuscular Mycorrhizal Fungi on Plant Growth, Yield, and Plant-Parasitic Nematode Control in Organic Tomato Production: A Field Study and a Systematic Review; Hochschule Rhein-Waal: Westfalen, Germany, 2022. [Google Scholar]
- Dhiman, M.; Sharma, L.; Kaushik, P.; Singh, A.; Sharma, M.M. Mycorrhiza: An Ecofriendly Bio-Tool for Better Survival of Plants in Nature. Sustainability 2022, 14, 10220. [Google Scholar] [CrossRef]
- Watts-Williams, S.J. Track and trace: How soil labelling techniques have revealed the secrets of resource transport in the arbuscular mycorrhizal symbiosis. Mycorrhiza 2022, 32, 257–267. [Google Scholar] [CrossRef]
- Gao, X.; Guo, H.; Zhang, Q.; Guo, H.; Zhang, L.; Zhang, C.; Gou, Z.; Liu, Y.; Wei, J.; Chen, A. Arbuscular mycorrhizal fungi (AMF) enhanced the growth, yield, fiber quality and phosphorus regulation in upland cotton (Gossypium hirsutum L.). Sci. Rep. 2020, 10, 2084. [Google Scholar] [CrossRef]
- Adisa, I.O.; Pullagurala, V.L.R.; Peralta-Videa, J.R.; Dimkpa, C.O.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Recent advances in nano-enabled fertilizers and pesticides: A critical review of mechanisms of action. Environ. Sci. Nano 2019, 6, 2002–2030. [Google Scholar] [CrossRef]
- Reicosky, D.; Calegari, A.; dos Santos, D.R.; Tiecher, T. Cover Crop Mixes for Diversity, Carbon and Conservation Agriculture. In Cover Crops and Sustainable Agriculture; CRC Press: Boca Raton, FL, USA, 2021; pp. 169–208. [Google Scholar]
- Kaur, J. Effects of Arbuscular Mycorrhizal Fungi (AMF) on Growth and Herbivore Defenses in Sorghum Sudangrass (Sorghum X drummondii). Master’s Thesis, The University of Texas Rio Grande Valley, Rio Grande Valley, TX, USA, 2020. [Google Scholar]
- Salomon, M.; Watts-Williams, S.; McLaughlin, M.; Brien, C.; Jewell, N.; Berger, B.; Cavagnaro, T. Evaluation of commercial composts and potting mixes and their ability to support arbuscular mycorrhizal fungi with maize (Zea mays) as host plant. Waste Manag. 2021, 134, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, W.-B.; Xing, L.-J.; Guo, S.-X. Effect of arbuscular mycorrhizal fungi (AMF) and plant growth-promoting rhizobacteria (PGPR) on microorganism of phenanthrene and pyrene contaminated soils. Int. J. Phytoremediation 2022, 25, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Konvalinková, T.; Püschel, D.; Řezáčová, V.; Gryndlerová, H.; Jansa, J. Carbon flow from plant to arbuscular mycorrhizal fungi is reduced under phosphorus fertilization. Plant Soil 2017, 419, 319–333. [Google Scholar] [CrossRef]
- Püschel, D.; Bitterlich, M.; Rydlová, J.; Jansa, J. Facilitation of plant water uptake by an arbuscular mycorrhizal fungus: A Gordian knot of roots and hyphae. Mycorrhiza 2020, 30, 299–313. [Google Scholar] [CrossRef]
- Van’t Padje, A.; Werner, G.D.; Kiers, E.T. Mycorrhizal fungi control phosphorus value in trade symbiosis with host roots when exposed to abrupt ‘crashes’ and ‘booms’ of resource availability. New Phytol. 2021, 229, 2933–2944. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Yousaf, S.; Sarwar, N.; Hussain, S. Zinc biofortification of cereals—Role of phosphorus and other impediments in alkaline calcareous soils. Environ. Geochem. Health 2019, 41, 2365–2379. [Google Scholar] [CrossRef]
- Wall, C.B.; Egan, C.P.; Swift, S.I.; Hynson, N.A. Three decades post-reforestation has not led to the reassembly of arbuscular mycorrhizal fungal communities associated with remnant primary forests. Mol. Ecol. 2020, 29, 4234–4247. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tienda, J.; Valderas, A.; Camañes, G.; García-Agustín, P.; Ferrol, N. Kinetics of NH 4+ uptake by the arbuscular mycorrhizal fungus Rhizophagus irregularis. Mycorrhiza 2012, 22, 485–491. [Google Scholar] [CrossRef]
- Jenks, G.K. Restoring the natural functional capacity of coastal dune ecosystems: Utilising research records for New Zealand littoral refurbishment as a proxy for analogous global responses. J. Coast. Conserv. 2018, 22, 623–665. [Google Scholar] [CrossRef]
- Xie, L.; Bi, Y.; Ma, S.; Shang, J.; Hu, Q.; Christie, P. Combined inoculation with dark septate endophytes and arbuscular mycorrhizal fungi: Synergistic or competitive growth effects on maize? BMC Plant Biol. 2021, 21, 498. [Google Scholar] [CrossRef]
- Schüßler, A.; Martin, H.; Cohen, D.; Fitz, M.; Wipf, D. Arbuscular mycorrhiza: Studies on the geosiphon symbiosis lead to the characterization of the first glomeromycotan sugar transporter. Plant Signal. Behav. 2007, 2, 431–434. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Ashwath, N. Glomeromycotan mycorrhizal fungi from tropical Australia III. Measuring diversity in natural and disturbed habitats. Plant Soil 2013, 370, 419–433. [Google Scholar] [CrossRef]
- Paranavithana, T.; Marasinghe, S.; Perera, G.; Ratnayake, R. Effects of crop rotation on enhanced occurrence of arbuscular mycorrhizal fungi and soil carbon stocks of lowland paddy fields in seasonaly dry tropics. Paddy Water Environ. 2021, 19, 217–226. [Google Scholar] [CrossRef]
- Burridge, J.D.; Findeis, J.L.; Jochua, C.N.; Miguel, M.A.; Mubichi-Kut, F.M.; Quinhentos, M.L.; Xerinda, S.A.; Lynch, J.P. A case study on the efficacy of root phenotypic selection for edaphic stress tolerance in low-input agriculture: Common bean breeding in Mozambique. Field Crops Res. 2019, 244, 107612. [Google Scholar] [CrossRef]
- An, J.; Zeng, T.; Ji, C.; de Graaf, S.; Zheng, Z.; Xiao, T.T.; Deng, X.; Xiao, S.; Bisseling, T.; Limpens, E. A Medicago truncatula SWEET transporter implicated in arbuscule maintenance during arbuscular mycorrhizal symbiosis. New Phytol. 2019, 224, 396–408. [Google Scholar] [CrossRef]
