
Biotechnological Tools to Elucidate the Mechanism of Plant and Nematode Interactions



Abstract
:1. Introduction
2. Biology of Root-Knot Nematodes
2.1. Life Cycle of Root-Knot Nematodes
2.2. The Genomes of the Root-Knot Nematodes
2.3. Exploration of Available Resources for Research of Root-Knot Nematodes
3. Molecular Strategies
3.1. Transcriptomic Technology
3.2. Effector Molecules
4. Molecular Genetics Approaches
4.1. siRNA Technology
4.2. RNAi Technology
4.3. Quantitative Trait Loci (QTLs)
4.4. Genetic Engineering Strategies
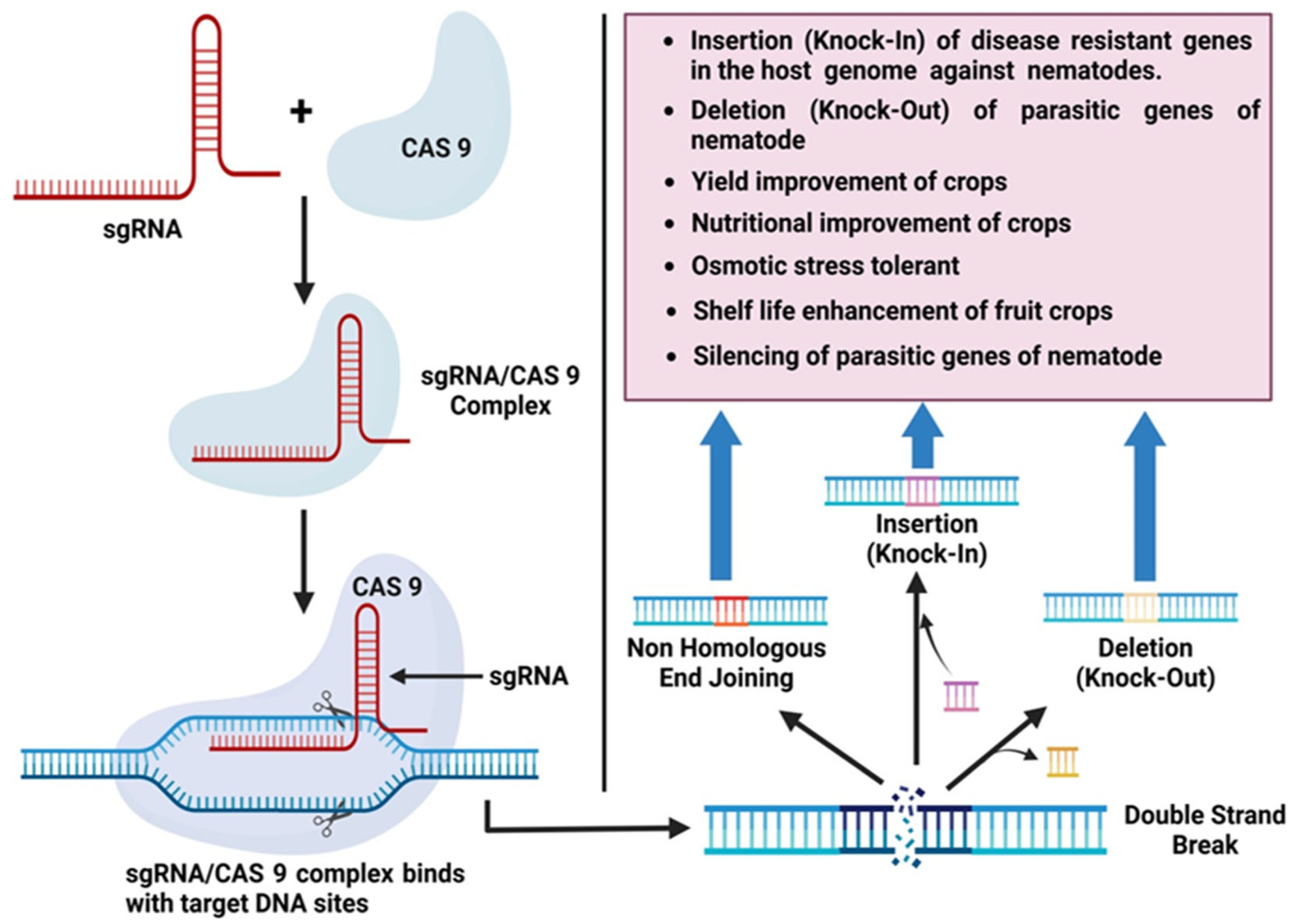
CRISPR/Cas9 Technology
5. Physiological Approaches
5.1. Metabolomics
5.2. Phytoalexins and Phytoanticipins
5.3. Volatile Organic Compounds (VOCs)
6. Conclusions
7. Future Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. State of Food Insecurity 2009; Food and Agriculture Organization: Rome, Italy, 2009. [Google Scholar]
- Mitreva, M.; Blaxter, M.L.; Bird, D.M.; McCarter, J.P. Comparative genomics of nematodes. Trends Genet. 2005, 21, 573–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, H.M.M.; Ahmad, E.M.; Martinez-Medina, A.; Aly, M.A.M. Effective approaches to study the plant-root knot nematode interaction. Plant Physiol. Biochem. 2019, 141, 332–342. [Google Scholar] [CrossRef]
- Nicol, J.M.; Turner, S.J.; Coyne, D.L.; Nijs, L.D.; Hockland, S.; Maafi, Z.T. Current nematode threats to world agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Springer: Dordrecht, The Netherlands, 2011; pp. 21–43. [Google Scholar] [CrossRef]
- Coyne, D.L.; Cortada, L.; Dalzell, J.J.; Claudius-Cole, A.O.; Haukeland, S.; Luambano, N.; Talwana, H. Plant-parasitic nematodes and food security in sub-Saharan Africa. Annu. Rev. Phytopathol. 2018, 56, 381–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decraemer, W.; Hunt, D. Plant nematology. In Structure and Classification; Perry, R.N., Moens, M., Eds.; CABI: Cambridge, MA, USA, 2006; pp. 3–32. [Google Scholar]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.; Gaur, H.S.; Helder, J.; Jones, M.G.; Perry, R.N. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.C.Z. Current nematode threats to Brazilian agriculture. Curr. Agric. Sci. Technol. 2014, 20, 26–35. [Google Scholar]
- Tapia-Vázquez, I.; Montoya-Martínez, A.C.; los Santos-Villalobos, D.; Ek-Ramos, M.J.; Montesinos-Matías, R.; Martínez-Anaya, C. Root-knot nematodes (Meloidogyne spp.) a threat to agriculture in Mexico: Biology, current control strategies, and perspectives. World J. Microbiol. Biotechnol. 2022, 38, 26. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Understanding molecular plant-nematode interactions to develop alternative approaches for nematode control. Plants 2022, 11, 2141. [Google Scholar] [CrossRef]
- Dutta, T.K.; Banakar, P.; Rao, U. The status of RNAi-based transgenic research in plant nematology. Front. Microbiol. 2015, 5, 760. [Google Scholar] [CrossRef]
- Gowda, M.; Rai, A.; Singh, B. Root Knot Nematode a Threat to Vegetable Production and Its Management; IIVR Technology: New York, NY, USA, 2017. [Google Scholar]
- Forghani, F.; Hajihassani, A. Recent advances in the development of environmentally benign treatments to control root-knot nematodes. Front. Plant Sci. 2020, 11, 1125. [Google Scholar] [CrossRef]
- Singh, P.R.; Karssen, G.; Couvreur, M.; Subbotin, S.A.; Bert, W. Integrative taxonomy and molecular phylogeny of the plant-parasitic nematode genus Paratylenchus (Nematoda: Paratylenchinae): Linking species with molecular barcodes. Plants 2021, 10, 408. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Ali, M.A.; Azeem, F.; Li, H.; Bohlmann, H. Smart parasitic nematodes use multifaceted strategies to parasitize plants. Front. Plant Sci. 2017, 8, 1699. [Google Scholar] [CrossRef] [Green Version]
- Sikandar, A.; Zhang, M.; Wang, Y.; Zhu, X.; Liu, X.; Fan, H.; Duan, Y. Nematodes a risk to agriculture. Appl. Ecol. Environ. Res. 2020, 18, 1679–1690. [Google Scholar] [CrossRef]
- Da Rocha, M.; Bournaud, C.; Dazeniere, J.; Thorpe, P.; Bailly-Bechet, M.; Pellegrin, C.; Danchin, E.G. Genome expression dynamics reveal the parasitism regulatory landscape of the root-knot nematode Meloidogyne incognita and a promoter motif associated with effector genes. Genes 2021, 12, 771. [Google Scholar] [CrossRef]
- Castagnone-Sereno, P.; Mulet, K.; Danchin, E.G.; Koutsovoulos, G.D.; Karaulic, M.; Da Rocha, M.; Abad, P. Gene copy number variations as signatures of adaptive evolution in the parthenogenetic, plant-parasitic nematode Meloidogyne incognita. Mol. Ecol. 2019, 28, 2559–2572. [Google Scholar] [CrossRef]
- Castagnone-Sereno, P.; Danchin, E.G.J. Parasitic success without sex—The nematode experience. J. Evol. Biol. 2014, 27, 1323–1333. [Google Scholar] [CrossRef]
- Baniya, A.; Joseph, S.; Duncan, L.; Crow, W.; Mengistu, T. The role of Caenorhabditis elegans sex-determination homologs, Misdc-1 and Mi-tra-1 in Meloidogyne incognita. Eur. J. Plant Pathol. 2021, 161, 439–452. [Google Scholar] [CrossRef]
- Eisenback, J.D.; Triantaphyllou, H.H. Root-knot nematodes: Meloidogyne species and races. In Manual of Agricultural Nematology; CRC Press: Boca Raton, FL, USA, 2020; pp. 191–274. [Google Scholar]
