
Improving Enzymatic Saccharification of Peach Palm (Bactris gasipaes) Wastes via Biological Pretreatment with Pleurotus ostreatus
 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Results and Discussion
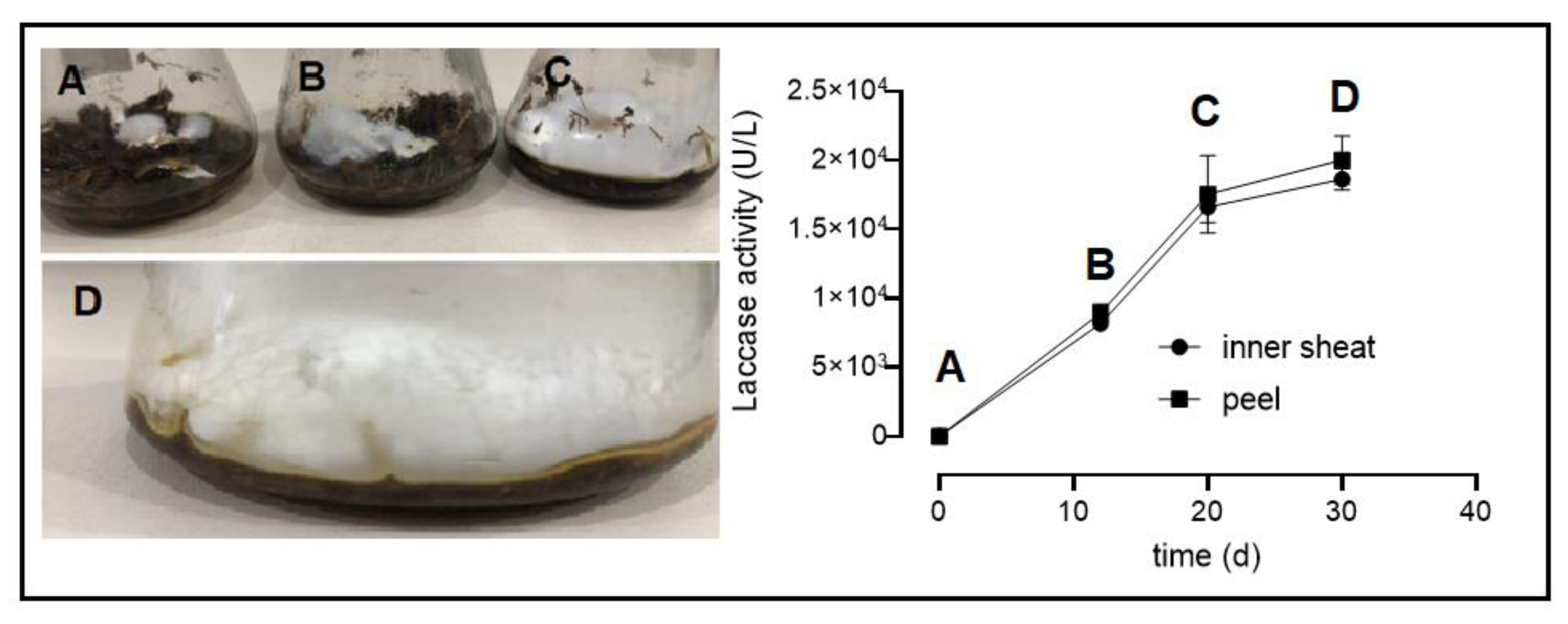
2.1. Growth of P. ostreatus and Enzyme Production
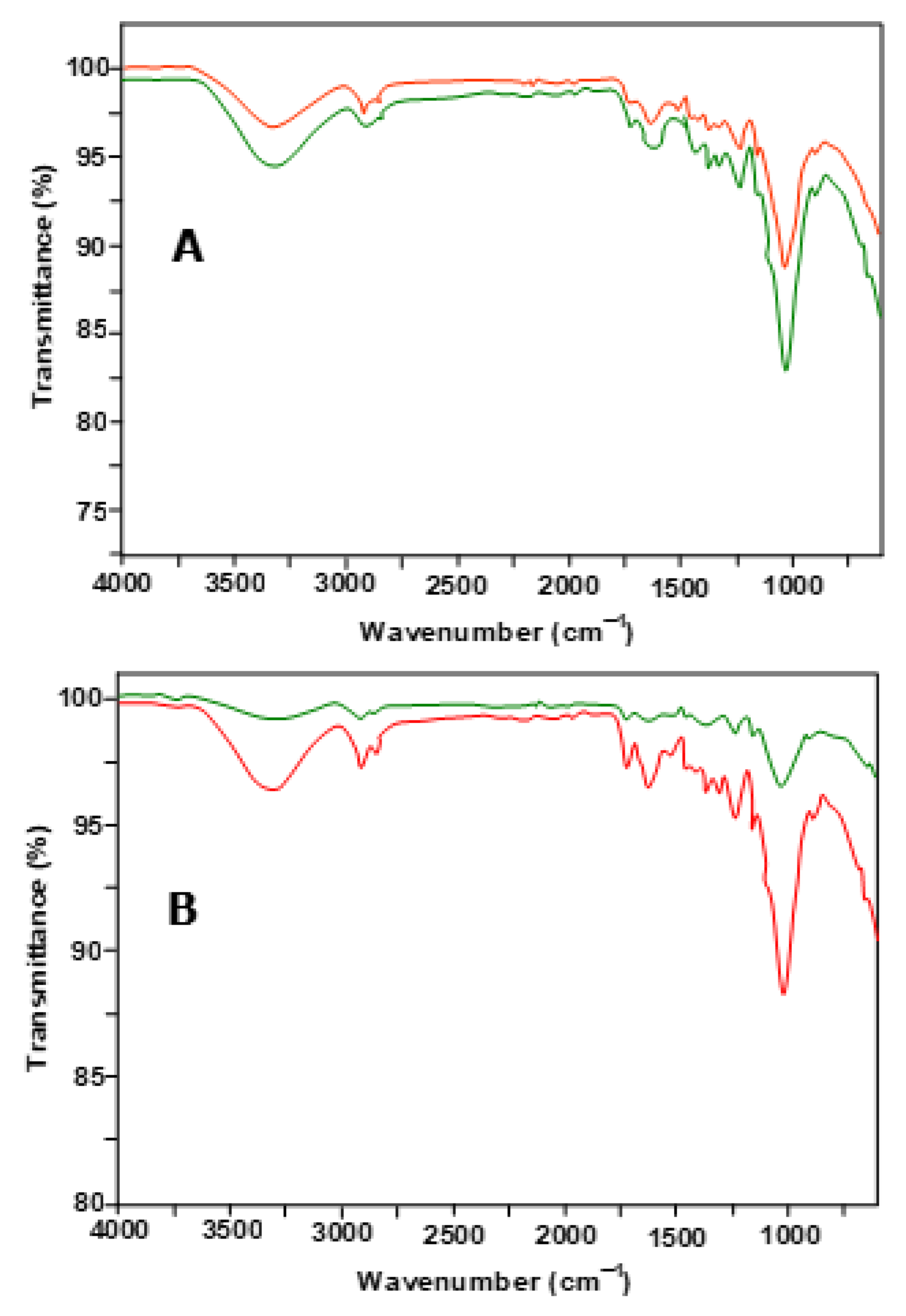
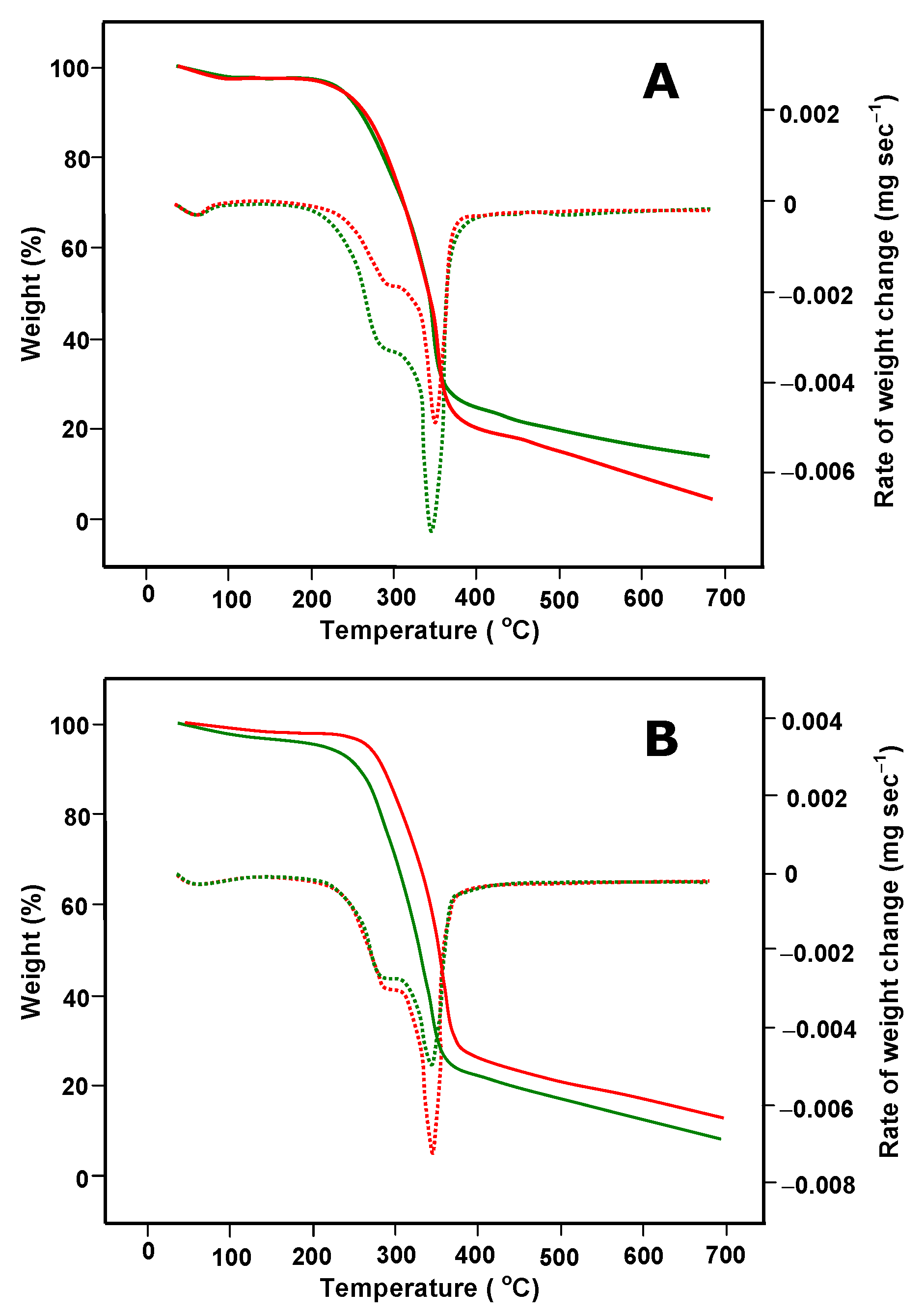
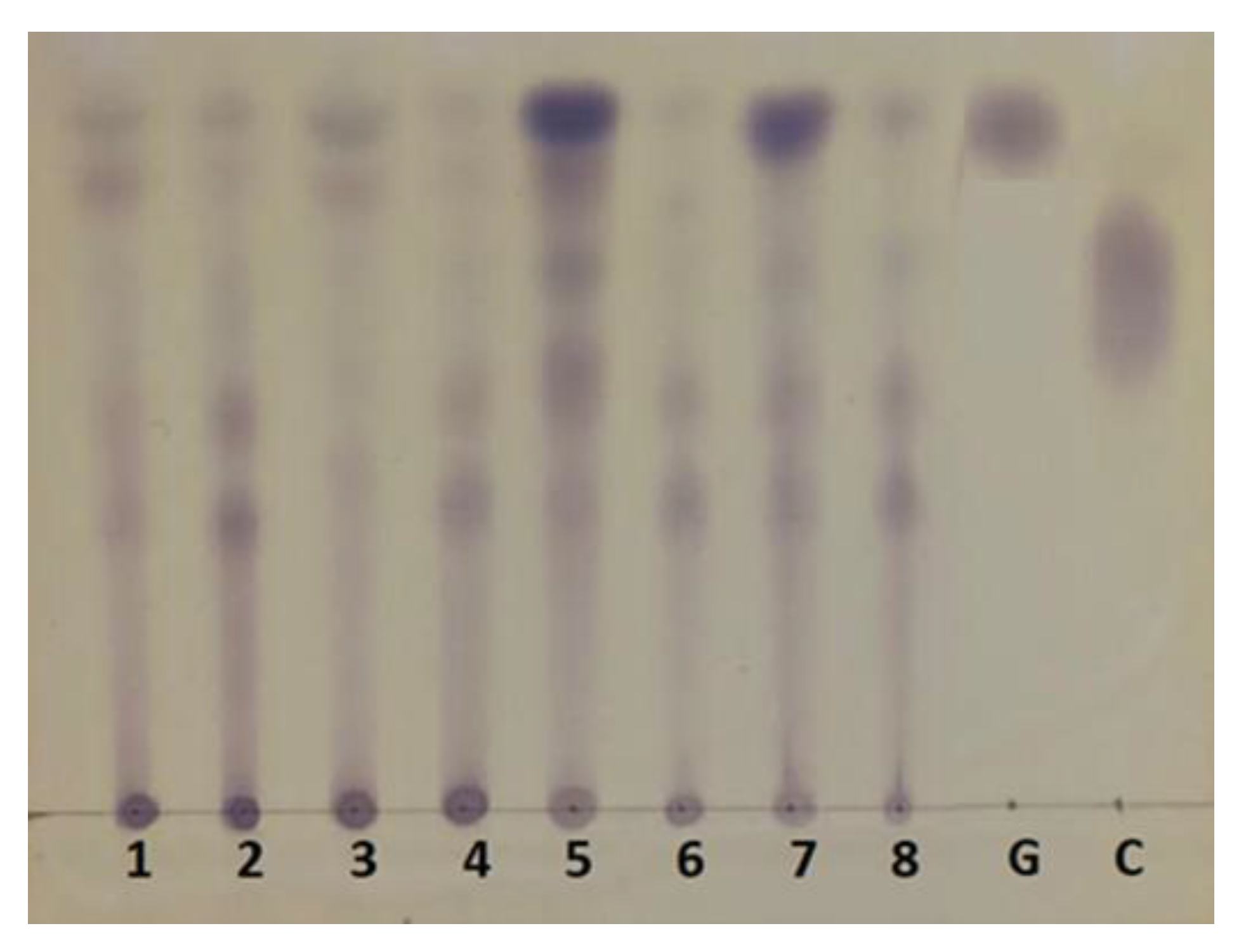
2.2. Chemical and Physical Characterization of Untreated and P. ostreatus-Pretreated Peach Pulp Waste
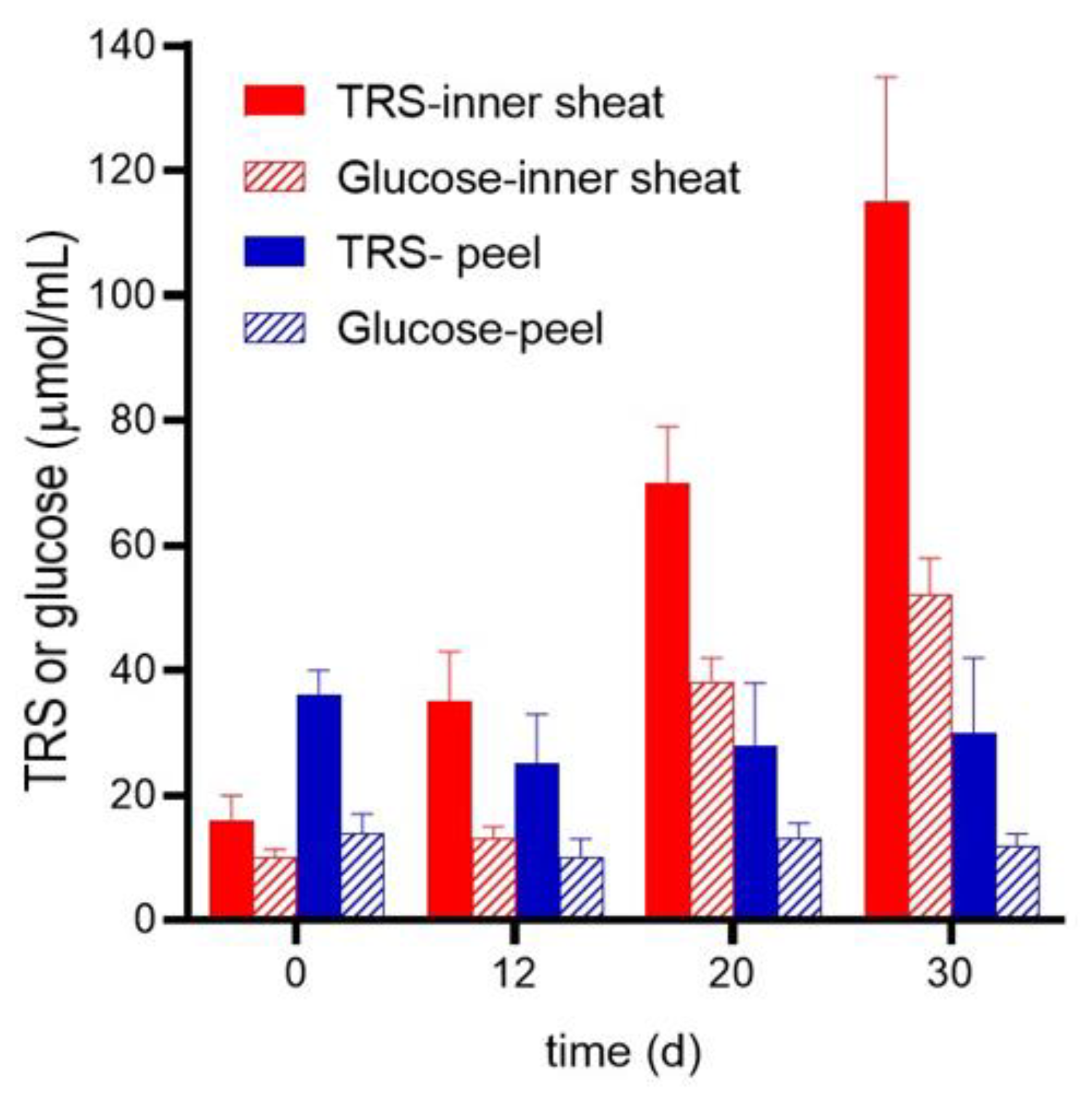
2.3. Enzymatic Hydrolysis of B. gasipaes Inner Sheath and Peel with and without Biological Pretreatment
2.4. Additional Relevant Aspects to Be Considered
3. Materials and Methods
3.1. Microorganism
3.2. Preparation and Characterization of Peach Palm Wastes (Inner Sheath and Peel)
3.3. Biological Pretreatment of Peach Palm Waste Using P. ostreatus
3.4. Laccase and Mn Peroxidase Assays
3.5. Characterization of Untreated and Treated Lignocellulose
3.6. Saccharification of Untreated and Pre-Treated Peach Palm Waste
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Meenakshisundaram, S.; Fayeulle, A.; Leonard, E.; Ceballos, C.; Pauss, A. Fiber degradation and carbohydrate production by combined biological and chemical/physicochemical pretreatment methods of lignocellulosic biomass—A review. Bioresour. Technol. 2021, 331, 125053. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, M.C.S.; Ramos, C.L.; Kudds, M.; Rodriguez-Couto, S.; Srivastava, N.; Ramteke, P.W.; Mishra, P.K.; Molina, G. Enzymatic potential for the valorization of agro-industrial by-products. Biotechnol. Lett. 2020, 42, 1799–1827. [Google Scholar] [CrossRef]
- Zhou, M.; Tian, X. Development of different pretreatments and related Technologies for efficient biomass conversion of lignocellulose. Int. J. Biol. Macromol. 2022, 202, 256–268. [Google Scholar] [CrossRef]
- Matos, K.A.N.; Lima, D.P.; Barbosa, A.P.P.; Mercadante, A.Z.; Chisté, R.S. Peels of tucumã (Astrocaryum vulgare) and peach palm (Bactris gasipaes) are by-products classified as very high carotenoid sources. Food Chem. 2019, 272, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Spacki, K.C.; Corrêa, R.C.G.; Uber, T.M.; Barros, L.; Ferreira, I.C.F.R.; Peralta, R.A.; Moreira, R.F.P.M.; Helm, C.V.; Lima, E.A.; Bracht, A.; et al. Full exploitation of peach palm (Bactris gasipaes Kunth): State of the art and perspectives. Plants 2022, 11, 3175. [Google Scholar] [CrossRef]