- Meena, R.S.; Vijayakumar, V.; Yadav, G.S.; Mitran, T. Response and interaction of Bradyrhizobium japonicum and arbuscular mycorrhizal fungi in the soybean rhizosphere. Plant Growth Regul. 2018, 84, 207–223. [Google Scholar] [CrossRef]
- Borowicz, V.A. Do arbuscular mycorrhizal fungi alter plant–pathogen relations? Ecology 2001, 82, 3057–3068. [Google Scholar]
- Bennett, A.E.; Classen, A.T. Climate change influences mycorrhizal fungal–plant interactions, but conclusions are limited by geographical study bias. Ecology 2020, 101, e02978. [Google Scholar] [CrossRef]
- Kazerooni, E.A.; Maharachchikumbura, S.S.; Al-Sadi, A.M.; Rashid, U.; Kang, S.-M.; Lee, I.-J. Actinomucor elegans and Podospora bulbillosa Positively Improves Endurance to Water Deficit and Salinity Stresses in Tomato Plants. J. Fungi 2022, 8, 785. [Google Scholar] [CrossRef]
- Liu, M.; Shen, Y.; Li, Q.; Xiao, W.; Song, X. Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen and phosphorus additions affects leaf C: N: P stoichiometry in Chinese fir (Cunninghamia lanceolata) forests. Plant Soil 2021, 461, 421–440. [Google Scholar] [CrossRef]
- Dubey, S.P.; Lahtinen, M.; Sillanpää, M. Tansy fruit mediated greener synthesis of silver and gold nanoparticles. Process Biochem. 2010, 45, 1065–1071. [Google Scholar] [CrossRef]
- Pearce, L.M. Critical Histories for Ecological Restoration. Ph.D. Thesis, The Australian National University (Australia), Canberra, Australia, 2019. [Google Scholar]
- Delavaux, C.S.; Smith-Ramesh, L.M.; Kuebbing, S.E. Beyond nutrients: A meta-analysis of the diverse effects of arbuscular mycorrhizal fungi on plants and soils. Ecology 2017, 98, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A.; Storer, K. Arbuscular mycorrhiza and nitrogen: Implications for individual plants through to ecosystems. Plant Soil 2015, 386, 1–19. [Google Scholar] [CrossRef]
- Liu, H.; Liang, C.; Ai, Z.; Zhang, J.; Wu, Y.; Xu, H.; Xue, S.; Liu, G. Plant-mycorrhizae association affects plant diversity, biomass, and soil nutrients along temporal gradients of natural restoration after farmland abandonment in the Loess Plateau, China. Land Degrad. Dev. 2019, 30, 1677–1690. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, Y.; Shi, Z.; Feng, G. Arbuscular mycorrhizal fungi can accelerate the restoration of degraded spring grassland in Central Asia. Rangel. Ecol. Manag. 2012, 65, 426–432. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zou, Y.-N.; Shu, B.; Wu, Q.-S. Deciphering molecular mechanisms regarding enhanced drought tolerance in plants by arbuscular mycorrhizal fungi. Sci. Hortic. 2023, 308, 111591. [Google Scholar] [CrossRef]
- Huang, D.; Ma, M.; Wang, Q.; Zhang, M.; Jing, G.; Li, C.; Ma, F. Arbuscular mycorrhizal fungi enhanced drought resistance in apple by regulating genes in the MAPK pathway. Plant Physiol. Biochem. 2020, 149, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Bi, Y.; Zhang, J.; Gong, Y.; Yang, H. Arbuscular mycorrhizal fungi promote the growth of plants in the mining associated clay. Sci. Rep. 2020, 10, 2663. [Google Scholar] [CrossRef] [PubMed]
- Benaffari, W.; Boutasknit, A.; Anli, M.; Ait-El-Mokhtar, M.; Ait-Rahou, Y.; Ben-Laouane, R.; Ben Ahmed, H.; Mitsui, T.; Baslam, M.; Meddich, A. The native arbuscular mycorrhizal fungi and vermicompost-based organic amendments enhance soil fertility, growth performance, and the drought stress tolerance of quinoa. Plants 2022, 11, 393. [Google Scholar] [CrossRef]
- Li, J.; Meng, B.; Chai, H.; Yang, X.; Song, W.; Li, S.; Lu, A.; Zhang, T.; Sun, W. Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. Front. Plant Sci. 2019, 10, 499. [Google Scholar] [CrossRef]
- Zhu, L.; Bai, Y.; Zhang, L.; Wu, G.; Si, W.; Wang, A. Water-land-food nexus for sustainable agricultural development in main grain-producing areas of North China Plain. Foods 2022, 12, 712. [Google Scholar] [CrossRef]
- Sagar, A.; Rathore, P.; Ramteke, P.W.; Ramakrishna, W.; Reddy, M.S.; Pecoraro, L. Plant growth promoting rhizobacteria, arbuscular mycorrhizal fungi and their synergistic interactions to counteract the negative effects of saline soil on agriculture: Key macromolecules and mechanisms. Microorganisms 2021, 9, 1491. [Google Scholar] [CrossRef]
- Donlao, N.; Ogawa, Y. The influence of processing conditions on catechin, caffeine and chlorophyll contents of green tea (Camelia sinensis) leaves and infusions. LWT 2019, 116, 108567. [Google Scholar] [CrossRef]
- Amani Machiani, M.; Javanmard, A.; Habibi Machiani, R.; Sadeghpour, A. Arbuscular mycorrhizal Fungi and Changes in Primary and Secondary Metabolites. Plants 2022, 11, 2183. [Google Scholar] [CrossRef] [PubMed]
- Kakouridis, A.; Hagen, J.A.; Kan, M.P.; Mambelli, S.; Feldman, L.J.; Herman, D.J.; Weber, P.K.; Pett-Ridge, J.; Firestone, M.K. Routes to roots: Direct evidence of water transport by arbuscular mycorrhizal fungi to host plants. New Phytol. 2022, 236, 210–221. [Google Scholar] [CrossRef]
- Liu, X.; Feng, Z.; Zhao, Z.; Zhu, H.; Yao, Q. Acidic soil inhibits the functionality of arbuscular mycorrhizal fungi by reducing arbuscule formation in tomato roots. Soil Sci. Plant Nutr. 2020, 66, 275–284. [Google Scholar] [CrossRef]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The combined effects of arbuscular mycorrhizal fungi (AMF) and lead (Pb) stress on Pb accumulation, plant growth parameters, photosynthesis, and antioxidant enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef]
- Diaconu, M.; Pavel, L.V.; Hlihor, R.-M.; Rosca, M.; Fertu, D.I.; Lenz, M.; Corvini, P.X.; Gavrilescu, M. Characterization of heavy metal toxicity in some plants and microorganisms—A preliminary approach for environmental bioremediation. New Biotechnol. 2020, 56, 130–139. [Google Scholar] [CrossRef]
