

Unraveling the Mechanisms of Virus-Induced Symptom Development in Plants

Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
| Virus Name | Symptoms | Crops/Plants | References |
|---|---|---|---|
| Banana bunchy top virus | Stunted growth, twisting of leaves | Banana | [38,39] |
| Barley yellow dwarf virus | Yellowing, stunting, reduced grain yield | Wheat | [40,41] |
| Cassava mosaic virus | Mosaic, distortion, reduced root yield | Cassava | [42] |
| Citrus tristeza virus | Yellowing, stem pitting, leaf curling | Citrus spp. | [43] |
| Cucumber mosaic virus | Mosaic, distortion, yellowing | Cucumber | [44] |
| Faba bean necrotic yellows virus | Yellowing, necrosis, stunting | Faba bean | [45] |
| Grapevine leafroll-associated viruses | Yellowing, leaf curling, reduced fruit quality | Grapevine | [46,47,48] |
| Maize dwarf mosaic virus | Mosaic, stunting, yellow streaks | Maize | [49] |
| Maize streak virus | Yellow streaks on leaves, stunted growth | Maize | [50,51] |
| Papaya ringspot virus | Ring spots, leaf distortion, fruit deformities | Papaya | [52,53] |
| Plum pox virus | Yellow rings, fruit deformation | Stone fruits | [54,55] |
| Potato virus Y | Leaf mosaic, yellowing, necrosis | Potato | [56,57] |
| Rice tungro spherical virus | Stunting, yellowing, reduced grain yield | Rice | [58] |
| Rice yellow mottle virus | Yellowing, stunting, yellow mottling on leaves | Rice | [59,60] |
| Southern rice black-streaked dwarf virus | Stunted growth, black streaks on leaves | Rice | [61] |
| Sugarcane mosaic virus | Mosaic, yellow streaks, stunting | Maize | [62] |
| Sweet potato chlorotic stunt virus | Yellowing, stunting, leaf deformation | Sweet potato | [63,64] |
| Tomato spotted wilt virus | Wilted leaves, ring spots, bronzing | Tomato | [65,66] |
| Tomato yellow leaf curl virus | Yellowing, leaf curling, reduced fruit set | Tomato | [67,68] |
| Zucchini yellow mosaic virus | Mosaic, yellowing, distortion | Cucurbits | [69,70] |
2. Alterations in Plant Cellular Processes
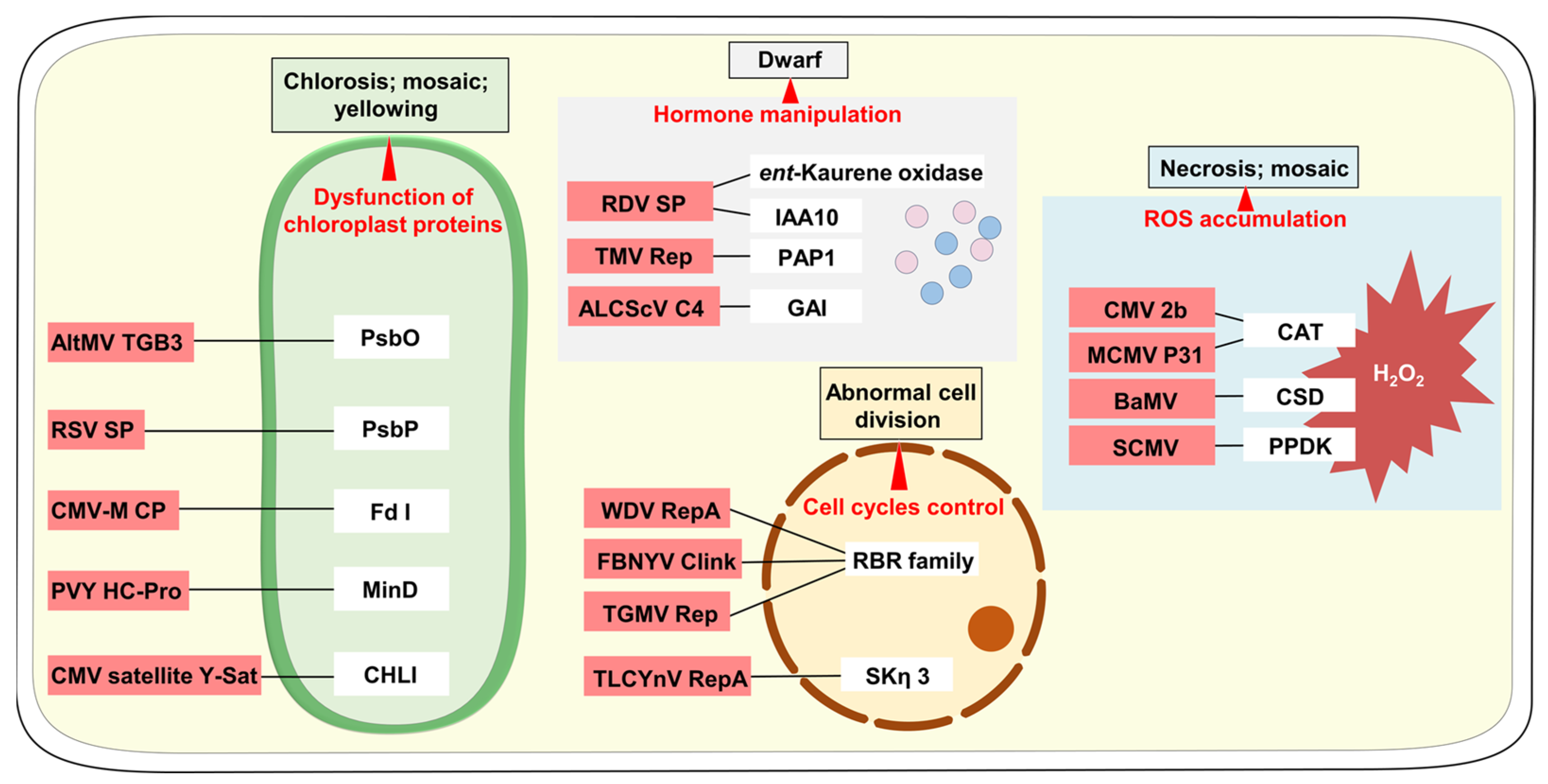
2.1. Dysfunction of Chloroplast Proteins
2.2. Hormone Manipulation
2.3. ROS Accumulation
2.4. Cell Cycle Control
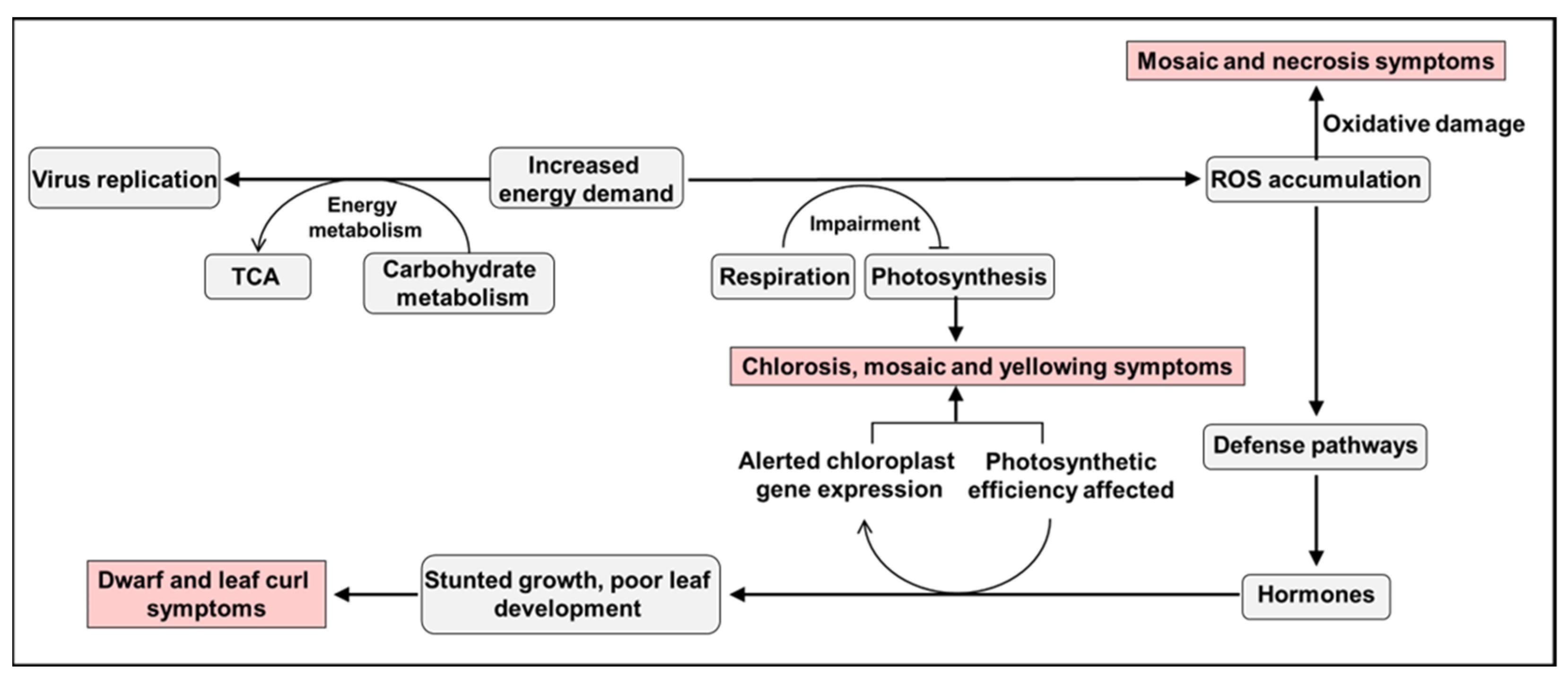
3. Host Metabolic and Genetic Responses to Virus-Induced Symptoms
3.1. Metabolic Disorders
3.2. Gene Expression Changes
4. The Relationship between Environmental Factors and Virus-Induced Symptoms
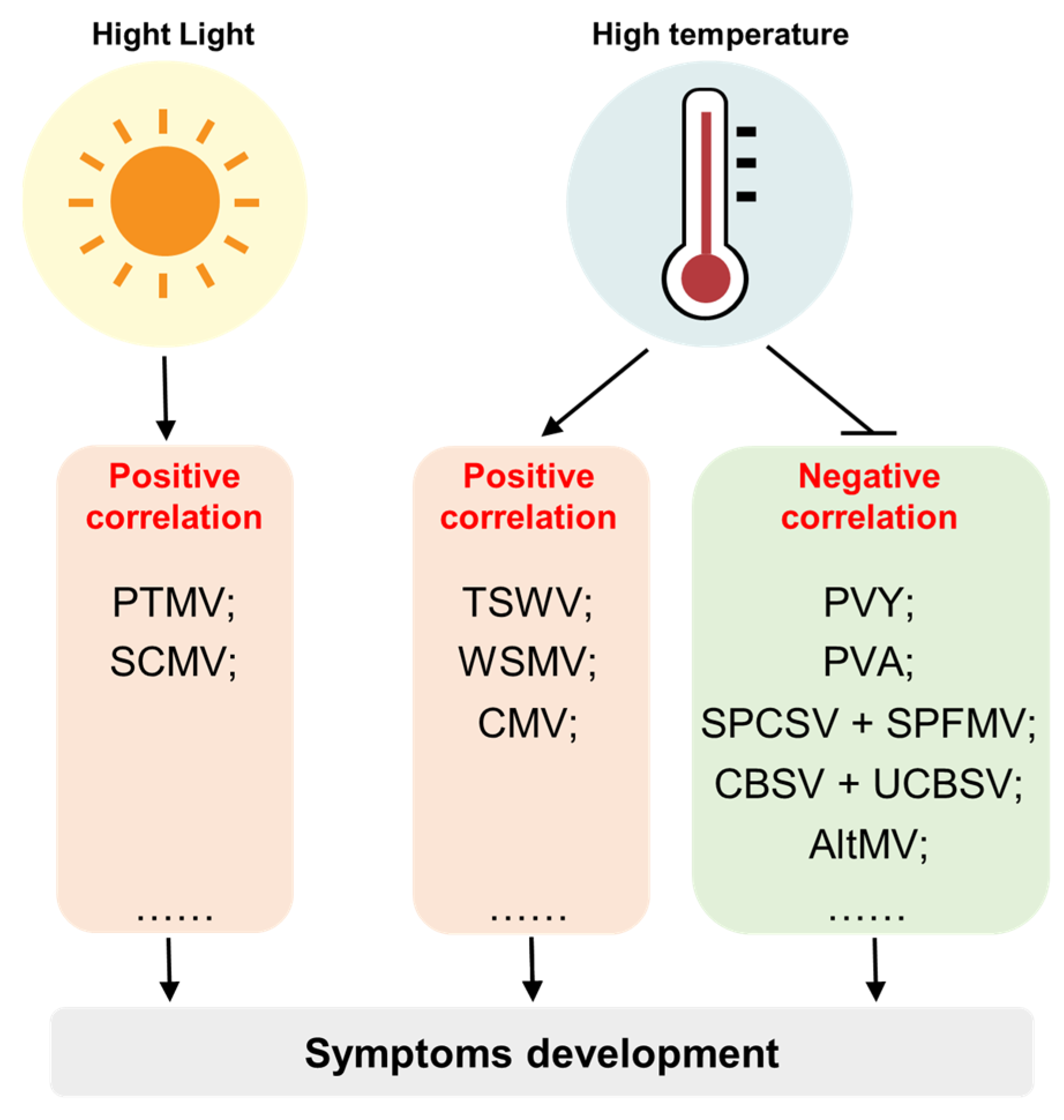
4.1. The Role of Light Conditions in Virus-Induced Symptom Development
4.2. The Role of Temperature in Virus-Induced Symptoms Development
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Roossinck, M.J. Lifestyles of plant viruses. Philos. Trans. R. Soc. B 2010, 365, 1899–1905. [Google Scholar] [CrossRef] [Green Version]
- Hull, R. Symptoms and Host Range. In Plant Virology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 145–198. [Google Scholar]
- Nicaise, V. Crop immunity against viruses: Outcomes and future challenges. Front. Plant Sci. 2014, 5, 660. [Google Scholar] [CrossRef] [PubMed]
- Llave, C. Dynamic cross-talk between host primary metabolism and viruses during infections in plants. Curr. Opin. Virol. 2016, 19, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Calvino, L.; Osorio, S.; Hernandez, M.L.; Hamada, I.B.; del Toro, F.J.; Donaire, L.; Yu, A.; Bustos, R.; Fernie, A.R.; Martinez-Rivas, J.M.; et al. Virus-induced alterations in primary metabolism modulate susceptibility to Tobacco rattle virus in Arabidopsis. Plant Physiol. 2014, 166, 1821–1838. [Google Scholar] [CrossRef] [PubMed]
- De Storme, N.; Geelen, D. Callose homeostasis at plasmodesmata: Molecular regulators and developmental relevance. Front. Plant Sci. 2014, 5, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stare, T.; Ramsak, Z.; Blejec, A.; Stare, K.; Turnsek, N.; Weckwerth, W.; Wienkoop, S.; Vodnik, D.; Gruden, K. Bimodal dynamics of primary metabolism-related responses in tolerant potato-Potato virus Y interaction. BMC Genom. 2015, 16, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimura, H.; Pantaleo, V.; Ishihara, T.; Myojo, N.; Inaba, J.; Sueda, K.; Burgyan, J.; Masuta, C. A viral satellite RNA induces yellow symptoms on tobacco by targeting a gene involved in chlorophyll biosynthesis using the RNA silencing machinery. PLoS Pathog. 2011, 7, e1002021. [Google Scholar] [CrossRef] [Green Version]
- Hamel, L.P.; Sekine, K.T.; Wallon, T.; Sugiwaka, Y.; Kobayashi, K.; Moffett, P. The chloroplastic protein THF1 interacts with the coiled-coil domain of the disease resistance protein N′ and regulates light-dependent cell death. Plant Physiol. 2016, 171, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Pallas, V.; Garcia, J.A. How do plant viruses induce disease? Interactions and interference with host components. J. Gen. Virol. 2011, 92, 2691–2705. [Google Scholar] [CrossRef]