- Vieira, T.F.; Corrêa, R.C.G.; Moreira, R.F.P.M.; Peralta, R.A.; de Lima, E.A.; Helm, C.V.; Garcia, J.A.A.; Bracht, A.; Peralta, R.M. Valorization of peach palm (Bactris gasipaes Kunth) waste: Production of antioxidant xylooligosaccharides. Waste Biomass Valorization 2021, 12, 727–740. [Google Scholar] [CrossRef]
- Franco, T.S.; Potulski, D.C.; Viana, L.C.; Forville, E.; Andrade, A.S.; Muniz, G.I.B. Nanocellulose obtained from residues of peach palm extraction (Bactris gasipaes). Carbohydr. Polym. 2019, 218, 8–19. [Google Scholar] [CrossRef]
- Adewuji, A. Underutilized lignocellulosic waste as sources of feedstock for biofuel production in developing countries. Front. Energy Res. 2022, 10, 741570. [Google Scholar] [CrossRef]
- Delmas, G.-H.; Banoud, J.H.; Delmas, M. Lignocellulosic biomass refining: A review promoting a method to produce sustainable hydrogen, fuels, and products. Waste Biomass Valorization 2022, 13, 2477–2491. [Google Scholar] [CrossRef]
- Sharma, P.; Gaur, V.K.; Kim, S.H.; Pandey, A. Microbial strategies for bio-transforming food waste into resources. Bioresour. Technol. 2020, 299, 122580. [Google Scholar] [CrossRef]
- Li, X.; Shi, Y.; Kong, W.; Wei, J.; Song, W.; Wang, S. Improving enzymatic hydrolysis of lignocellulosic biomass by bio-coordinated physicochemical pretreatment—A review. Energy Rep. 2022, 8, 696–709. [Google Scholar] [CrossRef]
- Mankar, A.R.; Pandey, A.; Modak, A.; Pant, K.K. Pretreatment of lignocellulosic biomass: A review on recent advances. Bioresour. Technol. 2021, 334, 125235. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xie, H.; Chen, W.; Wang, E.; Du, F.; Song, A. Biological pretreatment of corn stover with ligninolytic enzyme for high efficient enzymatic hydrolysis. Bioresour. Technol. 2013, 144, 572–578. [Google Scholar] [CrossRef]
- Castoldi, R.; Morais, G.R.; Baesso, M.L.; Correa, R.C.G.; Peralta, R.A.; Peralta Muniz Moreira, R.F.; Polizeli, M.L.T.M.; Souza, C.G.M.; Peralta, R.M. Biological pretreatment of Eucalyptus grandis sawdust with white-rot fungi: Study of degradation patterns and saccharification kinetics. Chem. Eng. J. 2014, 258, 240–246. [Google Scholar] [CrossRef]
- Masran, R.; Zanirun, Z.; Bahrin, E.K.; Ibrahim, M.F.; Yee, P.L.; Abd-Aziz, S. Harnessing the potential of ligninolytic enzymes for lignocellulosic biomass pretreatment. Appl. Microbiol. Biotechnol. 2016, 100, 5231–5249. [Google Scholar] [CrossRef]
- Sindhu, R.; Binod, P.; Pandey, A. Biological pretreatment of lignocellulosic biomass—An overview. Bioresour. Technol. 2016, 199, 76–82. [Google Scholar] [CrossRef]