- Escobar, C.; Barcala, M.; Cabrera, J.; Fenoll, C. Overview of root-knot nematodes and giant cells. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2015; Volume 73, pp. 1–32. [Google Scholar]
- Perry, R.N.; Moens, M. Introduction to plant-parasitic nematodes modes of parasitism. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Springer: Dordrecht, The Netherlands, 2011; pp. 3–20. [Google Scholar] [CrossRef]
- Vieira, P.; Danchin, E.G.; Neveu, C.; Crozat, C.; Jaubert, S.; Hussey, R.S.; Rosso, M.N. The plant apoplasm is an important recipient compartment for nematode secreted proteins. J. Exp. Bot. 2011, 62, 1241–1253. [Google Scholar] [CrossRef] [Green Version]
- Chitwood, D.J.; Lusby, W.R.; Thompson, M.J.; Kochansky, J.P.; Howarth, O.W. The glycosylceramides of the nematode Caenorhabditis elegans contain an unusual, branched-chain sphingoid base. Lipids 1995, 30, 567–573. [Google Scholar] [CrossRef]
- Chitwood, D.J. Research on plant-parasitic nematode biology conducted by the United States Department of Agriculture–Agricultural Research Service. Pest Manag. Sci. 2003, 59, 748–753. [Google Scholar] [CrossRef]
- Curtis, R.H.; Robinson, A.F.; Perry, R.N. Hatch and host location. In Root-Knot Nematodes; CABI: Wallingford, UK, 2009; pp. 139–162. [Google Scholar]
- Bird, D.M.; Opperman, C.H. Caenorhabditis elegans: A genetic guide to parasitic nematode biology. J. Nematol. 1998, 30, 299–308. [Google Scholar]
- Riddle, D.L.; Albert, P.S. Genetic and environmental regulation of dauer larva development. In Caenorhabditis Elegans, 2nd ed.; Riddle, D.L., Blumenthal, T., Meyer, J., Preiss, J.R., Eds.; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1997; pp. 739–768. [Google Scholar]
- Ramsay, K.; Wang, Z.; Jones, M.G.K. Using laser capture microdissection to study gene expression in early stages of giant cells induced by root-knot nematodes. Mol. Plant Pathol. 2004, 5, 587–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Magill, C.; Starr, J.L. Laser capture microdissection and real-time PCR for measuring mRNA in giant cells induced by Meloidogyne javanica. J. Nematol. 2005, 37, 308. [Google Scholar] [PubMed]
- Iqbal, S.; Fosu-Nyarko, J.; Jones, M.G. Attempt to silence genes of the RNAi pathways of the root-knot nematode, Meloidogyne incognita results in diverse responses including increase and no change in expression of some genes. Front. Plant Sci. 2020, 11, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, N.; Yadav, R.; Kaur, P.; Rasmussen, S.; Goel, S.; Agarwal, M.; Kumar, A. Transcriptome analysis of root-knot nematode (Meloidogyne incognita)-infected tomato (Solanum lycopersicum) roots reveals complex gene expression profiles and metabolic networks of both host and nematode during susceptible and resistance responses. Mol. Plant Pathol. 2018, 19, 615–633. [Google Scholar] [CrossRef] [Green Version]
- McCarter, J.P.; Dautova Mitreva, M.; Martin, J.; Dante, M.; Wylie, T.; Rao, U.; Waterston, R.H. Analysis and functional classification of transcripts from the nematode Meloidogyne incognita. Genome Biol. 2003, 4, R26. [Google Scholar] [CrossRef] [Green Version]
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Vuong, T.D.; Qiu, D.; Robbins, R.T.; Grover Shannon, J.; Li, Z.; Nguyen, H.T. Advancements in breeding, genetics, and genomics for resistance to three nematode species in soybean. Theor. Appl. Genet. 2016, 129, 2295–2311. [Google Scholar] [CrossRef]
- Bird, D.M.; Williamson, V.M.; Abad, P.; McCarter, J.; Danchin, E.G.; Castagnone-Sereno, P.; Opperman, C.H. The genomes of root-knot nematodes. Ann. Rev. Phytopathol. 2009, 47, 333–351. [Google Scholar] [CrossRef]
- Blanc-Mathieu, R.; Perfus-Barbeoch, L.; Aury, J.M.; Da Rocha, M.; Gouzy, J.; Sallet, E.; Danchin, E.G. Hybridization and polyploidy enable genomic plasticity without sex in the most devastating plant-parasitic nematodes. PLoS Genet. 2017, 13, e1006777. [Google Scholar] [CrossRef] [Green Version]
- Phan, N.T.; Orjuela, J.; Danchin, E.G.; Klopp, C.; Perfus-Barbeoch, L.; Kozlowski, D.K.; Bellafiore, S. Genome structure and content of the rice root-knot nematode (Meloidogyne graminicola). Ecol. Evol. 2020, 10, 11006–11021. [Google Scholar] [CrossRef]
- Susic, N.; Koutsovoulos, G.D.; Riccio, C.; Danchin, E.G.; Blaxter, M.L.; Lunt, D.H.; Stare, B.G. Genome sequence of the root-knot nematode. J. Nematol. 2020, 52, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Koutsovoulos, G.D.; Poullet, M.; Elashry, A.; Kozlowski, D.K.; Sallet, E.; Da Rocha, M.; Danchin, E.G. Genome assembly and annotation of Meloidogyne enterolobii, an emerging parthenogenetic root-knot nematode. Sci. Data 2020, 7, 324. [Google Scholar] [CrossRef]
- Lunt, D.H.; Kumar, S.; Koutsovoulos, G.; Blaxter, M.L. The complex hybrid origins of the root knot nematodes revealed through comparative genomics. Peer J. 2014, 2, e356. [Google Scholar] [CrossRef] [Green Version]
- Szitenberg, A.; Salazar-Jaramillo, L.; Blok, V.C.; Laetsch, D.R.; Joseph, S.; Williamson, V.M.; Lunt, D.H. Comparative genomics of apomictic root-knot nematodes: Hybridization, ploidy, and dynamic genome change. Genome Biol. Evol. 2017, 9, 2844–2861. [Google Scholar] [CrossRef] [Green Version]