- Parada, J.; Rubilar, O.; Fernández-Baldo, M.A.; Bertolino, F.A.; Durán, N.; Seabra, A.; Tortella, G. The nanotechnology among US: Are metal and metal oxides nanoparticles a nano or mega risk for soil microbial communities? Crit. Rev. Biotechnol. 2019, 39, 157–172. [Google Scholar] [CrossRef]
- Shuaib, M.; Azam, N.; Bahadur, S.; Romman, M.; Yu, Q.; Xuexiu, C. Variation and succession of microbial communities under the conditions of persistent heavy metal and their survival mechanism. Microb. Pathog. 2021, 150, 104713. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, J.; Ren, L.; Zhou, Y.; Gao, J.; Luo, L.; Yang, Y.; Peng, Q.; Huang, H.; Chen, A. Diagnosis of soil contamination using microbiological indices: A review on heavy metal pollution. J. Environ. Manag. 2019, 242, 121–130. [Google Scholar] [CrossRef]
- Rajendran, S.; Priya, T.; Khoo, K.S.; Hoang, T.K.; Ng, H.-S.; Munawaroh, H.S.H.; Karaman, C.; Orooji, Y.; Show, P.L. A critical review on various remediation approaches for heavy metal contaminants removal from contaminated soils. Chemosphere 2022, 287, 132369. [Google Scholar] [CrossRef]
- Siddiqua, K.S.; Farooqui, S.A.; Hussain, T.; Mohamed, H.I. Microbial Enzymes and Their Role in Phytoremediation. In Plant Growth-Promoting Microbes for Sustainable Biotic and Abiotic Stress Management; Springer: Berlin/Heidelberg, Germany, 2021; pp. 625–650. [Google Scholar]
- Khan, A. Promises and potential of in situ nano-phytoremediation strategy to mycorrhizo-remediate heavy metal contaminated soils using non-food bioenergy crops (Vetiver zizinoides & Cannabis sativa). Int. J. Phytoremediation 2020, 22, 900–915. [Google Scholar] [PubMed]
- Qin, X.; Zhang, K.; Fan, Y.; Fang, H.; Nie, Y.; Wu, X.-L. The bacterial MtrAB Two-Component System regulates the cell wall homeostasis responding to environmental alkaline stress. Microbiol. Spectr. 2022, 10, e02311–e02322. [Google Scholar] [CrossRef]
- Chen, J.; Guo, J.; Li, Z.; Liang, X.; You, Y.; Li, M.; He, Y.; Zhan, F. Effects of an Arbuscular Mycorrhizal Fungus on the Growth of and Cadmium Uptake in Maize Grown on Polluted Wasteland, Farmland and Slopeland Soils in a Lead-Zinc Mining Area. Toxics 2022, 10, 359. [Google Scholar] [CrossRef]
- Ouledali, S.; Ennajeh, M.; Ferrandino, A.; Khemira, H.; Schubert, A.; Secchi, F. Influence of arbuscular mycorrhizal fungi inoculation on the control of stomata functioning by abscisic acid (ABA) in drought-stressed olive plants. S. Afr. J. Bot. 2019, 121, 152–158. [Google Scholar] [CrossRef]
- Benami, M.; Isack, Y.; Grotsky, D.; Levy, D.; Kofman, Y. The economic potential of arbuscular mycorrhizal fungi in agriculture. In Grand Challenges in Fungal Biotechnology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 239–279. [Google Scholar]
- Nair, A.; Rao, A.S.; Bhanu, L.; More, V.S.; Anantharaju, K.; More, S.S. Arbuscular mycorrhizae, a treasured symbiont to agriculture. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 45–62. [Google Scholar]
- Manale, A.; Hyberg, S. A Guide to Understanding the Fundamental Principles of Environmental Management: It Ain’t Magic: Everything Goes Somewhere; IWA Publishing: London, UK, 2021. [Google Scholar]
- Huang, G.; Ren, H.; Wang, A.; Wan, X.; Wu, Z.; Zhong, X. iTRAQ-based proteomic analysis reveals the molecule mechanism of reducing higher alcohols in Chinese rice wine by nitrogen compensation. Ann. Microbiol. 2021, 71, 4. [Google Scholar] [CrossRef]
- Řezáčová, V.; Czakó, A.; Stehlík, M.; Mayerová, M.; Šimon, T.; Smatanová, M.; Madaras, M. Organic fertilization improves soil aggregation through increases in abundance of eubacteria and products of arbuscular mycorrhizal fungi. Sci. Rep. 2021, 11, 12548. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, M.; Liu, Y.; Zhang, F.; Hodge, A.; Feng, G. Carbon and phosphorus exchange may enable cooperation between an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. New Phytol. 2016, 210, 1022–1032. [Google Scholar] [CrossRef]
- Frey, S.D. Mycorrhizal fungi as mediators of soil organic matter dynamics. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 237–259. [Google Scholar] [CrossRef]
- Afrangan, F.; Kazemeini, S.A.; Alinia, M.; Mastinu, A. Glomus versiforme and Micrococcus yunnanensis reduce the negative effects of salinity stress by regulating the redox state and ion homeostasis in Brassica napus L. crops. In Biologia; Springer: New York, NY, USA, 2023; pp. 1–13. [Google Scholar]
- Jabborova, D.; Annapurna, K.; Paul, S.; Kumar, S.; Saad, H.A.; Desouky, S.; Ibrahim, M.F.; Elkelish, A. Beneficial features of biochar and arbuscular mycorrhiza for improving spinach plant growth, root morphological traits, physiological properties, and soil enzymatic activities. J. Fungi 2021, 7, 571. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; Laouane, R.B.; Anli, M.; Boutasknit, A.; Wahbi, S.; Meddich, A. Use of mycorrhizal fungi in improving tolerance of the date palm (Phoenix dactylifera L.) seedlings to salt stress. Sci. Hortic. 2019, 253, 429–438. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Zhang, F.; Huang, Y. Influence of arbuscular mycorrhiza on organic solutes in maize leaves under salt stress. Mycorrhiza 2011, 21, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Elhindi, K.M.; El-Din, A.S.; Elgorban, A.M. The impact of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects in sweet basil (Ocimum basilicum L.). Saudi J. Biol. Sci. 2017, 24, 170–179. [Google Scholar] [CrossRef]
- Borde, M.; Dudhane, M.; Jite, P.K. AM fungi influences the photosynthetic activity, growth and antioxidant enzymes in Allium sativum L. under salinity condition. Not. Sci. Biol. 2010, 2, 64–71. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Li, Y.; Wu, A.; Huang, J. Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS ONE 2018, 13, e0196408. [Google Scholar] [CrossRef]