- Wagner, A.O.; Lackner, N.; Mutschlechner, M.; Prem, E.M.; Mark, R.; IIImer, P. Biological pretreatments for second-generation lignocellulosic resources to enhance biogas production. Energies 2018, 11, 1797. [Google Scholar] [CrossRef]
- Van Kuijk, S.J.A.; Sonnenberg, A.S.M.; Baars, J.J.P.; Hendriks, W.H.; Cone, J.W. Fungal treatment of lignocellulosic biomass: Importance of fungal species, colonization and time on chemical composition and in vitro rumen degradability. Anim. Feed Sci. Technol. 2015, 209, 40–50. [Google Scholar] [CrossRef]
- Durán-Aranguren, D.D.; Meléndez-Melo, J.P.; Covo-Ospina, M.C.; Díaz-Rendón, J.; Reyes-Gutiérrez, D.N.; Reina, L.C.; Durán-Sequeda, D.; Sierra, R. Biological pretreatment of fruit residues using the genus Pleurotus: A review. Bioresour. Technol. Rep. 2021, 16, 100849. [Google Scholar] [CrossRef]
- Freitas, E.N.; Bubna, G.A.; Brugnari, T.; Kato, C.G.; Nolli, M.; Rauen, T.G.; Peralta Muniz Moreira, R.F.; Peralta, R.A.; Bracht, A.; Souza, C.G.M.; et al. Removal of bisphenol A by laccases from Pleurotus ostreatus and Pleurotus pulmonarius and evaluation of ecotoxicity of degradation products. Chem. Eng. J. 2017, 330, 1361–1369. [Google Scholar] [CrossRef]
- Corrêa, R.C.G.; Brugnari, T.; Bracht, A.; Peralta, R.M.; Ferreira, I.C.F.R. Biotechnological, nutritional, and therapeutic uses of Pleurotus spp. (Oyster mushroom) related with its chemical composition: A review on the past decade findings. Trends Food Sci. Technol. 2016, 50, 103–117. [Google Scholar] [CrossRef] [Green Version]
- Krishna, C. Solid-state fermentation systems—An overview. Crit. Rev. Biotechnol. 2005, 25, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Freitas, E.N.; Alnoch, R.C.; Contato, A.G.; Nogueira, K.M.V.; Crevelin, E.J.; Moraes, L.A.B.; Silva, R.N.; Martinez, C.A.; Polizeli, M.L.T.M. Enzymatic pretreatment with laccases from Lentinus sajor-caju induces structural modification in lignin and enhances the digestibility of tropical forage grass (Panicum maximum) grown under future climate conditions. Int. J. Mol. Sci. 2021, 22, 9445. [Google Scholar] [CrossRef]
- Malhotra, M.; Suman, S.K. Laccase-mediated delignification, and detoxification of lignocellulosic biomass: Removing obstacles in energy generation. Environ. Sci. Pollut. Res. 2021, 28, 58929–58944. [Google Scholar] [CrossRef]
- Singh, G.; Kumar, S.; Afreen, S.; Bhalla, A.; Khurana, J.; Chandel, S.; Aggarwal, A.; Arya, S.A. Laccase mediated delignification of wasted and non-food agricultural biomass: Recent developments and challenges. Int. J. Biol. Macromol. 2023, 235, 123840. [Google Scholar] [CrossRef] [PubMed]