- Opperman, C.H.; Bird, D.M.; Williamson, V.M.; Rokhsar, D.S.; Burke, M.; Cohn, J.; Cromer, J.; Diener, S.; Gajan, J.; Graham, S.; et al. Sequence and genetic map of Meloidogyne hapla: A compact nematode genome for plant parasitism. Proc. Natl. Acad. Sci. USA 2008, 105, 14802–14807. [Google Scholar] [CrossRef] [Green Version]
- Somvanshi, V.S.; Tathode, M.; Shukla, R.N.; Rao, U. Nematode Genome Announcement: A draft genome for rice root-knot nematode. J. Nematol. 2018, 50, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Harris, T.W.; Antoshechkin, I.; Bieri, T.; Blasiar, D.; Chan, J.; Chen, W.J.; De La Cruz, N.; Davis, P.; Duesbury, M.; Fang, R.; et al. WormBase: A comprehensive resource for nematode research. Nucleic Acids Res. 2010, 38, D463–D467. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; Rosa, B.A.; Ozersky, P.; Hallsworth-Pepin, K.; Zhang, X.; Bhonagiri-Palsikar, V.; Tyagi, R.; Wang, Q.; Choi, Y.J.; Gao, X.; et al. Helminth.net: Expansions to Nematode.net and an introduction to Trematode.net. Nucleic Acids Res. 2015, 43, D698–D706. [Google Scholar] [CrossRef] [Green Version]
- Geng, C.; Nie, X.; Tang, Z.; Zhang, Y.; Lin, J.; Sun, M.; Peng, D. A novel serine protease, Sep1, from Bacillus firmus DS-1 has nematicidal activity and degrades multiple intestinal-associated nematode proteins. Sci. Rep. 2016, 6, 25012. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, P.; Verma, A.; Verma, S.; Anwar, M.S.; Prasher, P.; Mudila, H.; Chen, S. Understanding phytomicrobiome: A potential reservoir for better crop management. Sustainability. 2020, 12, 5446. [Google Scholar] [CrossRef]
- Tsibidis, G.D.; Tavernarakis, N. Nemo: A computational tool for analyzing nematode locomotion. BMC Neurosci. 2007, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, J.; Mitreva, M. Transcriptomes of plant-parasitic nematodes. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Springer: Dordrecht, The Netherlands, 2011; pp. 119–138. [Google Scholar]
- Park, C.; Peng, Y.; Chen, X.; Dardick, C.; Ruan, D.; Bart, R.; Canlas, P.E.; Ronald, P.C. Rice XB15, a protein phosphatase 2C, negatively regulates cell death and XA21-mediated innate immunity. PLoS Biol. 2008, 6, e231. [Google Scholar]
- Beneventi, M.A.; da Silva, O.B.; de Sa, M.E.L.; Firmino, A.A.P.; de Amorim, R.M.S.; Albuquerque, E.V.S.; Grossi de-Sa, M.F. Transcription profile of soybean-root-knot nematode interaction reveals a key role of phythormones in the resistance reaction. BMC Genom. 2013, 14, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimaraes, P.M.; Guimaraes, L.A.; Morgante, C.V.; Silva, O.B., Jr.; Araujo, A.C.G.; Martins, A.C.; Brasileiro, A.C.M. Root transcriptome analysis of wild peanut reveals candidate genes for nematode resistance. PLoS ONE 2015, 10, e0140937. [Google Scholar] [CrossRef] [PubMed]
- Petitot, A.S.; Kyndt, T.; Haidar, R.; Dereeper, A.; Collin, M.; de Almeida Engler, J.; Fernandez, D. Transcriptomic and histological responses of African rice (Oryza glaberrima) to Meloidogyne graminicola provide new insights into root-knot nematode resistance in monocots. Ann. Bot. 2017, 119, 885–899. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Vuong, T.; Jiao, Y.; Joshi, T.; Zhang, H.; Nguyen, H.T. Whole-genome gene expression profiling revealed genes and pathways potentially involved in regulating interactions of soybean with cyst nematode (Heterodera glycines Ichinohe). BMC Genom. 2015, 16, 148. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Lin, Z.; Pang, S.; Zhang, W.; Bhatt, P.; Chen, S. Recent advanced technologies for the characterization of xenobiotic-degrading microorganisms and microbial communities. Front. Bioeng. Biotechnol. 2021, 9, 632059. [Google Scholar] [CrossRef]
- Samac, D.A.; Penuela, S.; Schnurr, J.A.; Hunt, E.N.; Foster-Hartnett, D.; Van-denbosch, K.A.; Gantt, J.S. Expression of coordinately regulated defence response genes and analysis of their role in disease resistance in Medicago truncatula. Mol. Plant Pathol. 2011, 12, 786–798. [Google Scholar] [CrossRef]
- Barilli, E.; Rubiales, D.; Gjetting, T.; Lyngkjaer, M.F. Differential gene transcript accumulation in peas in response to powdery mildew (Erysiphe pisi) attack. Euphytica 2014, 198, 13–28. [Google Scholar] [CrossRef]
- Hekman, J.P.; Johnson, J.L.; Kukekova, A.V. Transcriptome analysis in domesticated species: Challenges and strategies. Bioinform. Biol. Insights 2015, 9, 21–31. [Google Scholar] [CrossRef]
- Abad, P.; Williamson, V.M. Plant nematode interaction: A sophisticated dialogue. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2010; Volume 53, pp. 147–192. [Google Scholar]
- Schaff, J.E.; Nielsen, D.M.; Smith, C.P.; Scholl, E.H.; Bird, D.M. Comprehensive transcriptome profiling in tomato reveals a role for glycosyltransferase in Mi-mediated nematode resistance. Plant Physiol. 2007, 144, 1079–1092. [Google Scholar] [CrossRef] [Green Version]
- Portillo, M.E.; Corvec, S.; Borens, O.; Trampuz, A. Propionibacterium acnes: An underestimated pathogen in implant-associated infections. BioMed Res. Int. 2013, 2013, 804391. [Google Scholar] [CrossRef] [Green Version]