- Jiao, Z.; Tian, Y.; Cao, Y.; Wang, J.; Zhan, B.; Zhao, Z.; Sun, B.; Guo, C.; Ma, W.; Liao, Z.; et al. A novel pathogenicity determinant hijacks maize catalase 1 to enhance viral multiplication and infection. New Phytol. 2021, 230, 1126–1141. [Google Scholar] [CrossRef]
- Jiang, T.; Du, K.; Xie, J.; Sun, G.; Wang, P.; Chen, X.; Cao, Z.; Wang, B.; Chao, Q.; Li, X.; et al. Activated malate circulation contributes to the manifestation of light-dependent mosaic symptoms. Cell Rep. 2023, 42, 112333. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, Q.; Wu, Y.; Huang, F.; Ismayil, A.; Zhang, D.; Li, H.; Gu, H.; Ludman, M.; Fatyol, K.; et al. A calmodulin-binding transcription factor links calcium signaling to antiviral RNAi defense in plants. Cell Host Microbe 2021, 29, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, Z.; Yue, N.; Jin, X.; Zhang, X.; Li, Z.; Zhang, Y.; Wang, X.B.; Han, C.; Yu, J.; et al. Barley stripe mosaic virus γb protein disrupts chloroplast antioxidant defenses to optimize viral replication. EMBO J. 2021, 40, e107660. [Google Scholar] [CrossRef]
- Liu, S.; Wang, C.; Liu, X.; Navas-Castillo, J.; Zang, L.; Fan, Z.; Zhu, X.; Zhou, T. Tomato chlorosis virus-encoded p22 suppresses auxin signalling to promote infection via interference with SKP1-Cullin-F-box(TIR1) complex assembly. Plant Cell Environ. 2021, 44, 3155–3172. [Google Scholar] [CrossRef]
- Yang, M.; Ismayil, A.; Jiang, Z.; Wang, Y.; Zheng, X.; Yan, L.; Hong, Y.; Li, D.; Liu, Y. A viral protein disrupts vacuolar acidification to facilitate virus infection in plants. EMBO J. 2022, 41, e108713. [Google Scholar] [CrossRef]
- Yang, T.; Qiu, L.; Huang, W.; Xu, Q.; Zou, J.; Peng, Q.; Lin, H.; Xi, D. Chilli veinal mottle virus HCPro interacts with catalase to facilitate virus infection in Nicotiana tabacum. J. Exp. Bot. 2020, 71, 5656–5668. [Google Scholar] [CrossRef]
- Brown, S.L.; Garrison, D.J.; May, J.P. Phase separation of a plant virus movement protein and cellular factors support virus-host interactions. PLoS Pathog. 2021, 17, e1009622. [Google Scholar] [CrossRef]
- Mandadi, K.K.; Scholthof, K.B. Plant immune responses against viruses: How does a virus cause disease? Plant Cell 2013, 25, 1489–1505. [Google Scholar] [CrossRef] [Green Version]
- Wang, A. Dissecting the molecular network of virus-plant interactions: The complex roles of host factors. Annu. Rev. Phytopathol. 2015, 53, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, Y.; Wang, A. Research advances in potyviruses: From the laboratory bench to the field. Annu. Rev. Phytopathol. 2021, 59, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Liu, Y.; Molho, M.; Zhang, S.; Wang, L.; Xie, L.; Nagy, P.D. Co-opting the fermentation pathway for tombusvirus replication: Compartmentalization of cellular metabolic pathways for rapid ATP generation. PLoS Pathog. 2019, 15, e1008092. [Google Scholar] [CrossRef] [Green Version]
- Nagy, P.D.; Lin, W. Taking over cellular energy-metabolism for TBSV replication: The high ATP requirement of an RNA virus within the viral replication organelle. Viruses 2020, 12, 56. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Du, K.; Wang, P.; Wang, X.; Zang, L.; Peng, D.; Chen, X.; Sun, G.; Zhang, H.; Fan, Z.; et al. Sugarcane mosaic virus orchestrates the lactate fermentation pathway to support its successful infection. Front. Plant Sci. 2022, 13, 1099362. [Google Scholar] [CrossRef]
- Shalitin, D.; Wolf, S. Cucumber mosaic virus infection affects sugar transport in melon plants. Plant Physiol. 2000, 123, 597–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhu, O.P.; Annarao, S.; Pathre, U.; Snehi, S.K.; Raj, S.K.; Roy, R.; Tuli, R.; Khetrapal, C.L. Metabolic and histopathological alterations of Jatropha mosaic begomovirus-infected Jatropha curcas L. by HR-MAS NMR spectroscopy and magnetic resonance imaging. Planta 2010, 232, 85–93. [Google Scholar] [CrossRef]
- Lopez-Gresa, M.P.; Lison, P.; Kim, H.K.; Choi, Y.H.; Verpoorte, R.; Rodrigo, I.; Conejero, V.; Belles, J.M. Metabolic fingerprinting of Tomato Mosaic Virus infected Solanum lycopersicum. J. Plant Physiol. 2012, 169, 1586–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, R.; Kathiria, P.; Psychogios, N.; Bouatra, S.; Krishnamurthy, R.; Wishart, D.; Kovalchuk, I. Progeny of tobacco mosaic virus-infected Nicotiana tabacum plants exhibit trans-generational changes in metabolic profiles. Biocatal. Agric. Biotechnol. 2012, 1, 115–123. [Google Scholar] [CrossRef]
- Tecsi, L.I.; Smith, A.M.; Maule, A.J.; Leegood, R.C. A spatial analysis of physiological changes associated with infection of cotyledons of marrow plants with cucumber mosaic virus. Plant Physiol. 1996, 111, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Ballare, C.L. Light regulation of plant defense. Annu. Rev. Plant Biol. 2014, 65, 335–363. [Google Scholar] [CrossRef]