- Rencoret, J.; Pereira, A.; del Río, J.C.; Martínez, A.T.; Gutiérrez, A. Delignification and saccharification enhancement of sugarcane byproducts by a laccase-based pretreatment. ACS Sustain. Chem. Eng. 2017, 5, 7145–7154. [Google Scholar] [CrossRef]
- Muktham, R.; Bhargava, S.K.; Bankupalli, S.; Ball, A.S. A review on 1st and 2nd generation bioethanol production-recent progress. J. Sustain. Bioenergy Syst. 2016, 6, 72–92. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, P.S.; Barbosa, M.H.P.; Faria, B.F.H.; Teófilo, R.H. Improvements in the extractive and carbohydrate analysis of sugarcane bagasse. Waste Biomass Valorization 2021, 12, 3727–3740. [Google Scholar] [CrossRef]
- Shamsollahi, Z.; Partovinia, A. Recent advances on pollutants removal by rice husk as a bio-based adsorbent: A critical review. J. Environ. Manag. 2019, 246, 314–323. [Google Scholar] [CrossRef]
- Ghaffar, S.H. Wheat straw biorefinery for agricultural waste valorisation. Green Mat. 2020, 8, 60–67. [Google Scholar] [CrossRef]
- Casas, M.V.; Alonso, M.; Oliet, E.; Rojo, F.; Rodríguez, F. FTIR analysis of lignin regenerated from Pinus radiata and Eucalyptus globulus woods dissolved in imidazolium-based ionic liquids. J. Chem. Technol. Biotechnol. 2012, 87, 472–480. [Google Scholar] [CrossRef]
- Casas, A.; Oliet, M.; Alonso, M.V.; Rodriguez, F. Dissolution of Pinus radiata and Eucalyptus globulus woods in ionic liquids under microwave radiation: Lignin regeneration and characterization. Sep. Purif. Technol. 2012, 97, 115–122. [Google Scholar] [CrossRef]
- Saha, K.; Dasgupta, J.; Chakraborty, S.; Antunes, F.A.F.; Sikder, J.; Curcio, S.; dos Santos, J.C.; Arafat, H.A.; da Silva, S.S. Optimization of lignin recovery from sugarcane bagasse using ionic liquid aided pretreatment. Cellulose 2017, 24, 3191–3207. [Google Scholar] [CrossRef]
- Arun, Y.; Perumal, E.M.; Prakash, K.A.; Rajesh, M.; Tamilarasan, K. Sequential fractionation and characterization of lignin and cellulose fiber from waste rice bran. J. Environ. Chem. Eng. 2020, 8, 104124. [Google Scholar] [CrossRef]
- Salim, R.; Asik, J.; Sarjadi, M.S. Chemical functional groups of extractives, cellulose and lignin extracted from native Leucaena leucocephala bark. Wood Sci. Technol. 2021, 55, 295–313. [Google Scholar] [CrossRef]
- Castoldi, R.; Correa, V.G.; Morais, G.R.; Souza, C.G.M.; Bracht, A.; Peralta, R.A.; Peralta Muniz Moreira, R.F.; Peralta, R.M. Liquid nitrogen pretreatment of eucalyptus sawdust and rice hull for enhanced enzymatic saccharification. Bioresour. Technol. 2017, 224, 648–655. [Google Scholar] [CrossRef]