- Hameed, A.; Dilfuza, E.; Abd-Allah, E.F.; Hashem, A.; Kumar, A.; Ahmad, P. Salinity stress and arbuscular mycorrhizal symbiosis in plants. In Use of Microbes for the Alleviation of Soil Stresses; Springer: New York, NY, USA, 2014; Volume 1, pp. 139–159. [Google Scholar]
- Talaat, N.B.; Shawky, B.T. Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environ. Exp. Bot. 2014, 98, 20–31. [Google Scholar] [CrossRef]
- Kapoor, R.; Evelin, H.; Mathur, P.; Giri, B. Arbuscular mycorrhiza: Approaches for abiotic stress tolerance in crop plants for sustainable agriculture. In Plant Acclimation to Environmental Stress; Springer: New York, NY, USA, 2013; pp. 359–401. [Google Scholar]
- Aroca, R.; Ruiz-Lozano, J.M.; Zamarreño, Á.M.; Paz, J.A.; García-Mina, J.M.; Pozo, M.J.; López-Ráez, J.A. Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants. J. Plant Physiol. 2013, 170, 47–55. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Chaoxing, H. Does inoculation with Glomus mosseae improve salt tolerance in pepper plants? J. Plant Growth Regul. 2014, 33, 644–653. [Google Scholar] [CrossRef]
- Jabborova, D.; Annapurna, K.; Al-Sadi, A.M.; Alharbi, S.A.; Datta, R.; Zuan, A.T.K. Biochar and Arbuscular mycorrhizal fungi mediated enhanced drought tolerance in Okra (Abelmoschus esculentus) plant growth, root morphological traits and physiological properties. Saudi J. Biol. Sci. 2021, 28, 5490–5499. [Google Scholar] [CrossRef]
- Sharma, M.P.; Grover, M.; Chourasiya, D.; Bharti, A.; Agnihotri, R.; Maheshwari, H.S.; Pareek, A.; Buyer, J.S.; Sharma, S.K.; Schütz, L. Deciphering the role of trehalose in tripartite symbiosis among rhizobia, arbuscular mycorrhizal fungi, and legumes for enhancing abiotic stress tolerance in crop plants. Front. Microbiol. 2020, 11, 509919. [Google Scholar] [CrossRef]
- Hussain, S.; Shaukat, M.; Ashraf, M.; Zhu, C.; Jin, Q.; Zhang, J. Salinity stress in arid and semi-arid climates: Effects and management in field crops. Clim. Change Agric. 2019, 13, 201–226. [Google Scholar]
- Hidri, R.; Mahmoud, O.M.-B.; Farhat, N.; Cordero, I.; Pueyo, J.J.; Debez, A.; Barea, J.-M.; Abdelly, C.; Azcon, R. Arbuscular mycorrhizal fungus and rhizobacteria affect the physiology and performance of Sulla coronaria plants subjected to salt stress by mitigation of ionic imbalance. J. Plant Nutr. Soil Sci. 2019, 182, 451–462. [Google Scholar] [CrossRef]
- Kong, L.; Gong, X.; Zhang, X.; Zhang, W.; Sun, J.; Chen, B. Effects of arbuscular mycorrhizal fungi on photosynthesis, ion balance of tomato plants under saline-alkali soil condition. J. Plant Nutr. 2020, 43, 682–698. [Google Scholar] [CrossRef]
- Lutts, S.; Lefèvre, I. How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann. Bot. 2015, 115, 509–528. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Cui, M.; Wang, C.; Wang, J.; Wang, S.; Sun, Z.; Ren, F.; Wan, S.; Han, S. Elevated CO2, warming, N addition, and increased precipitation affect different aspects of the arbuscular mycorrhizal fungal community. Sci. Total Environ. 2022, 806, 150522. [Google Scholar] [CrossRef]
- Illescas, M.; Rubio, M.B.; Hernández-Ruiz, V.; Morán-Diez, M.E.; Martínez de Alba, A.E.; Nicolás, C.; Monte, E.; Hermosa, R. Effect of inorganic N top dressing and Trichoderma harzianum seed-inoculation on crop yield and the shaping of root microbial communities of wheat plants cultivated under high basal N fertilization. Front. Plant Sci. 2020, 11, 575861. [Google Scholar] [CrossRef] [PubMed]
- Ortaş, I.; Rafique, M.; Iqbal, M.T. Mycorrhizae Resource Allocation in Root Development and Root Morphology. In Plant Microbe Interface; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–26. [Google Scholar]
- Meacham-Hensold, K.; Montes, C.M.; Wu, J.; Guan, K.; Fu, P.; Ainsworth, E.A.; Pederson, T.; Moore, C.E.; Brown, K.L.; Raines, C. High-throughput field phenotyping using hyperspectral reflectance and partial least squares regression (PLSR) reveals genetic modifications to photosynthetic capacity. Remote Sens. Environ. 2019, 231, 111176. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.C.; Graham, J.H.; Smith, F.A. Functioning of mycorrhizal associations along the mutualism–parasitism continuum. New Phytol. 1997, 135, 575–585. [Google Scholar] [CrossRef]
- Kapetsky, J.M.; Aguilar-Manjarrez, J.; Jenness, J. A global assessment of offshore mariculture potential from a spatial perspective. In FAO Fisheries and Aquaculture Technical Paper; FAO: United Nations: Roma, Italy, 2013; p. I. [Google Scholar]
- Smith, S.E.; Smith, F.A. Fresh perspectives on the roles of arbuscular mycorrhizal fungi in plant nutrition and growth. Mycologia 2012, 104, 1–13. [Google Scholar] [CrossRef]
- Huey, C.J.; Gopinath, S.C.; Uda, M.; Zulhaimi, H.I.; Jaafar, M.N.; Kasim, F.H.; Yaakub, A.R.W. Mycorrhiza: A natural resource assists plant growth under varied soil conditions. 3 Biotech 2020, 10, 204. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Pichtel, J. Mycorrhizae: An overview. In Mycorrhizae: Sustainable Agriculture and Forestry; Springer: Berlin/Heidelberg, Germany, 2008; pp. 1–35. [Google Scholar]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Saboor, A.; Ali, M.A.; Hussain, S.; El Enshasy, H.A.; Hussain, S.; Ahmed, N.; Gafur, A.; Sayyed, R.; Fahad, S.; Danish, S. Zinc nutrition and arbuscular mycorrhizal symbiosis effects on maize (Zea mays L.) growth and productivity. Saudi J. Biol. Sci. 2021, 28, 6339–6351. [Google Scholar] [CrossRef] [PubMed]
- Khan, Y.; Shah, S.; Tian, H. The roles of arbuscular mycorrhizal fungi in influencing plant nutrients, photosynthesis, and metabolites of cereal crops—A review. Agronomy 2022, 12, 2191. [Google Scholar] [CrossRef]