- Kangasjarvi, S.; Neukermans, J.; Li, S.; Aro, E.M.; Noctor, G. Photosynthesis, photorespiration, and light signalling in defence responses. J. Exp. Bot. 2012, 63, 1619–1636. [Google Scholar] [CrossRef] [Green Version]
- Osmond, C.B.; Berry, J.A.; Balachandran, S.; Buchenosmond, C.; Daley, P.F.; Hodgson, R.A.J. Potential consequences of virus-infection for shade-sun accumulation in leaves. Bot. Acta 1990, 103, 226–229. [Google Scholar] [CrossRef]
- Harrison, B.D.; Jones, R.A.C. Effects of light and temperature on symptom development and virus content of tobacco leaves inoculated with potato mop-top virus. Ann. Appl. Biol. 1971, 67, 377. [Google Scholar] [CrossRef]
- Masinde, E.A.; Mkamillo, G.; Ogendo, J.O.; Hillocks, R.; Mulwa, R.M.S.; Kimata, B.; Maruthi, M.N. Genotype by environment interactions in identifying cassava (Manihot esculenta Crantz) resistant to cassava brown streak disease. Field Crops Res. 2018, 215, 39–48. [Google Scholar] [CrossRef]
- Godfrey, S.; Settumba, M.; Samuel, K. Effect of temperature on sweet potato virus disease symptom expression. Afr. J. Agric. Res. 2017, 12, 2295–2309. [Google Scholar] [CrossRef]
- Chung, B.N.; Lee, J.H.; Kang, B.C.; Koh, S.W.; Joa, J.H.; Choi, K.S.; Ahn, J.J. HR-mediated defense response is overcome at high temperatures in capsicum species. Plant Pathol. J. 2018, 34, 71–77. [Google Scholar] [CrossRef]
- del Toro, F.J.; Rakhshandehroo, F.; Larruy, B.; Aguilar, E.; Tenllado, F.; Canto, T. Effects of simultaneously elevated temperature and CO2 levels on Nicotiana benthamiana and its infection by different positive-sense RNA viruses are cumulative and virus type-specific. Virology 2017, 511, 184–192. [Google Scholar] [CrossRef]
- Kumar, P.L.; Selvarajan, R.; Iskra-Caruana, M.L.; Chabannes, M.; Hanna, R. Biology, Etiology, and Control of virus diseases of Banana and Plantain. In Advances in Virus Research, Vol 91: Control of Plant Virus Diseases Vegetatively-Propagated Crops; Loebenstein, G., Katis, N.I., Eds.; Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2015; Volume 91, pp. 229–269. [Google Scholar]
- Qazi, J. Banana bunchy top virus and the bunchy top disease. J. Gen. Plant Pathol. 2016, 82, 2–11. [Google Scholar] [CrossRef]
- Miller, W.A.; Liu, S.J.; Beckett, R. Barley yellow dwarf virus: Luteoviridae or Tombusviridae? Mol. Plant Pathol. 2002, 3, 177–183. [Google Scholar] [CrossRef]
- Walls, J.; Rajotte, E.; Rosa, C. The past, present, and future of barley yellow dwarf management. Agriculture 2019, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Eni, A.O.; Efekemo, O.P.; Onile-ere, O.A.; Pita, J.S. South West and North Central Nigeria: Assessment of cassava mosaic disease and field status ofAfrican cassava mosaic virusandEast African cassava mosaic virus. Ann. Appl. Biol. 2021, 178, 466–479. [Google Scholar] [CrossRef]
- Dawson, W.O.; Garnsey, S.M.; Tatineni, S.; Folimonova, S.Y.; Harper, S.J.; Gowda, S. Citrus tristeza virus-host interactions. Front. Microbiol. 2013, 4, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallitelli, D. The ecology of Cucumber mosaic virus and sustainable agriculture. Virus Res. 2000, 71, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Makkouk, K.; Pappu, H.; Kumari, S.G. Virus diseases of peas, beans, and faba bean in the mediterranean region. In Viruses and Virus Diseases of Vegetables in the Mediterranean Basin; Loebenstein, G., Lecoq, H., Eds.; Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2012; Volume 84, pp. 367–402. [Google Scholar]
- Almeida, R.P.P.; Daane, K.M.; Bell, V.A.; Blaisdell, G.K.; Cooper, M.L.; Herrbach, E.; Pietersen, G. Ecology and management of grapevine leafroll disease. Front. Microbiol. 2013, 4, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naidu, R.; Rowhani, A.; Fuchs, M.; Golino, D.; Martelli, G.P. Grapevine leafroll: A complex viral disease affecting a high-value fruit crop. Plant Disease 2014, 98, 1172–1185. [Google Scholar] [CrossRef]
- Song, Y.S.; Hanner, R.H.; Meng, B.Z. Probing into the effects of grapevine leafroll-associated viruses on the physiology, fruit quality and gene expression of grapes. Viruses 2021, 13, 593. [Google Scholar] [CrossRef]
- Kannan, M.; Ismail, I.; Bunawan, H. Maize dwarf mosaic virus: From genome to disease management. Viruses 2018, 10, 492. [Google Scholar] [CrossRef] [Green Version]
- Emeraghi, M.; Achigan-Dako, E.G.; Nwaoguala, C.N.C.; Oselebe, H. Maize streak virus research in Africa: An end or a crossroad. Theor. Appl. Genet. 2021, 134, 3785–3803. [Google Scholar] [CrossRef]
- Shepherd, D.N.; Martin, D.P.; van der Walt, E.; Dent, K.; Varsani, A.; Rybicki, E.P. Maize streak virus: An old and complex ‘emerging’ pathogen. Mol. Plant Pathol. 2010, 11, 1–12. [Google Scholar] [CrossRef]
- Castillo, X.A.O.; Fermin, G.; Tabima, J.; Rojas, Y.; Tennant, P.F.; Fuchs, M.; Sierra, R.; Bernal, A.J.; Restrepo, S. Phylogeography and molecular epidemiology of Papaya ringspot virus. Virus Res. 2011, 159, 132–140. [Google Scholar] [CrossRef]