- Taniguchi, M.; Suzuki, H.; Watanabe, D.; Sakai, K.; Hoshino, K.; Tanaka, T. Evaluation of pretreatment with Pleurotus ostreatus for enzymatic hydrolysis of rice straw. J. Biosci. Bioeng. 2005, 100, 637–643. [Google Scholar] [CrossRef]
- Sun, F.H.; Li, J.; Yuan, Y.X.; Yan, Z.Y. Effect of biological pretreatment with Trametes hirsuta yj9 on enzymatic hydrolysis of corn stover. Int. Biodeterior. Biodegrad. 2011, 65, 931–938. [Google Scholar] [CrossRef]
- Ooshima, H.; Sakata, M.; Harano, Y. Enhancement of enzymatic hydrolysis of cellulose by surfactant. Biotechnol. Bioeng. 1986, 28, 1727–1734. [Google Scholar] [CrossRef]
- Hemraj-Benny, T.; Bandosz, T.J.; Wong, S.S. Effect of ozonolysis on the pore structure, surface chemistry, and bundling of single-walled carbon nanotubes. J. Colloid Interface Sci. 2008, 317, 375–382. [Google Scholar] [CrossRef]
- Singh, S.; Varanasi, P.; Singh, P.; Adams, P.D.; Auer, M.; Simmons, B.A. Understanding the impact of ionic liquid pretreatment on cellulose and lignin via thermochemical analysis. Biomass Bioenergy 2013, 54, 276–283. [Google Scholar] [CrossRef]
- Singh, S.; Cheng, G.; Sathitsuksanoh, N.; Wu, D.; Varansi, P.; George, A.; Balan, V.; Gao, X.; Kumar, R.; Dale, B.E.; et al. Comparison of different biomass pretreatment techniques and their impact on chemistry and structure. Front. Energy Res. 2015, 2, 62. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Feng, L.; Wang, D.; Zhang, R.; Liu, G.; Cheng, G. Thermogravimetric analysis of lignocellulosic biomass with ionic liquid pretreatment. Bioresour. Technol. 2014, 153, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, H.; Huang, H.; Liu, Y. Evaluation of biological pretreatment with white rot fungi for the enzymatic hydrolysis of bamboo culms. Int. Biodeterior. Biodegrad. 2007, 60, 159–164. [Google Scholar] [CrossRef]
- Andersen, S.L.F.; Castoldi, R.; Garcia, J.A.A.; Bracht, A.; Peralta, R.A.; Lima, E.A.; Helm, C.V.; Peralta Muniz Moreira, R.F.; Peralta, R.M. Improving enzymatic saccharification of Eucalyptus grandis branches by ozone pretreatment. Wood Sci. Technol. 2019, 53, 49–69. [Google Scholar] [CrossRef]
- Zhao, X.; Meng, X.; Rogauskas, A.J.; Lai, C.; Ling, Z.; Huang, C.; Yong, Q. Unlocking the secret of lignin-enzyme interactions: Recent advances in developing state-of-the-art analytical techniques. Biotechnol. Adv. 2022, 54, 107830. [Google Scholar] [CrossRef]