- Hammes, U.Z.; Schachtman, D.P.; Berg, R.H.; Nielsen, E.; Koch, W.; McIntyre, L.M.; Taylor, C.G. Nematode-induced changes of transporter gene expression in Arabidopsis roots. Mol. Plant Microbe Interact. 2005, 18, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- Fuller, V.L.; Lilley, C.J.; Atkinson, H.J.; Urwin, P.E. Differential gene expression in Arabidopsis following infection by plant-parasitic nematodes Meloidogyne incognita and Heterodera schachtii. Mol. Plant Pathol. 2007, 8, 595–609. [Google Scholar] [CrossRef]
- Barcala, M.; García, A.; Cabrera, J.; Casson, S.; Lindsey, K.; Favery, B.; Escobar, C. Early transcriptomic events in microdissected Arabidopsis nematode-induced giant cells. Plant J. 2010, 61, 698–712. [Google Scholar] [CrossRef]
- Bagnaresi, P.; Sala, T.; Irdani, T.; Scotto, C.; Lamontanara, A.; Beretta, M.; Sabatini, E. Solanum torvum responses to the root-knot nematode Meloidogyne incognita. BMC Genom. 2013, 14, 540. [Google Scholar] [CrossRef] [Green Version]
- Jammes, F.; Lecomte, P.; de Almeida-Engler, J.; Bitton, F.; Martin-Magniette, M.L.; Renou, J.P.; Favery, B. Genome-wide expression profiling of the host response to root-knot nematode infection in Arabidopsis. Plant J. 2005, 44, 447–458. [Google Scholar] [CrossRef]
- Caillaud, M.C.; Dubreuil, G.; Quentin, M.; Perfus-Barbeoch, L.; Lecomte, P.; de Almeida Engler, J. Root-knot nematodes manipulate plant cell functions during a compatible interaction. Plant Physiol. 2008, 165, 104–113. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, D.; Shuang, L.; Xiao, D.; Xuan, Y.; Duan, Y.; Zhu, X. Transcriptome analysis of rice roots in response to root-knot nematode infection. Int. J. Mol. Sci. 2020, 21, 848. [Google Scholar] [CrossRef] [Green Version]
- Eves-van den Akker, S.; Stojilkovic, B.; Gheysen, G. Recent applications of biotechnological approaches to elucidate the biology of plant–nematode interactions. Curr. Opin. Biotechnol. 2021, 70, 122–130. [Google Scholar] [CrossRef]
- Niu, J.; Liu, P.; Liu, Q.; Chen, C.; Guo, Q.; Yin, J.; Jian, H. Msp40 effector of root-knot nematode manipulates plant immunity to facilitate parasitism. Sci. Rep. 2016, 6, 19443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, E.L.; Hussey, R.S.; Mitchum, M.G.; Baum, T.J. Parasitism proteins in nematode–plant interactions. Curr. Opin. Plant Biol. 2008, 11, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Mitchum, M.G.; Hussey, R.S.; Baum, T.J.; Wang, X.; Elling, A.A.; Wubben, M.; Davis, E.L. Nematode effector proteins: An emerging paradigm of parasitism. New Phytol. 2013, 199, 879–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haegeman, A.; Mantelin, S.; Jones, J.T.; Gheysen, G. Functional roles of effectors of plant-parasitic nematodes. Gene 2012, 492, 19–31. [Google Scholar] [CrossRef]
- Ali, M.A.; Abbas, A.; Azeem, F.; Javed, N.; Bohlmann, H. Plant-nematode interactions: From genomics to metabolomics. Int. J. Agric Biol. 2015, 17, 1071–1082. [Google Scholar]
- Gheysen, G.; Fenoll, C. Gene expression in nematode feeding sites. Ann. Rev. Phytopathol. 2002, 40, 191–219. [Google Scholar] [CrossRef]
- Gao, B.; Allen, R.; Maier, T.; Davis, E.L.; Baum, T.J.; Hussey, R.S. Identification of a new beta-1,4-endoglucanase gene expressed in the esophageal subventral gland cells of Heterodera glycines. J. Nematol. 2002, 34, 12–15. [Google Scholar]
- Qin, L.; Kudla, U.; Roze, E.H.; Goverse, A.; Popeijus, H.; Nieuwland, J.; Helder, J. A nematode expansin acting on plants. Nature 2004, 427, 30. [Google Scholar] [CrossRef]
- Kudla, U.; Qin, L.; Milac, A.; Kielak, A.; Maissen, C.; Overmars, H.; Popeijus, H.; Roze, E.; Petrescu, A.; Smant, G.; et al. Origin, distribution and 3D-modeling of Gr-EXPB1, an expansin from the potato cyst nematode Globodera rostochiensis. FEBS Lett. 2005, 579, 2451–2457. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Allen, R.; Davis, E.L.; Baum, T.J.; Hussey, R.S. Molecular characterization and developmental expression of a cellulose-binding protein gene in the soybean cyst nematode Heterodera glycines. Int. J. Parasitol. 2004, 34, 1377–1383. [Google Scholar] [CrossRef]
- Truong, N.M.; Nguyen, C.N.; Abad, P.; Quentin, M.; Favery, B. Function of root-knot nematode effectors and their targets in plant parasitism. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2015; Volume 73, pp. 293–324. [Google Scholar]
- Jaouannet, M.; Perfus-Barbeoch, L.; Deleury, E.; Magliano, M.; Engler, G.; Vieira, P.; Rosso, M.N. A root-knot nematode-secreted protein is injected into giant cells and targeted to the nuclei. New Phytol. 2012, 194, 924–931. [Google Scholar] [CrossRef]
- Lin, J.; Mazarei, M.; Zhao, N.; Zhu, J.J.; Zhuang, X.; Liu, W.; Chen, F. Overexpression of a soybean salicylic acid methyltransferase gene confers resistance to soybean cyst nematode. Plant Biotechnol. J. 2013, 11, 1135–1145. [Google Scholar] [CrossRef]
- Rivas, S.; Genin, S. A plethora of virulence strategies hidden behind nuclear targeting of microbial effectors. Front. Plant Sci. 2011, 2, 104. [Google Scholar] [CrossRef] [Green Version]