- Tripathi, S.; Suzuki, J.Y.; Ferreira, S.A.; Gonsalves, D. Papaya ringspot virus-P: Characteristics, pathogenicity, sequence variability and control. Mol. Plant Pathol. 2008, 9, 269–280. [Google Scholar] [CrossRef]
- Cambra, M.; Capote, N.; Myrta, A.; Llacer, G. Plum pox virus and the estimated costs associated with sharka disease. EPPO Bull. 2006, 36, 202–204. [Google Scholar] [CrossRef]
- Garcia, J.A.; Glasa, M.; Cambra, M.; Candresse, T. Plum pox virus and sharka: A model potyvirus and a major disease. Mol. Plant Pathol. 2014, 15, 226–241. [Google Scholar] [CrossRef]
- Gray, S.; De Boer, S.; Lorenzen, J.; Karasev, A.; Whitworth, J.; Nolte, P.; Singh, R.; Boucher, A.; Xu, H.M. Potato virus Y: An evolving concern for potato crops in the united states and canada. Plant Dis. 2010, 94, 1384–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karasev, A.V.; Gray, S.M. Continuous and emerging challenges of Potato virus Y in potato. Annu. Rev. Phytopathol. 2013, 51, 571–586. [Google Scholar]
- Nihad, S.A.I.; Manidas, A.C.; Kamrul, H.; Hasan, M.A.I.; Omma, H.; Latif, M.A. Genetic variability, heritability, genetic advance and phylogenetic relationship between rice tungro virus resistant and susceptible genotypes revealed by morphological traits and SSR markers. Curr. Plant Biol. 2021, 25, 100194. [Google Scholar] [CrossRef]
- Pinel-Galzi, A.; Traore, O.; Sere, Y.; Hebrard, E.; Fargette, D. The biogeography of viral emergence: Rice yellow mottle virus as a case study. Curr. Opin. Virol. 2015, 10, 7–13. [Google Scholar] [CrossRef]
- Fargette, D.; Pinel, A.; Abubakar, Z.; Traore, O.; Brugidou, C.; Fatogoma, S.; Hebrard, E.; Choisy, M.; Sere, Y.; Fauquet, C.; et al. Inferring the evolutionary history of Rice yellow mottle virus from genomic, phylogenetic, and phylogeographic studies. J. Virol. 2004, 78, 3252–3261. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Xu, D.; Xu, D.; Zhang, M. Southern rice black-streaked dwarf virus: A white-backed planthopper-transmitted fijivirus threatening rice production in Asia. Front. Microbiol. 2013, 4, 270. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Z.Y.; Tian, Y.Y.; Wang, J.; Ismail, R.G.; Bondok, A.; Fan, Z.F. Advances in research on maize lethal necrosis, a devastating viral disease. Phytopathol. Res. 2022, 4, 14. [Google Scholar] [CrossRef]
- Clark, C.A.; Davis, J.A.; Mukasa, S.B.; Abad, J.A.; Tugume, A.K.; Cuellar, W.J.; Fuentes, S.; Kreuze, J.F.; Tairo, F.D.; Gibson, R.W.; et al. Sweetpotato viruses: 15 years of progress on understanding and managing complex diseases. Plant Dis. 2012, 96, 168–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, D.L.; Fuentes, S.; Salazar, L.F. Sweetpotato virus disease (SPVD): Distribution, incidence, and effect on sweetpotato yield in Peru. Plant Dis. 2003, 87, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappu, H.R.; Jones, R.A.C.; Jain, R.K. Global status of tospovirus epidemics in diverse cropping systems: Successes achieved and challenges ahead. Virus Res. 2009, 141, 219–236. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; van Grinsven, I.L.; Kormelink, R.; Tao, X.R. Paving the way to tospovirus infection: Multilined interplays with plant innate immunity. Ann. Rev. Phytopathol. 2019, 57, 41–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Pendon, J.A.; Canizares, M.C.; Moriones, E.; Bejarano, E.R.; Czosnek, H.; Navas-Castillo, J. Tomato yellow leaf curl viruses: Menage a trois between the virus complex, the plant and the whitefly vector. Mol. Plant Pathol. 2010, 11, 441–450. [Google Scholar] [CrossRef]
- Prasad, A.; Sharma, N.; Hari-Gowthem, G.; Muthamilarasan, M.; Prasad, M. Tomato yellow leaf curl virus: Impact, challenges, and management. Trends Plant Sci. 2020, 25, 897–911. [Google Scholar] [CrossRef]
- Coutts, B.A.; Kehoe, M.A.; Jones, R.A.C. Minimising losses caused by Zucchini yellow mosaic virus in vegetable cucurbit crops in tropical, sub-tropical and Mediterranean environments through cultural methods and host resistance. Virus Res. 2011, 159, 141–160. [Google Scholar] [CrossRef]
- Simmons, H.E.; Dunham, J.P.; Zinn, K.E.; Munkvold, G.P.; Holmes, E.C.; Stephenson, A.G. Zucchini yellow mosaic virus (ZYMV, Potyvirus): Vertical transmission, seed infection and cryptic infections. Virus Res. 2013, 176, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhang, X.; Hong, Y.; Liu, Y. Chloroplast in plant-virus interaction. Front. Microbiol. 2016, 7, 1565. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cui, H.; Cui, X.; Wang, A. The altered photosynthetic machinery during compatible virus infection. Curr. Opin. Virol. 2016, 17, 19–24. [Google Scholar] [CrossRef]