- Houfani, A.A.; Anders, N.; Spiess, A.N.; Baldrian, P.; Benallaoua, S. Insights from enzymatic degradation of cellulose and hemicellulose to fermentable sugars—A review. Biomass Bioenergy 2021, 134, 105481. [Google Scholar] [CrossRef]
- Lu, X.; Li, F.; Zhou, X.; Hu, J.; Liu, P. Biomass, lignocellulolytic enzyme production and lignocellulose degradation patterns by Auricularia auricula during solid state fermentation of corn stalk residues under different pretreatments. Food Chem. 2022, 384, 132622. [Google Scholar] [CrossRef]
- Saritpongteeraka, K.; Chaiprapat, S.; Boonsawang, P.; Sung, S. Solid state co-fermentation as pretreatment of lignocellulosic palm empty fruit bunch for organic acid recovery and fiber property improvement. Int. Biodeterior. Biodegrad. 2015, 100, 172–180. [Google Scholar] [CrossRef]
- Lin, C.A.; Cheng, C.; Chen, L.W.; Chen, C.W.; Duan, K.J. Ethanol production using the whole solid-state fermented sugarcane bagasse cultivated by Trichoderma reesei RUT-C30 supplemented with commercial cellulase. Biocatal. Agricult. Biotechnol. 2023, 50, 102667. [Google Scholar] [CrossRef]
- Rollin, J.A.; Zhu, Z.; Sathitsuksanoh, N.; Zhang, Y.-H.P. Increasing cellulose accessibility is more important than removing lignin: A comparison of cellulose solvent-based lignocellulose fractionation and soaking in aqueous ammonia. Biotechnol. Bioeng. 2011, 108, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, J.B.; Thygesen, L.G.; Felby, C.; Jørgensen, H.; Elder, T. Cell-wall structural changes in wheat straw pretreated for bioethanol production. Biotechnol. Biofuels 2008, 1, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Fan, C.; Hu, H.; Li, Y.; Sun, D.; Wang, Y.; Peng, L. Genetic modification of plant cell walls to enhance biomass yield and biofuel production in bioenergy crops. Biotechnol. Adv. 2016, 34, 997–1017. [Google Scholar] [CrossRef]
- El Hage, R.; Chrusciel, L.; Desharnais, L.; Brosse, N. Effect of autohydrolysis of Miscanthus giganteus on lignin structure and organosolv delignification. Bioresour. Technol. 2010, 101, 9321–9329. [Google Scholar] [CrossRef]
- Silverstein, R.A.; Chen, Y.; Sharma-Shivappa, R.R.; Boyette, M.D.; Osborne, J. A comparison of chemical pretreatment methods for improving saccharification of cotton stalks. Bioresour. Technol. 2007, 98, 3000–3011. [Google Scholar] [CrossRef]