- Pepe, A.; Di Baccio, D.; Magnani, E.; Giovannetti, M.; Sbrana, C. Zinc and Iron Biofortification and Accumulation of Health-Promoting Compounds in Mycorrhizal Cichorium intybus L. J. Soil Sci. Plant Nutr. 2022, 22, 4703–4716. [Google Scholar] [CrossRef]
- Roleda, M.Y.; Hurd, C.L. Seaweed nutrient physiology: Application of concepts to aquaculture and bioremediation. Phycologia 2019, 58, 552–562. [Google Scholar] [CrossRef]
- Hallama, M.; Pekrun, C.; Lambers, H.; Kandeler, E. Hidden miners–the roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 2019, 434, 7–45. [Google Scholar] [CrossRef]
- Achatz, M.; Morris, E.K.; Müller, F.; Hilker, M.; Rillig, M.C. Soil hypha-mediated movement of allelochemicals: Arbuscular mycorrhizae extend the bioactive zone of juglone. Funct. Ecol. 2014, 28, 1020–1029. [Google Scholar] [CrossRef]
- Ll, H.; Li, X.; Dou, Z.; Zhang, J.; Wang, C. Earthworm (Aporrectodea trapezoides)–mycorrhiza (Glomus intraradices) interaction and nitrogen and phosphorus uptake by maize. Biol. Fertil. Soils 2012, 48, 75–85. [Google Scholar] [CrossRef]
- Soumare, A.; Diedhiou, A.G.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kouisni, L. Exploiting biological nitrogen fixation: A route towards a sustainable agriculture. Plants 2020, 9, 1011. [Google Scholar] [CrossRef]
- Lone, R.; Shuab, R.; Khan, S.; Ahmad, J.; Koul, K. Arbuscular mycorrhizal fungi for sustainable agriculture. In Probiotics and Plant Health; Springer: Berlin/Heidelberg, Germany, 2017; pp. 553–577. [Google Scholar]
- Vannini, C.; Domingo, G.; Fiorilli, V.; Seco, D.G.; Novero, M.; Marsoni, M.; Wisniewski-Dye, F.; Bracale, M.; Moulin, L.; Bonfante, P. Proteomic analysis reveals how pairing of a Mycorrhizal fungus with plant growth-promoting bacteria modulates growth and defense in wheat. Plant Cell Environ. 2021, 44, 1946–1960. [Google Scholar] [CrossRef]
- Jach-Smith, L.C.; Jackson, R.D. Inorganic N addition replaces N supplied to switchgrass (Panicum virgatum) by arbuscular mycorrhizal fungi. Ecol. Appl. 2020, 30, e02047. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; Ullah, H.; Himanshu, S.K.; Tisarum, R.; Cha-Um, S.; Datta, A. Arbuscular mycorrhizal fungi inoculation and phosphorus application improve growth, physiological traits, and grain yield of rice under alternate wetting and drying irrigation. J. Plant Physiol. 2022, 278, 153829. [Google Scholar] [CrossRef] [PubMed]
- Pompelli, M.F.; Espitia-Romero, C.A.; de Diós Jaraba-Navas, J.; Rodriguez-Paez, L.A.; Jarma-Orozco, A. Stevia rebaudiana under a CO2 Enrichment Atmosphere: Can CO2 Enrichment Overcome Stomatic, Mesophilic and Biochemical Barriers That Limit Photosynthesis? Sustainability 2022, 14, 14269. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, L.; Shangguan, Z. Appropriate N addition improves soil aggregate stability through AMF and glomalin-related soil proteins in a semiarid agroecosystem. Land Degrad. Dev. 2022, 34, 710–722. [Google Scholar] [CrossRef]
- Bonfante, P.; Desirò, A. Arbuscular mycorrhizas: The lives of beneficial fungi and their plant hosts. In Principles of Plant-Microbe Interactions; Springer: Berlin/Heidelberg, Germany, 2015; pp. 235–245. [Google Scholar]
- Atakan, A.; Özkaya, H.Ö. Arbuscular Mycorrhizal Fungi and Glomalin. Turk. J. Agric.-Food Sci. Technol. 2021, 9, 2371–2375. [Google Scholar] [CrossRef]
- Li, Z.; Wu, S.; Liu, Y.; Yi, Q.; You, F.; Ma, Y.; Thomsen, L.; Chan, T.-S.; Lu, Y.-R.; Hall, M. Arbuscular mycorrhizal symbiosis enhances water stable aggregate formation and organic matter stabilization in Fe ore tailings. Geoderma 2022, 406, 115528. [Google Scholar] [CrossRef]
- Morris Jr, S.M. Arginases and arginine deficiency syndromes. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 64. [Google Scholar] [CrossRef]
- Rubin, J.A.; Görres, J.H. Potential for mycorrhizae-assisted phytoremediation of phosphorus for improved water quality. Int. J. Environ. Res. Public Health 2021, 18, 7. [Google Scholar] [CrossRef]
- Smit, A.L.; Bindraban, P.S.; Schröder, J.; Conijn, J.; Van der Meer, H. Phosphorus in agriculture: Global Resoources, Trends and Developments: Report to the Steering Committee Technology Assessment of the Ministery of Agriculture, Nature and Food Quality, The Netherlands, and in Collaboration with the Nutrient Flow Task Group (NFTG), Supported by DPRN (Development Policy Review Network); Plant Research International: Wageningen, The Netherlands, 2009; pp. 1566–7790. [Google Scholar]
- Etesami, H. Enhanced phosphorus fertilizer use efficiency with microorganisms. In Nutrient Dynamics for Sustainable Crop Production; Springer: Berlin/Heidelberg, Germany, 2020; pp. 215–245. [Google Scholar]
- Bononi, L.; Chiaramonte, J.B.; Pansa, C.C.; Moitinho, M.A.; Melo, I.S. Phosphorus-solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. Sci. Rep. 2020, 10, 2858. [Google Scholar] [CrossRef]
- El Maaloum, S.; Elabed, A.; Alaoui-Talibi, Z.E.; Meddich, A.; Filali-Maltouf, A.; Douira, A.; Ibnsouda-Koraichi, S.; Amir, S.; El Modafar, C. Effect of arbuscular mycorrhizal fungi and phosphate-solubilizing bacteria consortia associated with phospho-compost on phosphorus solubilization and growth of tomato seedlings (Solanum lycopersicum L.). Commun. Soil Sci. Plant Anal. 2020, 51, 622–634. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
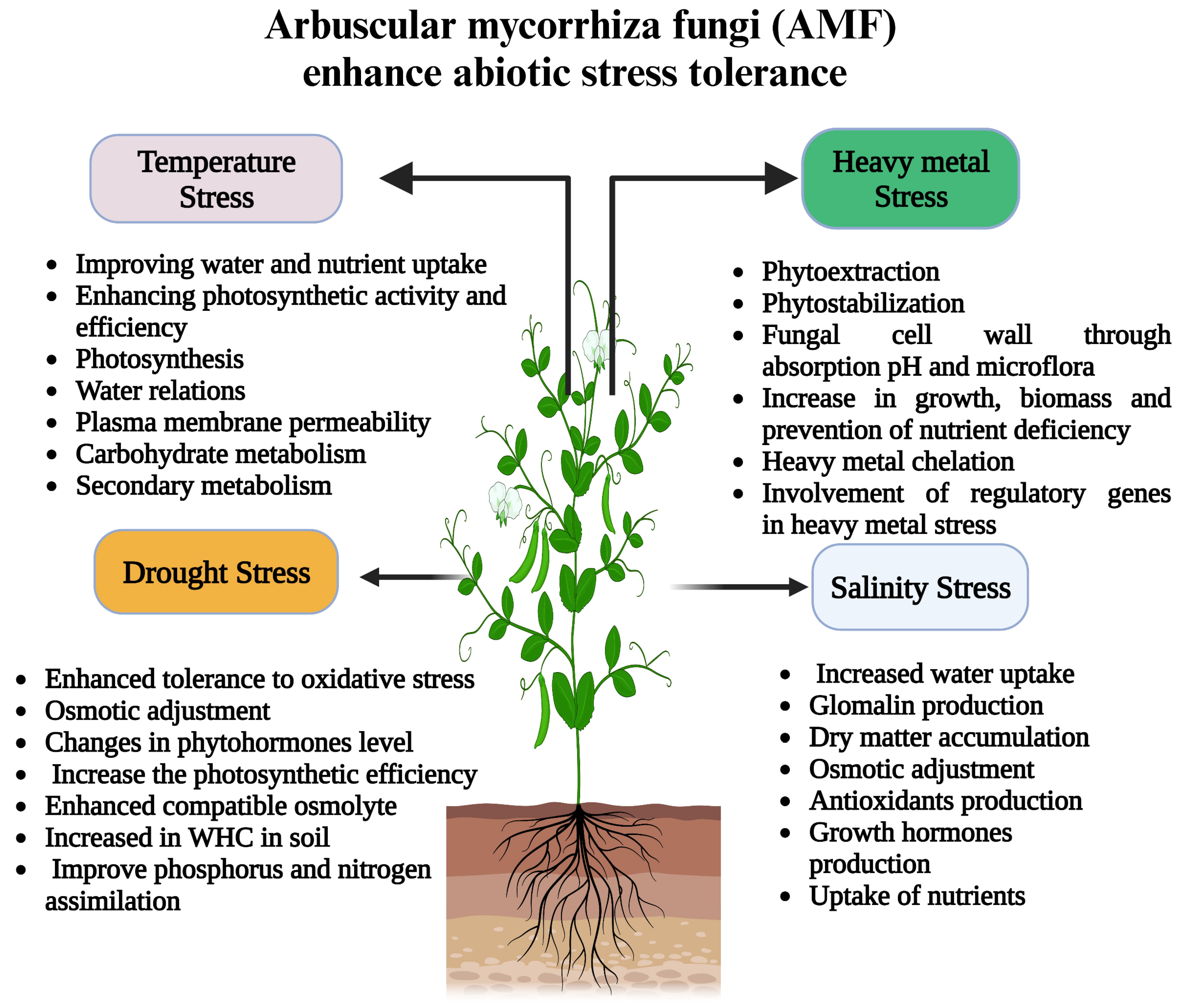{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMF Symbionts | Crop Tested | Agroecosystem Service | Conditions | References |
|---|---|---|---|---|
| Rhizophagus irregularis and Funneliformis mosseae | Chickpea (Cicer arietinum L.) | Enhanced plant mass and superior grain characteristics. | Agriculture land | [24] |