- Kong, L.; Wu, J.; Lu, L.; Xu, Y.; Zhou, X. Interaction between Rice stripe virus disease-specific protein and host PsbP enhances virus symptoms. Mol. Plant 2014, 7, 691–708. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, D.; Chakraborty, S. Chloroplast: The Trojan horse in plant-virus interaction. Mol. Plant Pathol. 2018, 19, 504–518. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Zhang, Y.; Wang, C.; Lei, R.; Wu, Y.; Li, X.; Zhu, S. Cucumber mosaic virus coat protein induces the development of chlorotic symptoms through interacting with the chloroplast ferredoxin I protein. Sci. Rep. 2018, 8, 1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, D.; Xu, X.; Yan, Z.; Tettey, C.; Fang, L.; Yang, G.; Geng, C.; Tian, Y.; Li, X. The chloroplast ribosomal protein large subunit 1 interacts with viral polymerase and promotes virus infection. Plant Physiol. 2021, 187, 174–186. [Google Scholar] [CrossRef]
- Tu, Y.Y.; Jin, Y.S.; Ma, D.Y.; Li, H.; Zhang, Z.Q.; Dong, J.L.; Wang, T. Interaction between PVY HC- Pro and the NtCF(1)beta-subunit reduces the amount of chloroplast ATP synthase in virus- infected tobacco. Sci. Rep. 2015, 5, srep15605. [Google Scholar] [CrossRef] [PubMed]
- Angel, C.A.; Lutz, L.; Yang, X.H.; Rodriguez, A.; Adair, A.; Zhang, Y.; Leisner, S.M.; Nelson, R.S.; Schoelz, J.E. The P6 protein of Cauliflower mosaic virus interacts with CHUP1, a plant protein which moves chloroplasts on actin microfilaments. Virology 2013, 443, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Caplan, J.L.; Kumar, A.S.; Park, E.; Padmanabhan, M.S.; Hoban, K.; Modla, S.; Czymmek, K.; Dinesh-Kumar, S.P. Chloroplast stromules function during innate immunity. Dev. Cell 2015, 34, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Jang, C.; Seo, E.Y.; Nam, J.; Bae, H.; Gim, Y.G.; Kim, H.G.; Cho, I.S.; Lee, Z.W.; Bauchan, G.R.; Hammond, J.; et al. Insights into Alternanthera mosaic virus TGB3 functions: Interactions with nicotiana benthamiana PsbO correlate with chloroplast vesiculation and veinal necrosis caused by TGB3 over-expression. Front. Plant Sci. 2013, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Abbink, T.E.; Peart, J.R.; Mos, T.N.; Baulcombe, D.C.; Bol, J.F.; Linthorst, H.J. Silencing of a gene encoding a protein component of the oxygen-evolving complex of photosystem II enhances virus replication in plants. Virology 2002, 295, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundin, B.; Hansson, M.; Schoefs, B.; Vener, A.V.; Spetea, C. The Arabidopsis PsbO2 protein regulates dephosphorylation and turnover of the photosystem II reaction centre D1 protein. Plant J. 2007, 49, 528–539. [Google Scholar] [CrossRef]
- Lundin, B.; Thuswaldner, S.; Shutova, T.; Eshaghi, S.; Samuelsson, G.; Barber, J.; Andersson, B.; Spetea, C. Subsequent events to GTP binding by the plant PsbO protein: Structural changes, GTP hydrolysis and dissociation from the photosystem II complex. Biochim. Biophys. Acta (BBA)—Bioenerg. 2007, 1767, 500–508. [Google Scholar] [CrossRef] [Green Version]
- Bengyella, L.; Waikhom, S.D.; Allie, F.; Rey, C. Virus tolerance and recovery from viral induced-symptoms in plants are associated with transcriptome reprograming. Plant Mol. Biol. 2015, 89, 243–252. [Google Scholar] [CrossRef]
- Lehto, K.; Tikkanen, M.; Hiriart, J.B.; Paakkarinen, V.; Aro, E.M. Depletion of the photosystem II core complex in mature tobacco leaves infected by the flavum strain of Tobacco mosaic virus. Mol. Plant Microbe Interact. 2003, 16, 1135–1144. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.Q.; Liu, Z.M.; Xu, J.; Zhou, T.; Wang, M.; Chen, Y.T.; Li, H.F.; Fan, Z.F. HC-Pro protein of sugarcane mosaic virus interacts specifically with maize ferredoxin-5 in vitro and in planta. J. Gen. Virol. 2008, 89, 2046–2054. [Google Scholar] [CrossRef] [PubMed]
- Hanke, G.; Mulo, P. Plant type ferredoxins and ferredoxin-dependent metabolism. Plant Cell Environ. 2013, 36, 1071–1084. [Google Scholar] [CrossRef] [PubMed]
- Holtgrefe, S.; Bader, K.P.; Horton, P.; Scheibe, R.; von Schaewen, A.; Backhausen, J.E. Decreased content of leaf ferredoxin changes electron distribution and limits photosynthesis in transgenic potato plants. Plant Physiol. 2003, 133, 1768–1778. [Google Scholar] [CrossRef] [Green Version]
- Song, X.S.; Wang, Y.J.; Mao, W.H.; Shi, K.; Zhou, Y.H.; Nogues, S.; Yu, J.Q. Effects of cucumber mosaic virus infection on electron transport and antioxidant system in chloroplasts and mitochondria of cucumber and tomato leaves. Physiol. Plant 2009, 135, 246–257. [Google Scholar] [CrossRef]
- Ma, Y.; Zhou, T.; Hong, Y.; Fan, Z.; Li, H. Decreased level of ferredoxin I in Tobacco mosaic virus-infected tobacco is associated with development of the mosaic symptom. Physiol. Mol. Plant Pathol. 2008, 72, 39–45. [Google Scholar] [CrossRef]
- Lei, R.; Jiang, H.; Hu, F.; Yan, J.; Zhu, S. Chlorophyll fluorescence lifetime imaging provides new insight into the chlorosis induced by plant virus infection. Plant Cell Rep. 2017, 36, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.A.; Eamens, A.L.; Wang, M.B. Viral small interfering RNAs target host genes to mediate disease symptoms in plants. PLoS Pathog. 2011, 7, e1002022. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Ma, D.; Dong, J.; Li, D.; Deng, C.; Jin, J.; Wang, T. The HC-pro protein of potato virus Y interacts with NtMinD of tobacco. Mol. Plant Microbe Interact. 2007, 20, 1505–1511. [Google Scholar] [CrossRef] [Green Version]