- Liu, C.; Wyman, C.E. Partial flow of compressed-hot water through corn stover to enhance hemicellulose sugar recovery and enzymatic digestibility of cellulose. Bioresour. Technol. 2005, 96, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- de Martini, J.D.; Pattathil, S.; Avci, U.; Szekalski, K.; Mazumder, K.; Hahn, M.G.; Wyman, C.E. Application of monoclonal antibodies to investigate plant cell wall deconstruction for biofuels production. Energy Environ. Sci. 2011, 4, 4332–4339. [Google Scholar] [CrossRef]
- Huang, Y.; Wei, X.; Zhou, S.; Liu, M.; Tu, Y.; Li, A.; Chen, P.; Wang, Y.; Zhang, X.; Tai, H.; et al. Steam enzymatic saccharification of cotton stalks by largely reducing cellulose polymerization degree in G. barbadense and G. hirsutum. Bioresour. Technol. 2015, 181, 224–230. [Google Scholar] [CrossRef]
- Jin, W.; Chen, L.; Hu, M.; Sun, D.; Li, A.; Li, Y.; Hu, Z.; Zhou, S.; Tu, Y.; Xia, T.; et al. Tween-80 is effective for enhancing steam exploded biomass enzymatic saccharification and ethanol production by specifically lessening cellulose absorption with lignin in common reed. Appl. Energy 2016, 175, 82–90. [Google Scholar] [CrossRef]
- Pasko, R.Z.; Timm, T.G.; de Lima, G.G.; Helm, C.V.; de Lima, E.A.; Henriques, G.S.; Tavares, L.B.B. In Vivo evaluation and nutritional quality of by-products subjected to solid-state fermentation using shiitake culinary-medicinal mushroom, Lentinula edodes (Agaricomycetes). Int. J. Med. Mushrooms 2022, 24, 53–66. [Google Scholar] [CrossRef]
- Carvalho, E.A.; Nunes, L.V.; Goes, L.M.S.; Silva, E.G.P.; Franco, M.; Gross, E.; Uetanabaro, A.P.; Costa, A.M. Peach-palm (Bactris gasipaes Kunth.) waste as substrate for xylanase production by Trichoderma stromaticum AM7. Chem. Eng. Commun. 2018, 205, 975–985. [Google Scholar] [CrossRef]
- Bezerra, C.O.; Carneiro, L.L.; Carvalho, E.A.; Chagas, T.P.; de Carvalho, L.R.; Uetanabaro, A.P.T.; da Silva, G.P.; da Silva, E.G.P.; da Costa, A.M. Artificial intelligence as a combinatorial optimization strategy for cellulase production by Trichoderma stromaticum AM7 using peach-palm waste under solid-state fermentation. Bioenergy Res. 2021, 14, 1161–1170. [Google Scholar] [CrossRef]
- Chicatto, J.A.; Rainert, K.T.; Gonçalves, M.J.; Helm, C.V.; Altmajer-Vaz, D.; Tavares, L.B.B. Decolorization of textile industry wastewater in solid state fermentation with peach-palm (Bactris gasipaes) residue. Braz. J. Biol. 2018, 78, 718–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giombelli, C.; Iwassa, I.J.; da Silva, C.; Barros, B.C.B. Valorization of peach palm by-product through subcritical water extraction of soluble sugars and phenolic compounds. J. Supercrit. Fluids 2020, 165, 104985. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 35–83. [Google Scholar] [CrossRef]