| A mixture of Glomus clarum, Gigaspora margarita, and Acaulospora sp. | Coffee (Coffea arabica L.) | Protection against Cu and Zn toxicity. | Greenhouse | [25] |
| Indigenous AMF consortium: G. etunicatum, G. fasciculatum, Glomus mosseae, G. intraradices, and Scutellospora sp. | Green pepper (Capsicum annuum L.), parsley, and tomato (Solanum lycopersicum L.) | Elevated crop yields and superior plant vitality. | Greenhouse and field | [26] |
| G. mosseae Consortium: G. intraradices, G. aggregatum/intraradices, Entrophospora infrequens Acaulospora trappei, and Glomus sp. | Lettuce (Lactuca sativa L.) and Onion (Allium cepa L.) | Tolerance to saline environments. | Greenhouse | [27] |
| G. fasciculatum, G. etunicatum, G. clarum and G. versiforme | Long pepper (Piper longum L.) | Superior growth attributes. | Garden centre and field | [28] |
| G. aggregatum, Consortium: R. intraradices, G. viscosum, G. etunicatum, and G. claroideum | Maize (Zea Mays L.) | Optimal crop growth, output, and grain attributes. | Arena | [29] |
| Native AMF | Maize (Z. mays L.) | Augmented absorption of K, Ca, and Mg. | Arena | [30] |
| G. mosseae and G. intraradices | Lucerne (Medicago sativa L.) | It boosted GRSP and better soil cohesiveness. | Greenhouse | [31] |
| G. etunicatum, G. mosseae, G. fasciculatum, and Gigaspora margarita | Pepper (C. annuum) | Growth promotion and mitigation of Phytophthora infections. | Pot, greenhouse, and field | [32] |
| Glomus clarum | Pepper (C. annuum) | Augmented fruit production in saline-rich conditions. | Glasshouse | [33] |
| On-farm produced G. intraradices | Potato (Solanum tuberosum L.) | Bigger harvests. | Field | [34] |
| G. etunicatum | Soybean (Glycine max (L.) Merr.) | It helps to elevate growth performance under saline circumstances. | Greenhouse | [35] |
| G. mosseae | Soybean (G. max) and Lentil (Lens culinaris Medic) | Enhanced absorption of Zn. | Greenhouse | [36] |
| G. intraradices, G. mosseae, and G. etunicatum | Strawberry (Fragaria × ananassa Duch.) | Top-tier quality. | Field | [37,38] |
| G. intraradices | Strawberry (F. × ananassa) | Boosted fruit attributes. | Glasshouse | [39] |
| G. mosseae and G. hoi | Sunflower (Helianthus annuus L.) | It helps to promote advanced agricultural yields. | Glasshouse | [32] |
| G. mosseae, G. intraradices, and G. coronatum | Sunflower (H. annuus) | Tackling wildflowers and optimizing phosphorus absorption. | Greenhouse | [39] |
| Native AMF inoculum (consortium) | Tomato (Solanum lycopersicum L.) | Modifying plant responses to zinc inclusions. | Environment chamber | [40] |
| G. intraradices | Tomato (S. lycopersicum) | Addressing the root disease caused by Alternaria solani. | Climate chamber | [41] |
| G. intraradices | Tomato (S. lycopersicum) | It enhanced cultivation, processing, and fruit quality in waterlogged conditions. | Field | [42] |
| G. mosseae | Tomato (S. lycopersicum) | Addressing issues from root-knot nematode Meloidogyne incognita. | Greenhouse | [43] |
| G. intraradices | Tomato (S. lycopersicum) | Superior resilience to saline conditions. | Glasshouse | [44] |
| G. intraradices | Tomato (S. lycopersicum) and Onion (A. cepa) | Advanced harvest results. | Field | [29] |
| G. intraradices | Tomato (S. lycopersicum) | Dealing with the deceptive root-knot nematode Nacobbus aberrans. | Greenhouse | [45] |
| G. intraradices | Tomato (S. lycopersicum) | Augmented growth, flowering, and yield. | Field | [46] |
| G. mosseae | Tomato (S. lycopersicum) | Boosted growth cycle, flowering stage, and fruit formation. | Growth chamber | [47] |
| G. mosseae and G. intraradices isolates and native AMF inoculum (consortium) | Clover (Trifolium alexandrinum L.) and maize (Z. mays L.) | Superior agricultural output. | Field | [26] |
| G. etunicatum and G. mosseae | Wheat (Triticum aestivum L.) and Brinjal (Solanum melongena) | It helps to enhance plant growth, yield, and nutrient absorption. | Field | [48] |
| G. mosseae, G. hoi, G. etunicatum, A. scrobiculata, and A. spinosa | Yam (Dioscorea rotundata) | It helps to boost root expansion. | Greenhouse | [26] |
| Consortium of G. mosseae, G. deserticola, and A. laevis | Yam (D. rotundata and D. alata) | Elevated nutritional content. | Glasshouse | [49] |
| Host Plant | AMF Strains | Stress | Observed Responses | References |
|---|---|---|---|---|
| Solanum lycopersicum | Rhizophagus irregularis | Salinity | Growth hormone promotes plant health, increasing root and shoot weight and improving leaf structure. | [106] |
| Leymus chinensis | Glomus mosseae | Salinity | Positive outcomes include elevated phosphorus and nitrogen levels, enhanced seedling weight, and increased plant water content. | [107] |
| Cucumis sativus L. | Claroideoglomus etunicatum, Rhizophagus intraradices, Funneliformis mosseae | Salinity | Enhanced growth, higher antioxidant enzyme activity, elevated proline and phenolic content, and improved uptake of vital mineral elements have been observed. Furthermore, the absorption of sodium ions was reduced. | [108] |
| Medicago sativa | Glomus mosseae | Salinity | Among the crucial nutrients for plants, phosphorus (P), nitrogen (N), and potassium (K) play pivotal roles. | [109] |
| Glycine max L. Merrill | Claroideoglomus etunicatum, Rhizophagus intraradices, Funneliformis mosseae | Salinity | Benefits encompass improved plant and root systems, heightened nutrient uptake, minimized lipid peroxidation, and reduced membrane damage. | [27] |
| Prunus dulcis x Prunus persica hybrid | Rhizophagus intraradices, Funneliformis mosseae | Salinity | Plant growth sees improvements through elevated antioxidant enzymes, increased photosynthetic compounds, soluble sugars, and proline content. | [110] |