- Deslandes, L.; Rivas, S. The plant cell nucleus: A true arena for the fight between plants and pathogens. Plant Signal. Behav. 2012, 6, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Leelarasamee, N.; Zhang, L.; Gleason, C. The root-knot nematode effector MiPFN3 disrupts plant actin filaments and promotes parasitism. PLoS Pathog. 2018, 14, e1006947. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.N.; Perfus-Barbeoch, L.; Quentin, M.; Zhao, J.; Magliano, M.; Marteu, N.; Favery, B. A root-knot nematode small glycine and cysteine-rich secreted effector, MiSGCR1, is involved in plant parasitism. New Phytol. 2018, 217, 687–699. [Google Scholar] [CrossRef] [Green Version]
- Mejias, J.; Bazin, J.; Truong, N.M.; Chen, Y.; Marteu, N.; Bouteiller, N.; Quentin, M. The root-knot nematode effector MiEFF18 interacts with the plant core spliceosomal protein SmD1 required for giant cell formation. New Phytol. 2021, 229, 3408–3423. [Google Scholar] [CrossRef]
- Chen, J.; Hu, L.; Sun, L.; Lin, B.; Huang, K.; Zhuo, K.; Liao, J. A novel Meloidogyne graminicola effector, MgMO237, interacts with multiple host defence-related proteins to manipulate plant basal immunity and promote parasitism. Mol. Plant Pathol. 2018, 19, 1942–1955. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.; Chang, H.Y.; Daugherty, L.; Fraser, M.; Hunter, S.; Lopez, R.; McAnulla, C.; McMenamin, C.; Nuka, G.; Pesseat, S.; et al. The InterPro protein families database: The classification resource after 15 years. Nucleic Acids Res. 2015, 43, D213–D221. [Google Scholar] [CrossRef]
- Kudla, U.; Milac, A.L.; Qin, L.; Overmars, H.; Roze, E.; Holterman, M.; Smant, G. Structural and functional characterization of a novel, host penetration-related pectate lyase from the potato cyst nematode Globodera rostochiensis. Mol. Plant Pathol. 2007, 8, 293–305. [Google Scholar] [CrossRef]
- Li, X.; Zhuo, K.; Luo, M.; Sun, L.; Liao, J. Molecular cloning and characterization of a calreticulin cDNA from the pinewood nematode Bursaphelenchus xylophilus. Exp. Parasitol. 2011, 128, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Jaouannet, M.; Magliano, M.; Arguel, M.J.; Gourgues, M.; Evangelisti, E.; Abad, P.; Rosso, M.N. The root-knot nematode calreticulin Mi-CRT is a key effector in plant defense suppression. Mol. Plant-Microbe Interact. 2013, 26, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iberkleid, I.; Vieira, P.; de Almeida-Engler, J.; Firester, K.; Spiegel, Y.; Horowitz, S.B. Fatty acid-and retinol-binding protein, Mj-FAR-1 induces tomato host susceptibility to root-knot nematodes. PLoS ONE 2013, 8, e64586. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Cui, R.; Sun, L.; Lin, B.; Zhuo, K.; Liao, J. Molecular and biochemical characterization of the b-1,4-endoglucanase gene Mj-eng-3 in the root-knot nematode Meloidogyne javanica. Exp. Parasitol. 2013, 135, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Zhuo, K.; Chen, S.; Hu, L.; Sun, L.; Wang, X.; Liao, J. A novel nematode effector suppresses plant immunity by activating host reactive oxygen species-scavenging system. New Phytol. 2016, 209, 1159–1173. [Google Scholar] [CrossRef]
- Li, J.; Han, S.; Ding, X.; He, T.; Dai, J.; Yang, S.; Gai, J. Comparative transcriptome analysis between the cytoplasmic male sterile line NJCMS1A and its maintainer NJCMS1B in soybean (Glycine max (L.) Merr.). PLoS ONE 2015, 10, e0126771. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Li, S.; Mo, C.; Wang, G.; Xiao, X.; Xiao, Y. A novel Meloidogyne incognita effector Misp12 suppresses plant defense response at latter stages of nematode parasitism. Front. Plant Sci. 2016, 7, 964. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Peng, H.; Cui, J.; Huang, W.; Kong, L.; Clarke, J.L.; Peng, D. Molecular characterization of a novel effector expansin-like protein from Heterodera avenae that induces cell death in Nicotiana benthamiana. Sci. Rep. 2016, 6, 35677. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, K.; Chen, J.; Lin, B.; Wang, J.; Sun, F.; Hu, L.; Liao, J. A novel Meloidogyne enterolobii effector MeTCTP promotes parasitism by suppressing programmed cell death in host plants. Mol. Plant Pathol. 2017, 18, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lin, B.; Huang, Q.; Hu, L.; Zhuo, K.; Liao, J. A novel Meloidogyne graminicola effector, MgGPP, is secreted into host cells and undergoes glycosylation in concert with proteolysis to suppress plant defenses and promote parasitism. PLoS Pathog. 2017, 13, e1006301. [Google Scholar] [CrossRef] [Green Version]
- Naalden, D.; Haegeman, A.; de Almeida-Engler, J.; Birhane Eshetu, F.; Bauters, L.; Gheysen, G. The Meloidogyne graminicola effector Mg16820 is secreted in the apoplast and cytoplasm to suppress plant host defense responses. Mol. Plant Pathol. 2018, 19, 2416–2430. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Mao, Z.; Zhang, X.; Ling, J.; Lin, R.; Zhang, X.; Xie, B. The novel secreted Meloidogyne incognita effector MiISE6 targets the host nucleus and facilitates parasitism in Arabidopsis. Front. Plant Sci. 2018, 9, 252. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Mejias, J.; Quentin, M.; Chen, Y.; de Almeida-Engler, J.; Mao, Z.; Jian, H. The root-knot nematode effector MiPDI1 targets a stress-associated protein (SAP) to establish disease in Solanaceae and Arabidopsis. New Phytol. 2020, 228, 1417–1430. [Google Scholar] [CrossRef]