- Vogel, H.J. A convenient growth medium for Neurospora (Medium N). Microb. Genet. Bull. 1956, 13, 42–43. [Google Scholar]
- Mota, T.R.; Kato, C.G.; Peralta, R.A.; Bracht, A.; Morais, G.R.; Baesso, M.L.; Souza, C.G.M.; Peralta, R.M. Decolourization of Congo red by Ganoderma lucidum laccase: Evaluation of degradation products and toxicity. Water Air Soil Poll. 2015, 226, 351. [Google Scholar] [CrossRef]
- Miller, G.L. Dinitrosalicylic acid reagent for determination of reducing sugars. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bergmeyer, H.U.; Bernt, E. Determination of glucose with glucose oxidase and peroxidase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: London, UK, 1974; pp. 1205–1215. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Van Soest Fiber Analysis | Peach Palm Inner Sheaths (g/100 g Material) | Peach Palm Peel (g/100 g Material) | ||
|---|---|---|---|---|
| Untreated | Pretreated | Untreated | Pretreated | |
| Cellulose | 35.0 ± 5 | 30 ± 4 | 32.0 ± 4 | 29.0 ± 4 |
| Hemicellulose | 20.0 ± 3 | 14 ± 3 | 23.0 ± 5 | 18.3 ± 3 |
| Acid detergent lignin | 24.0 ± 4 | 21 ± 4 | 20.0 ± 5 | 19.2 ± 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Cássia Spacki, K.; Novi, D.M.P.; de Oliveira-Junior, V.A.; Durigon, D.C.; Fraga, F.C.; dos Santos, L.F.O.; Helm, C.V.; de Lima, E.A.; Peralta, R.A.; de Fátima Peralta Muniz Moreira, R.; et al. Improving Enzymatic Saccharification of Peach Palm (Bactris gasipaes) Wastes via Biological Pretreatment with Pleurotus ostreatus. Plants 2023, 12, 2824. https://doi.org/10.3390/plants12152824
de Cássia Spacki K, Novi DMP, de Oliveira-Junior VA, Durigon DC, Fraga FC, dos Santos LFO, Helm CV, de Lima EA, Peralta RA, de Fátima Peralta Muniz Moreira R, et al. Improving Enzymatic Saccharification of Peach Palm (Bactris gasipaes) Wastes via Biological Pretreatment with Pleurotus ostreatus. Plants. 2023; 12(15):2824. https://doi.org/10.3390/plants12152824
Chicago/Turabian Stylede Cássia Spacki, Kamila, Danielly Maria Paixão Novi, Verci Alves de Oliveira-Junior, Daniele Cocco Durigon, Fernanda Cristina Fraga, Luís Felipe Oliva dos Santos, Cristiane Vieira Helm, Edson Alves de Lima, Rosely Aparecida Peralta, Regina de Fátima Peralta Muniz Moreira, and et al. 2023. "Improving Enzymatic Saccharification of Peach Palm (Bactris gasipaes) Wastes via Biological Pretreatment with Pleurotus ostreatus" Plants 12, no. 15: 2824. https://doi.org/10.3390/plants12152824