| Pisum sativum L. | Rhizoglomus intraradices, Funneliformis mosseae, Rhizoglomus fasciculatum, Gigaspora spp. | Salinity | Enhanced biomass, chlorophyll content, nutrient absorption, and accumulation of compatible osmolytes contribute to overall plant well-being. | [111] |
| Euonymus maackii Rupr | Rhizophagus intraradices | Salinity | Notable advantages include enhanced photosynthesis, increased nutrient assimilation, and improved antioxidant enzyme activity. | [112] |
| Citrullus lanatus L. | Glomus mosseae, Gigaspora gigantean | Salinity | Plants display heightened foliage coverage, larger fruit dimensions, improved root establishment, elevated nutrient concentrations, and increased antioxidant enzyme activity. | [113] |
| Eucalyptus camaldulensis | Glomus spp., Gigaspora albida, Gigaspora decipiens | Salinity | An increase in photosynthetic pigments, reduced leaf proline, and favorable effects on physiological and biochemical parameters were noted. | [114] |
| Zea mays L. | Rhizophagus intraradices, Funneliformis mosseae, Funneliformis geosporum | High temperature | Positive plant attributes, augmented photosynthetic transpiration rate, and improved pigments enhance overall growth. | [115] |
| Triticum aestivum | Rhizophagus irregularis, Funneliformis mosseae, Funneliformis geosporum, Claroideoglomus claroideum | High temperature | Improved nutrient uptake and increased grain numbers are among the observed effects. | [116] |
| Zea mays L. | Glomus tortuosum | Temperature stress | Elevated levels of shoot nitrogen (N), phosphorus (P), potassium (K), and copper (Cu), along with increased nitrate reductase activity, were recorded. | [117] |
| Zea mays L. | Glomus tortuosum | Cold stress | A notable increase in amino acid concentrations was observed. | [118,119] |
| Elymus nutans Griseb. | Funneliformis mosseae | Cold stress | Enhanced antioxidant enzymes, photosynthetic pigments, and overall plant growth were evident. | [120] |
| Hordeum vulgare L. | Glomus versiforme, Rhizophagus irregularis | Cold stress | The presence of elevated antioxidants, osmoprotectants, and increased potassium uptake positively impacted plant growth and metabolism of phenolics. | [121] |
| Cucumis sativus L. | Rhizophagus irregularis | Cold stress | Photosynthetic efficiency and carbon sink both showed improvement. | [122] |
| Grapevine (Vitis vinifera L.) | Rhizoglomus irregulare, Funneliformis mosseae | High-temperature stress | The growth rate increased alongside enhanced substrate carbon conversion efficiency and stomatal conductance. | [123] |
| Zea maize L. | Funneliformis | High temperature | Regulation of photosystem (PS) II heterogeneity was observed. | [115] |
| Solanum lycopersicum, Capiscum annuum, Cucumis sativus | Rhizophagus irregularis | High-temperature stress | Increased vigour, productivity, and fruit quality were prominent outcomes. | [124] |
| Saccharum arundinaceum | Glomus spp. | Drought | Enhanced levels of antioxidant enzymes, phenolics, chlorophyll, and plant biomass were found in leaves. | [125] |
| Triticum aestivum | Glomus mosseae | Drought | Improved chlorophyll levels, higher content of antioxidant enzymes ascorbic acid, and increased nitrogen (N), phosphorus (P), and potassium (K) content were noted. | [126] |
| Ipomoea batatas | Glomus spp. | Drought | Osmoprotectants played a role in adjusting osmotic potential. | [127] |
| Lycopersicon esculatum, Capsicum annuum | Rhizophagus irregularis, Rhizophagus fasciculatus | Drought | Increases in biomass, root and shoot length, and photosynthetic pigments were observed, while proline concentration decreased. | [128] |
| Solanum lycopersicum | Funneliformis mosseae, Rhizophagus irregularis | Drought | Enhanced plant height, stomatal conductance, water use efficiency, biomass, and reduced levels of reactive oxygen species (ROS) and abscisic acid (ABA) were recorded. | [129] |
| Triticum aestivum L. | Glomus mosseae, Glomus fasciculatum, Gigaspora decipiens | Drought | Positive effects on plant growth parameters and photosynthetic pigments were evident. | [130] |
| Digitaria eriantha | Rhizophagus irregularis | Drought | Increased shoot dry weight, stomatal conductance, lipid peroxidation, and ROS levels were observed in both shoot and root. | [131] |
| Triticum durum | Rhizophagus intraradices | Drought | Enhanced grain biomass, micronutrient content, and gliadins in grains were notable outcomes. | [132] |
| Poncirus trifoliate | Funneliformis mosseae, Paraglomus occultum | Drought | Growth attributes saw improvement through increased root weight and length, higher fructose and glucose levels, lower sucrose levels, and proline accumulation. | [133] |
| Cupressus arizonica | Rhizophagus irregularis, Funneliformis mosseae | Drought | Growth was enhanced, and levels of hydrogen peroxide and malondialdehyde were reduced. | [134] |
| Ephedra foliata Boiss | Glomus etunicatum, Rhizophagus intraradices, Funneliformis mosseae | Drought | Antioxidant enzyme activity, proline, glucose, and total soluble protein levels increased, improving nitrogen metabolism. | [135] |
| Zea mays L. | Rhizophagus irregularis | Drought | AM plant roots demonstrated diamine oxidase activity, converting putrescine into aminobutyric acid (GABA). | [136] |
| Ceratonia silique | Glomus, Gigaspora, Acaulospora, Entrophospora | Drought | Positive impacts included increased plant growth, nutrient levels, stomatal conductance, photosystem II (PSII) efficiency, and water content. | [137] |
| Catalpa bungee C.A.Mey | Rhizophagus intraradices | Drought | Root morphology, water content, biomass, photosynthetic pigments, and various plant hormones (except ABA) all showed improvement. | [138] |