- Truong, N.M.; Chen, Y.; Mejias, J.; Soulé, S.; Mulet, K.; Jaouannet, M.; Quentin, M. The Meloidogyne incognita nuclear effector MiEFF1 interacts with Arabidopsis cytosolic glyceraldehyde-3-phosphate dehydrogenases to promote parasitism. Front. Plant Sci. 2021, 12, 641480. [Google Scholar] [CrossRef]
- Qin, X.; Xue, B.; Tian, H.; Fang, C.; Yu, J.; Chen, C.; Wang, X. An unconventionally secreted effector from the root knot nematode Meloidogyne incognita, Mi-ISC-1, promotes parasitism by disrupting salicylic acid biosynthesis in host plants. Mol. Plant Pathol. 2022, 23, 516–529. [Google Scholar] [CrossRef]
- Nobori, T.; Tsuda, K. The plant immune system in heterogeneous environments. Curr. Opin. Plant Biol. 2019, 50, 58–66. [Google Scholar] [CrossRef]
- Bakhetia, M.; Charlton, W.L.; Urwin, P.E.; McPherson, M.J.; Atkinson, H.J. RNA interference and plant parasitic nematodes. Trends Plant Sci. 2005, 10, 362–367. [Google Scholar] [CrossRef]
- Sen, G.L.; Blau, H.M. A brief history of RNAi: The silence of the genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef] [Green Version]
- Runo, S.; Alakonya, A.; Machuka, J.; Sinha, N. RNA interference as a resistance mechanism against crop parasites in Africa: A ‘Trojan horse’ approach. Pest Manag. Sci. 2011, 67, 129–136. [Google Scholar] [CrossRef]
- Lilley, C.J.; Davies, L.J.; Urwin, P.E. RNA interference in plant parasitic nematodes: A summary of the current status. Parasitol. 2012, 139, 630–640. [Google Scholar] [CrossRef]
- Yadav, B.C.; Veluthambi, K.; Subramaniam, K. Host-generated double stranded RNA induces RNAi in plant-parasitic nematodes and protects the host from infection. Mol. Biochem. Parasit. 2006, 148, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.Z.; Allen, R.; Davis, E.L.; Baum, T.J.; Hussey, R.S. Engineering broad root-knot resistance in transgenic plants by RNAi silencing of a conserved and essential root-knot nematode parasitism gene. Proc. Natl. Acad. Sci. USA 2006, 103, 14302–14306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlton, W.L.; Harel, H.Y.M.; Bakhetia, M.; Hibbard, J.K.; Atkinson, H.J.; McPherson, M.J. Additive effects of plant expressed double-stranded RNAs on root-knot nematode development. Int. J. Parasitol. 2010, 40, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.M.; Alkharouf, N.W.; Meyer, S.L.; Aly, M.A.; El-Din, A.E.K.Y.G.; Hussein, E.H.; Matthews, B.F. Post-transcriptional gene silencing of root-knot nematode in transformed soybean roots. Exp. Parasitol. 2011, 127, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Joshi, I.; Kohli, D.; Pal, A.; Chaudhury, A.; Sirohi, A.; Jain, P.K. Host delivered-RNAi of effector genes for imparting resistance against root-knot and cyst nematodes in plants. Physiol. Mol. Plant Pathol. 2022, 118, 101802. [Google Scholar] [CrossRef]
- Urwin, P.E.; Lilley, C.J.; Atkinson, H.J. Ingestion of double-stranded RNA by preparasitic juvenile cyst nematodes leads to RNA interference. Mol. Plant Microbe Interact. 2002, 15, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Rosso, M.N.; Dubrana, M.P.; Cimbolini, N.; Jaubert, S.; Abad, P. Application of RNA interference to root-knot nematode genes encoding esophageal gland proteins. Mol. Plant Microbe Interact. 2005, 18, 615–620. [Google Scholar] [CrossRef] [Green Version]
- Steeves, R.M.; Todd, T.C.; Essig, J.S.; Trick, H.N. Transgenic soybeans expressing siRNAs specific to a major sperm protein gene suppress Heterodera glycines reproduction. Funct. Biol. 2006, 33, 991–999. [Google Scholar] [CrossRef]
- Kumar, A.; Kakrana, A.; Sirohi, A.; Subramaniam, K.; Srinivasan, R.; Abdin, M.Z.; Jain, P.K. Host-delivered RNAi-mediated root-knot nematode resistance in Arabidopsis by targeting splicing factor and integrase genes. J. Gen. Plant Pathol. 2017, 83, 91–97. [Google Scholar] [CrossRef]
- Dinh, P.T.Y.; Brown, C.R.; Elling, A.A. RNA Interference of effector gene Mc16D10L confers resistance against Meloidogyne chitwoodi in Arabidopsis and Potato. Phytopathology 2014, 104, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Kakrana, A.; Kumar, A.; Satheesh, V.; Abdin, M.Z.; Subramaniam, K.; Bhattacharya, R.C.; Jain, P.K. Identification, validation and utilization of novel nematode-responsive root-specific promoters in Arabidopsis for inducing host-delivered RNAi mediated root-knot nematode resistance. Front. Plant Sci. 2017, 8, 2049. [Google Scholar] [CrossRef] [Green Version]
- Tamilarasan, S.; Rajam, M.V. Engineering crop plants for nematode resistance through host-derived RNA interference. Cell Dev. Biol. 2013, 2, 114. [Google Scholar] [CrossRef]
- Chaudhary, S.; Dutta, T.K.; Tyagi, N.; Shivakumara, T.N.; Papolu, P.K.; Chobhe, K.A.; Rao, U. Host-induced silencing of Mi-msp-1 confers resistance to root-knot nematode Meloidogyne incognita in eggplant. Transg. Res. 2019, 28, 327–340. [Google Scholar] [CrossRef]
- Chan, Y.L.; He, Y.; Hsiao, T.T.; Wang, C.J.; Tian, Z.; Yeh, K.W. Pyramiding taro cystatin and fungal chitinase genes driven by a synthetic promoter enhances resistance in tomato to root-knot nematode Meloidogyne incognita. Plant Sci. 2015, 231, 74–81. [Google Scholar] [CrossRef]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Mundt, C.C. Use of multiline cultivars and cultivar mixtures for disease management. Annu. Rev. Phytopathol. 2002, 40, 381–410. [Google Scholar] [CrossRef] [Green Version]