- Blazquez, M.A.; Nelson, D.C.; Weijers, D. Evolution of plant hormone response pathways. Annu. Rev. Plant Biol. 2020, 71, 327–353. [Google Scholar] [CrossRef] [Green Version]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Padmanabhan, M.S.; Goregaoker, S.P.; Golem, S.; Shiferaw, H.; Culver, J.N. Interaction of the tobacco mosaic virus replicase protein with the Aux/IAA protein PAP1/IAA26 is associated with disease development. J. Virol. 2005, 79, 2549–2558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.; Qin, Q.; Wang, Y.; Pu, Y.; Liu, L.; Wen, X.; Ji, S.; Wu, J.; Wei, C.; Ding, B.; et al. Rice dwarf virus P2 protein hijacks auxin signaling by directly targeting the rice OsIAA10 protein, enhancing viral infection and disease development. PLoS Pathog. 2016, 12, e1005847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Guo, L.; Lang, X.; Li, M.; Wu, G.; Wu, R.; Wang, L.; Zhao, M.; Qing, L. Geminivirus C4 proteins inhibit GA signaling via prevention of NbGAI degradation, to promote viral infection and symptom development in N. benthamiana. PLoS Pathog. 2022, 18, e1010217. [Google Scholar] [CrossRef]
- Zhao, S.; Li, Y. Current understanding of the interplays between host hormones and plant viral infections. PLoS Pathog. 2021, 17, e1009242. [Google Scholar] [CrossRef]
- Zhu, S.; Gao, F.; Cao, X.; Chen, M.; Ye, G.; Wei, C.; Li, Y. The rice dwarf virus P2 protein interacts with ent-kaurene oxidases in vivo, leading to reduced biosynthesis of gibberellins and rice dwarf symptoms. Plant Physiol. 2005, 139, 1935–1945. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Okada, K.; Fukazawa, J.; Takahashi, Y. DELLA-dependent and -independent gibberellin signaling. Plant Signal Behav. 2018, 13, e1445933. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Qin, L.; Lee, S.; Fu, X.; Richards, D.E.; Cao, D.; Luo, D.; Harberd, N.P.; Peng, J. Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function. Development 2004, 131, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Van Breusegem, F.; Dat, J.F. Reactive oxygen species in plant cell death. Plant Physiol. 2006, 141, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Berlett, B.S.; Stadtman, E.R. Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef] [Green Version]
- Montillet, J.L.; Chamnongpol, S.; Rusterucci, C.; Dat, J.; van de Cotte, B.; Agnel, J.P.; Battesti, C.; Inze, D.; Van Breusegem, F.; Triantaphylides, C. Fatty acid hydroperoxides and H2O2 in the execution of hypersensitive cell death in tobacco leaves. Plant Physiol. 2005, 138, 1516–1526. [Google Scholar] [CrossRef] [Green Version]
- Riedle-Bauer, M. Role of Reactive oxygen species and antioxidant enzymes in systemic virusInfections of plants. J. Phytopathol. 2000, 148, 297–302. [Google Scholar] [CrossRef]
- Inaba, J.-i.; Kim, B.M.; Shimura, H.; Masuta, C. Virus-induced necrosis is a consequence of direct protein-protein interaction between a viral RNA-silencing suppressor and a host catalase. Plant Physiol. 2011, 156, 2026–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Z.; Zhao, Z.; Chen, L.; Li, M.; Zhou, T.; Deng, C.; Zhou, Q.; Fan, Z. Synergistic infection of two viruses MCMV and SCMV increases the accumulations of both MCMV and MCMV-derived siRNAs in maize. Sci. Rep. 2016, 6, 20520. [Google Scholar] [CrossRef]
- Lin, K.Y.; Wu, S.Y.; Hsu, Y.H.; Lin, N.S. MiR398-regulated antioxidants contribute to Bamboo mosaic virus accumulation and symptom manifestation. Plant Physiol. 2021, 188, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Inze, D.; De Veylder, L. Cell cycle regulation in plant development. Annu. Rev. Genet. 2006, 40, 77–105. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.J.; Orozco, B.M.; Roe, J.L.; Nagar, S.; Ou, S.; Feiler, H.S.; Durfee, T.; Miller, A.B.; Gruissem, W.; Robertson, D.; et al. A geminivirus replication protein interacts with the retinoblastoma protein through a novel domain to determine symptoms and tissue specificity of infection in plants. EMBO J. 2000, 19, 3485–3495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronson, M.N.; Meyer, A.D.; Györgyey, J.; Katul, L.; Vetten, H.J.; Gronenborn, B.; Timchenko, T. Clink, a nanovirus-encoded protein, binds both pRB and SKP1. J. Virol. 2000, 74, 2967–2972. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.Z.; Yang, X.L.; Huang, C.J.; Zhang, X.R.; Zhou, X.P. Tomato leaf curl Yunnan virus-encoded C4 induces cell division through enhancing stability of Cyclin D 1.1 via impairing NbSKη-mediated phosphorylation in Nicotiana benthamiana. PLoS Pathog. 2018, 14, e1006789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desvoyes, B.; Gutierrez, C. Roles of plant retinoblastoma protein: Cell cycle and beyond. EMBO J. 2020, 39, e105802. [Google Scholar] [CrossRef] [PubMed]