| Cinnamomum migao | Glomus lamellosum, Glomus etunicatum | Drought | Enhanced antioxidant enzyme activity and osmoprotectants led to reduced malondialdehyde levels. | [139] |
| Sesamum indicum L. | Funneliformis mosseae, Rhizophagus intraradices | Drought | Improved oil and seed yield, total soluble protein, leaf phosphorus content, and heightened photosynthetic pigments were observed. | [140] |
| Lonicera japonica Thunb. | Rhizophagus intraradices, Glomus versiforme | Cd | Lower cadmium (Cd) levels in shoots and roots were noted, with more significant accumulation in roots. This indicated enhanced Cd tolerance. | [141] |
| Solanum lycopersicum L. | Funneliformis mosseae, Rhizophagus intraradices, Claroideoglomus etunicatum | Cd | Reduced malondialdehyde and ROS levels provided improved protection against Cd stress. | [142] |
| Cajanus cajan L. | Rhizophagus irregularis | Metals—cadmium and zinc | Increases in root biomass, macro- and micronutrients, and proline formation were observed. | [108] |
| Zea mays L. | Glomus intraradices | Heavy metal: cadmium | Combined effects were seen regarding soil alkalinization, Cd immobilization, and reduced Cd phytoavailability. | [143] |
| Trigonella foenumgraecum | Glomus monosporum, Glomus clarum, Gigaspora nigra | Metals—cadmium | Enhanced antioxidant enzyme activities and malondialdehyde content contributed to phytostabilization. | [144] |
| Trigonella foenumgraecum | Glomus monosporum, Glomus clarum, Gigaspora nigra | Cd | HX3 and HN89 plants showed no significant impacts on Cd accumulation or translocation. | [145] |
| Glycine max | Rhizophagus irregularis | Cd | Increased glomalin production and metal uptake were evident in plants. | [145] |
| Helianthus annuus | Glomus mosseae, Glomus intraradices | Cr, Mn, Ni, Cu, Zn, Al, Pb, Co, Mo, Fe, and Si | Root colonization was enhanced, increasing root and shoot dry weight and higher phosphorus content. | [146] |
| Zea mays L. | Rhizophagus fasciculatus, Funneliformis mosseae, Rhizophagus intraradices, Glomus aggregatum | Cd, Cr, Ni, Pb | Elevated levels of total chlorophyll content and net photosynthesis rate were observed. | [147] |
| Phragmites australis | Funneliformis mosseae | TiO2NPs | The plant’s biomass, growth, and physiological properties all experienced increases. | [148] |
| Cynodon dactylon | Funneliformis mosseae, Diversisporas purcum | Pb, Zn, Cd | The vulnerability of standing milkvetch to powdery mildew was enhanced, accompanied by improved shoot and root growth. | [149] |
| Medicago sativa | Glomus aggregatum, Glomus intraradices, Glomus elunicatum, Glomus versiforme | Cd | Dry weight, growth, yield, and production of antimicrobial substances all increased in plants. | [150] |
| Medicago truncatula | Rhizophagus irregularis | Pb | Reduced crop plant infections resulted in improved growth and yield. | [151] |
| Phragmites australis | Rhizophagus irregularis | Cu | Plant growth was heightened, along with an increase in functional leaf quantity. | [152] |
| Sorghum vulgare | Acaulospora fragilissima, Acaulospora saccata, Claroideoglomus etunicatum, Pervetustus simplex, Rhizophagus neocaledonicus, Scutellospora ovalis, Rhizophagus neocaledonicus | Ultramafic soils (Fe, Mn, Ni, Cr, and Co) | Growth hormone promotes plant health, increasing root and shoot weight and improving leaf structure. | [153] |
| Solanum lycopersicum L. | Funneliformis mosseae | Cladosporium fulvum | Positive outcomes include elevated phosphorus and nitrogen levels, enhanced seedling weight, and increased plant water content. | [154] |
| Saccharum offcinarum L. | Gigaspora margarita, G. etunicatum, Scutellospora fulgida | - | Enhanced growth, higher antioxidant enzyme activity, elevated proline and phenolic content, and improved uptake of vital mineral elements have been observed. Furthermore, the absorption of sodium ions was reduced. | [155] |
| Astragalus adsurgens var. Shanxi Yulin | Claroideoglomus etunicatum, Glomus versiforme, Funneliformis mosseae | Erysiphe pisi DC 1805 | Among the crucial nutrients for plants, phosphorus (P), nitrogen (N), and potassium (K) play pivotal roles. | [156] |
| Lycopersicon esculentum, Capsicum annuum | Rhizophagus irregularis, Rhizophagus fasciculatus | Fusarium oxysporum f. sp. lycopersici | Benefits encompass improved plant and root systems, heightened nutrient uptake, minimized lipid peroxidation, and reduced membrane damage. | [157] |
| Capsicum annum | Glomus spp. | Pythium aphanidermatum | Plant growth sees improvements through elevated antioxidant enzymes, increased photosynthetic compounds, soluble sugars, and proline content. | [158] |
| Glycine max (L.) Merr | Rhizophagus irregularis | Macrophomina phaseolina | Enhanced biomass, chlorophyll content, nutrient absorption, and accumulation of compatible osmolytes contribute to overall plant well-being. | [159] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wahab, A.; Muhammad, M.; Munir, A.; Abdi, G.; Zaman, W.; Ayaz, A.; Khizar, C.; Reddy, S.P.P. Role of Arbuscular Mycorrhizal Fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses. Plants 2023, 12, 3102. https://doi.org/10.3390/plants12173102
Wahab A, Muhammad M, Munir A, Abdi G, Zaman W, Ayaz A, Khizar C, Reddy SPP. Role of Arbuscular Mycorrhizal Fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses. Plants. 2023; 12(17):3102. https://doi.org/10.3390/plants12173102
Chicago/Turabian StyleWahab, Abdul, Murad Muhammad, Asma Munir, Gholamreza Abdi, Wajid Zaman, Asma Ayaz, Chandni Khizar, and Sneha Priya Pappula Reddy. 2023. "Role of Arbuscular Mycorrhizal Fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses" Plants 12, no. 17: 3102. https://doi.org/10.3390/plants12173102