- Pink, D. Strategies using genes for non-durable disease resistance. Euphytica 2002, 124, 227–236. [Google Scholar] [CrossRef]
- Nissan, N.; Mimee, B.; Cober, E.R.; Golshani, A.; Smith, M.; Samanfar, B. A broad review of soybean research on the ongoing race to overcome soybean cyst nematode. Biology 2022, 11, 211. [Google Scholar] [CrossRef]
- Ballen-Taborda, C.; Chu, Y.; Ozias-Akins, P.; Timper, P.; Holbrook, C.C.; Jackson, S.A.; Leal-Bertioli, S.C. A new source of root-knot nematode resistance from Arachis stenosperma incorporated into allotetraploid peanut (Arachis hypogaea). Sci. Rep. 2019, 9, 17702. [Google Scholar] [CrossRef] [Green Version]
- Fourie, H.; Mienie, C.M.; Mc Donald, A.H.; De Waele, D. Identification and validation of genetic markers associated with Meloidogyne incognita race 2 resistance in soybean, Glycine max (L.) Merr. Nematology 2008, 10, 651–661. [Google Scholar] [CrossRef]
- Shearin, Z.P.; Finnerty, S.L.; Wood, E.D.; Hussey, R.S.; Boerma, H.R. A Southern root-knot nematode resistance QTL linked to the T-Locus in Soybean. Crop Sci. 2009, 49, 467–472. [Google Scholar] [CrossRef]
- Liu, L. Genetic Mapping and Quantitative Trait Locus (QTL) Analysis of Root-Knot Nematode Resistance in Pearl Millet. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2012. [Google Scholar]
- Zwart, R.S.; Thompson, J.P.; Milgate, A.W.; Bansal, U.K.; Williamson, P.M.; Raman, H.; Bariana, H.S. QTL mapping of multiple foliar disease and root-lesion nematode resistances in wheat. Mol. Breed. 2010, 26, 107–124. [Google Scholar] [CrossRef]
- Galeng-Lawilao, J.; Kumar, A.; De Waele, D. QTL mapping for resistance to and tolerance for the rice root-knot nematode, Meloidogyne graminicola. BMC Genet. 2018, 19, 53. [Google Scholar] [CrossRef] [Green Version]
- Khanal, C.; McGawley, E.C.; Overstreet, C.; Stetina, S.R. The elusive search for reniform nematode resistance in cotton. Phytopathology 2018, 108, 532–541. [Google Scholar] [CrossRef] [Green Version]
- Vander Vossen, E.; Sikkema, A.; Hekkert, B.T.L.; Gros, J.; Stevens, P.; Muskens, M.; Allefs, S. An ancient R gene from the wild potato species Solanum bulbocastanum confers broad-spectrum resistance to Phytophthora infestans in cultivated potato and tomato. Plant J. 2003, 36, 867–882. [Google Scholar] [CrossRef]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.; Embden, J.D.V.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microboil. 2002, 43, 1565–1575. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, Y.; Chi, Y.; Fan, B.; Chen, Z. Characterization of soybean WRKY gene family and identification of soybean WRKY genes that promote resistance to soybean cyst nematode. Sci. Rep. 2017, 7, 17804. [Google Scholar] [CrossRef] [Green Version]
- Ansari, W.A.; Chandanshive, S.U.; Bhatt, V.; Nadaf, A.B.; Vats, S.; Katara, J.L.; Deshmukh, R. Genome editing in cereals: Approaches, applications and challenges. Int. J. Mol. Sci. 2020, 21, 4040. [Google Scholar] [CrossRef]
- Jinek, M.; Jiang, F.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.; Anders, C.; Hauer, M.; Zhou, K.; Lin, S.; et al. Structures of Cas9 endonucleases reveal RNA-mediated conformational activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef] [Green Version]
- Puchta, H. Applying CRISPR/Cas for genome engineering in plants: The best is yet to come. Curr. Opin. Plant Biol. 2017, 36, 1–8. [Google Scholar] [CrossRef]
- Meng, X.; Yu, H.; Zhang, Y.; Zhuang, F.; Song, X.; Gao, S.; Li, J. Construction of a genome-wide mutant library in rice using CRISPR/Cas9. Mol. Plant 2017, 10, 1238–1241. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Mutti, J.; Young, J.K.; Yang, M.; Schroder, M.; Lenderts, B.; Wang, L.; Peterson, D.; St. Clair, G.; Jones, S.; et al. Complex Trait Loci in Maize Enabled by CRISPR-Cas9 Mediated Gene Insertion. Front. Plant Sci. 2020, 11, 535. [Google Scholar] [CrossRef]
- Zheng, N.; Li, T.; Dittman, J.D.; Su, J.; Li, R.; Gassmann, W.; Peng, D.; Whitham, S.A.; Liu, S.; Yang, B. CRISPR/Cas9-Based Gene Editing Using Egg Cell-Specific Promoters in Arabidopsis and Soybean. Front. Plant Sci. 2020, 11, 800. [Google Scholar] [CrossRef]
- Zinov’eva, S.V.; Vasyukova, N.I.; Ozeretskovskaya, O.L. Biochemical aspects of plant interactions with phytoparasitic nematodes: A review. Appl. Biochem. Microbiol. 2004, 40, 111–119. [Google Scholar] [CrossRef]
- Hofmann, J.; El Ashry, A.E.N.; Anwar, S.; Erban, A.; Kopka, J.; Grundler, F. Profiling reveals local and systemic responses of host plants to nematode parasitism. Plant J. 2010, 62, 1058–1071. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, P.; Bhatt, K.; Chen, W.J.; Huang, Y.; Xiao, Y.; Wu, S.; Lei, Q.; Zhong, J.; Zhu, X.; Chen, S. Bioremediation potential of laccase for catalysis of glyphosate, isoproturon, lignin, and parathion: Molecular docking, dynamics, and simulation. J. Hazard. Mater. 2023, 443, 130319. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, W.J.; Chen, S.F.; Lei, Q.; Li, J.; Bhatt, P.; Mishra, S.; Chen, S. Cellular response and molecular mechanism of glyphosate degradation by Chryseobacterium sp. Y16C. J. Agric. Food Chem. 2023, 71, 6650–6661. [Google Scholar] [CrossRef]