- Gordon-Kamm, W.; Dillces, B.P.; Lowe, K.; Hoerster, G.; Sun, X.; Ross, M.; Church, L.; Bunde, C.; Farrell, J.; Sykes, L.; et al. Stimulation of the cell cycle and maize transformation by disruption of the plant retinoblastoma pathway. Proc. Natl. Acad. Sci. USA 2002, 99, 11975–11980. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Chen, H.; Teng, K.; Zhao, Q.; Zhang, Z.; Li, Y.; Liang, L.; Xia, R.; Wu, Y.; Guo, H.; et al. RKP, a RING finger E3 ligase induced by BSCTV C4 protein, affects geminivirus infection by regulation of the plant cell cycle. Plant J. 2009, 57, 905–917. [Google Scholar] [CrossRef]
- Dewitte, W.; Riou-Khamlichi, C.; Scofield, S.; Healy, J.M.; Jacqmard, A.; Kilby, N.J.; Murray, J.A. Altered cell cycle distribution, hyperplasia, and inhibited differentiation in Arabidopsis caused by the D-type cyclin CYCD3. Plant Cell 2003, 15, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Nakagami, H.; Kawamura, K.; Sugisaka, K.; Sekine, M.; Shinmyo, A. Phosphorylation of retinoblastoma-related protein by the cyclin D/cyclin-dependent kinase complex is activated at the G1/S-phase transition in tobacco. Plant Cell 2002, 14, 1847–1857. [Google Scholar] [CrossRef] [Green Version]
- Dewitte, W.; Scofield, S.; Alcasabas, A.A.; Maughan, S.C.; Menges, M.; Braun, N.; Collins, C.; Nieuwland, J.; Prinsen, E.; Sundaresan, V.; et al. Arabidopsis CYCD3 D-type cyclins link cell proliferation and endocycles and are rate-limiting for cytokinin responses. Proc. Natl. Acad. Sci. USA 2007, 104, 14537–14542. [Google Scholar] [CrossRef]
- Caplan, J.L.; Zhu, X.; Mamillapalli, P.; Marathe, R.; Anandalakshmi, R.; Dinesh-Kumar, S.P. Induced ER chaperones regulate a receptor-like kinase to mediate antiviral innate immune response in plants. Cell Host Microbe 2009, 6, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Di Carli, M.; Benvenuto, E.; Donini, M. Recent insights into plant-virus interactions through proteomic analysis. J. Proteome Res. 2012, 11, 4765–4780. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.R.; Zhou, Q.; Yang, J.M.; Guo, H.X.; Yang, L.J.; Liu, W.Q. iTRAQ protein profile analysis provides integrated insight into mechanisms of tolerance to TMV in tobacco (Nicotiana tabacum). J. Proteomics 2016, 132, 21–30. [Google Scholar] [CrossRef]
- Seo, J.K.; Kim, M.K.; Kwak, H.R.; Choi, H.S.; Nam, M.; Choe, J.; Choi, B.; Han, S.J.; Kang, J.H.; Jung, C. Molecular dissection of distinct symptoms induced by tomato chlorosis virus and tomato yellow leaf curl virus based on comparative transcriptome analysis. Virology 2018, 516, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Du, K.; Jiang, T.; Chen, H.; Murphy, A.M.; Carr, J.P.; Du, Z.; Li, X.; Fan, Z.; Zhou, T. Viral perturbation of alternative splicing of a host transcript benefits infection. Plant Physiol. 2020, 184, 1514–1531. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.S.; Nagy, P.D. Direct inhibition of tombusvirus plus-strand RNA synthesis by a dominant negative mutant of a host metabolic enzyme, glyceraldehyde-3-phosphate dehydrogenase, in yeast and plants. J. Virol. 2011, 85, 9090–9102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, C.K.; Prasanth, K.R.; Nagy, P.D. The glycolytic pyruvate kinase is recruited directly into the viral replicase complex to generate ATP for RNA synthesis. Cell Host Microbe 2017, 22, 639–652.e7. [Google Scholar] [CrossRef]
- Otulak, K.; Chouda, M.; Bujarski, J.; Garbaczewska, G. The evidence of Tobacco rattle virus impact on host plant organelles ultrastructure. Micron 2015, 70, 7–20. [Google Scholar] [CrossRef]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef]
- Kogovsek, P.; Pompe-Novak, M.; Petek, M.; Fragner, L.; Weckwerth, W.; Gruden, K. Primary Metabolism, phenylpropanoids and antioxidant pathways are regulated in potato as a response to potato virus Y infection. PLoS ONE 2016, 11, e0146135. [Google Scholar] [CrossRef]
- Hyodo, K.; Okuno, T. Hijacking of host cellular components as proviral factors by plant-infecting viruses. Adv. Virus Res. 2020, 107, 37–86. [Google Scholar] [CrossRef]
- Sumbria, D.; Berber, E.; Mathayan, M.; Rouse, B.T. Virus infections and host metabolism-can we manage the interactions? Front. Immunol. 2020, 11, 594963. [Google Scholar] [CrossRef]
- Moon, J.Y.; Park, J.M. Cross-talk in viral defense signaling in plants. Front. Microbiol. 2016, 07, 2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, T.; Fernie, A.R. The use of metabolomics to dissect plant responses to abiotic stresses. Cell Mol. Life Sci. 2012, 69, 3225–3243. [Google Scholar] [CrossRef] [Green Version]
- Babu, M.; Griffiths, J.S.; Huang, T.S.; Wang, A. Altered gene expression changes in Arabidopsis leaf tissues and protoplasts in response to Plum pox virus infection. BMC Genom. 2008, 9, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Yadav, C.B.; Alatar, A.A.; Faisal, M.; Jyothsna, P.; Malathi, V.G.; Praveen, S. Gene expression changes in tomato during symptom development in response to leaf curl virus infection. J. Plant Biochem. Biotechnol. 2014, 24, 347–354. [Google Scholar] [CrossRef]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2018, 220, 692–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Du, Z.X.; Kong, J.; Chen, L.N.; Qiu, Y.H.; Li, G.F.; Meng, X.H.; Zhu, S.F. Transcriptome analysis of Nicotiana tabacum infected by Cucumber mosaic virus during systemic symptom development. PLoS ONE 2012, 7, e43447. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.B.; Lu, T.C.; Wang, H.X.; Shen, J.; Bu, T.T.; Chao, Q.; Gao, Z.F.; Zhu, X.G.; Wang, Y.F.; Wang, B.C. Posttranslational modification of maize chloroplast pyruvate orthophosphate dikinase reveals the precise regulatory mechanism of its enzymatic activity. Plant Physiol. 2014, 165, 534–549. [Google Scholar] [CrossRef] [Green Version]
- Chastain, C. Structure, Function, and Post-Translational Regulation of C4 Pyruvate Orthophosphate Dikinase; Raghavendra, A.S., Sage, R.F., Eds.; Springer Science + Business Media: Berlin, Germany, 2011; pp. 301–315. [Google Scholar]
- Ludwig, M. The roles of organic acids in C4 photosynthesis. Front. Plant Sci. 2016, 7, 647. [Google Scholar] [CrossRef] [Green Version]
- Selman, I.W.; Grant, S.A. The influence of temperature and daylength on spotted wilt virus disease of tomato. Ann. Appl. Biol. 1957, 45, 312–317. [Google Scholar] [CrossRef]
- Kassanis, B. Some effects of high temperature on the susceptibility of plants to infection with viruses. Ann. Appl. Biol. 1952, 39, 358–369. [Google Scholar] [CrossRef]
- Wosula, E.N.; Tatineni, S.; Wegulo, S.N.; Hein, G.L. Effect of temperature on wheat streak mosaic disease development in winter wheat. Plant Dis. 2017, 101, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, B.N.; Canto, T.; Tenllado, F.; Choi, K.S.; Joa, J.H.; Ahn, J.J.; Kim, C.H.; Do, K.S. The effects of high temperature on infection by potato virus Y, potato virus A, and potato leafroll virus. Plant Pathol. J. 2016, 32, 321–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Li, Y.; Chen, L.; Zhu, L.; Ren, H.; Lin, H.; Xi, D. Temperature dependent defence of Nicotiana tabacum against Cucumber mosaic virus and recovery occurs with the formation of dark green islands. J. Plant Biol. 2016, 59, 293–301. [Google Scholar] [CrossRef]
- Lim, H.S.; Nam, J.; Seo, E.Y.; Nam, M.; Vaira, A.M.; Bae, H.; Jang, C.Y.; Lee, C.H.; Kim, H.G.; Roh, M.; et al. The coat protein of Alternanthera mosaic virus is the elicitor of a temperature-sensitive systemic necrosis in Nicotiana benthamiana, and interacts with a host boron transporter protein. Virology 2014, 452–453, 264–278. [Google Scholar] [CrossRef] [Green Version]
- Vasquez, D.F.; Hernandez, A.; Torres, D.; Borrero-Echeverry, F.; Zuluaga, P.; Rincon, D.F. Drought as a modulator of plant–virus–vector interactions: Effects on symptom expression, plant immunity and vector behaviour. Plant Pathol. 2022, 71, 1282–1292. [Google Scholar] [CrossRef]
- Prasad, A.; Sett, S.; Prasad, M. Plant-virus-abiotic stress interactions: A complex interplay. Environ. Exp. Bot. 2022, 199, 104869. [Google Scholar] [CrossRef]
- Bilgin, D.D.; Aldea, M.; O’Neill, B.F.; Benitez, M.; Li, M.; Clough, S.J.; DeLucia, E.H. Elevated ozone alters soybean-virus interaction. Mol. Plant Microbe Interact. 2008, 21, 1297–1308. [Google Scholar] [CrossRef] [Green Version]
- Vanhaelewyn, L.; Van Der Straeten, D.; De Coninck, B.; Vandenbussche, F. Ultraviolet radiation from a plant perspective: The plant-microorganism context. Front. Plant Sci. 2020, 11, 597642. [Google Scholar] [CrossRef]
- Tatineni, S.; Hein, G.L. Plant viruses of agricultural importance: Current and future perspectives of virus disease management strategies. Phytopathology 2023, 113, 117–141. [Google Scholar] [CrossRef]
- Hinge, V.R.; Chavhan, R.L.; Kale, S.P.; Suprasanna, P.; Kadam, U.S. Engineering resistance against viruses in field crops using CRISPR- Cas9. Curr. Genom. 2021, 22, 214–231. [Google Scholar] [CrossRef]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of plant viruses and disease management: Relevance of genetic diversity and evolution. Front. Plant Sci. 2020, 11, 1092. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, T.; Zhou, T. Unraveling the Mechanisms of Virus-Induced Symptom Development in Plants. Plants 2023, 12, 2830. https://doi.org/10.3390/plants12152830
Jiang T, Zhou T. Unraveling the Mechanisms of Virus-Induced Symptom Development in Plants. Plants. 2023; 12(15):2830. https://doi.org/10.3390/plants12152830
Chicago/Turabian StyleJiang, Tong, and Tao Zhou. 2023. "Unraveling the Mechanisms of Virus-Induced Symptom Development in Plants" Plants 12, no. 15: 2830. https://doi.org/10.3390/plants12152830