- Bhatt, P.; Bhatt, K.; Huang, Y.; Li, J.; Wu, S.; Chen, S. Biofilm formation in xenobiotic-degrading microorganisms. Crit. Rev. Biotechnol. 2022, 28, 1–21. [Google Scholar] [CrossRef]
- Li, J.; Chen, W.J.; Zhang, W.; Zhang, Y.; Lei, Q.; Wu, S.; Huang, Y.; Mishra, S.; Bhatt, P.; Chen, S. Effects of free or immobilized bacterium Stenotrophomonas acidaminiphila Y4B on glyphosate degradation performance and indigenous microbial community structure. J. Agric. Food Chem. 2022, 70, 13945–13958. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Medina, A.; Mbaluto, C.M.; Maedicke, A.; Weinhold, A.; Vergara, F.; van Dam, N.M. Leaf herbivory counteracts nematode-triggered repression of jasmonate-related defenses in tomato roots. Plant Physiol. 2021, 187, 1762–1778. [Google Scholar] [CrossRef] [PubMed]
- Caboni, P.; Tronci, L.; Liori, B.; Tocco, G.; Sasanelli, N.; Diana, A. Tulipaline A: Structure–activity aspects as a nematicide and V-ATPase inhibitor. Pesticide Biochem. Physiol. 2014, 112, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Naz, I.; Abdulkafi, S.; Munir, I.; Ahmad, M.; Ali, A.; Sultan, A.; Ahmad, I. Cis-and trans-protopinium, a novel nematicide, for the eco-friendly management of root-knot nematodes. Crop Protec. 2016, 81, 138–144. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Caboni, P. Botanical nematicides in the mediterranean basin. Phytochem. Rev. 2012, 11, 351–359. [Google Scholar] [CrossRef]
- Van Etten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two classes of plant antibiotics: Phytoalexins versus “phytoanticipins”. Plant Cell 1994, 6, 1191–1192. [Google Scholar] [CrossRef]
- Abawi, G.S.; Lorbeer, J.W. Pathological histology of four onion cultivars infected by Fusarium oxysporum f. sp. cepae. Phytopathology 1971, 61, 1164–1169. [Google Scholar] [CrossRef]
- Baetz, U.; Martinoia, E. Root exudates: The hidden part of plant defense. Trends Plant Sci. 1994, 19, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, D.T.; Keen, N.T.; Thomason, I.J. Association of glycollin with the incompatible response of soybean roots to Meloidogyne incognita. Physiol. Plant Pathol. 1980, 16, 309–318. [Google Scholar] [CrossRef]
- Chin, S.; Behm, C.A.; Mathesius, U. Functions of flavonoids in plant–nematode interactions. Plants 2018, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, V.K.; Rahman, A.; Dung, N.T.; Huh, M.K.; Kang, S.C. In vitro inhibition of food spoilage and foodborne pathogenic bacteria by essential oil and leaf extracts of Magnolia liliflora. J. Food Sci. 2008, 73, 314–320. [Google Scholar] [CrossRef]
- Kessler, A.; Halitschke, R.; Diezel, C.; Baldwin, I.T. Priming of plant defense responses in nature by airborne signaling between Artemisia tridentata and Nicotiana attenuata. Oecologia 2006, 148, 280–292. [Google Scholar] [CrossRef]
- Wilschut, R.A.; Silva, J.C.P.; Garbeva, P.; van der Putten, W.H. Below ground plant-herbivore interactions vary among climate-driven range-expanding plant species with different degrees of novel chemistry. Front. Plant Sci. 2017, 8, 1861. [Google Scholar] [CrossRef]
- Terra, W.C.; Campos, V.P.; Pedroso, M.P.; Costa, A.L.; Freire, E.S.; Pinto, I.P.; Silva, J.C.P.; Lopez, L.E.; Santos, T.C.N. Volatile molecules of Fusarium oxysporum strain 21 are retained in water and control Meloidogyne incognita. Biol. Control 2017, 112, 34–40. [Google Scholar] [CrossRef]
- Silva, J.C.P.; Campos, V.P.; Barros, A.F.; Pedroso, L.A.; Silva, M.F.; Souza, J.T.; Pedroso, M.P.; Medeiros, F.H.V. Performance of volatiles emitted from different plant species against juveniles and eggs of Meloidogyne incognita. Crop Protect. 2019, 116, 196–203. [Google Scholar] [CrossRef]
- Silva, J.C.P.; Campos, V.P.; Barros, A.F.; Pedroso, M.P.; Terra, W.C.; Lopez, L.E.; Souza, J.T. Plant volatiles reduce the viability of the root-knot nematode Meloidogyne incognita either directly or when retained in water. Plant Dis. 2018, 112, 2170–2179. [Google Scholar] [CrossRef] [Green Version]
- De Freitas Silva, M.; Campos, V.P.; Barros, A.F.; da Silva, J.C.P.; Pedroso, M.P.; de Jesus Silva, F.; Justino, J.C. Medicinal plant volatiles applied against the root-knot nematode Meloidogyne incognita. Crop Prot. 2020, 130, 105057. [Google Scholar] [CrossRef]
- Barros, A.F.; Oliveira, R.D.L.; Lima, I.M.; Coutinho, R.R.; Ferreira, A.O.; Costa, A. Root-knot nematodes, a growing problem for Conilon coffee in Espírito Santo state, Brazil. Crop Prot. 2014, 55, 74–79. [Google Scholar] [CrossRef]
- Mei, X.; Wang, X.; Li, G. Pathogenicity and volatile nematicidal metabolites from Duddingtonia flagrans against Meloidogyne incognita. Microorganisms 2021, 9, 2268. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Root-Knot Nematode Species | Strain Designation | Number of Predicted Genes | Assembly Size (Mb) | Number of Scaffolds | Protein-Coding Region (Mb) | GC Content (%) | References |
|---|---|---|---|---|---|---|---|
| Meloidogyne hapla | VW9 | 14,220 | 53.01 | 3450 | - | 27.4 | [45] |
| M. floridensis | - | - | 96.67 | 58,696 | - | 30.0 | [43] |
| M. incognita | W1 | 24,714 | 121.96 | 33,735 | 43.7 | 30.6 | [44] |
| M. javanica | VW4 | 26,917 | 150.35 | 34,394 | 75.2 | 30.2 | [44] |
| M. incognita | V3 | 45,351 | 183.53 | 12,091 | - | 29.8 | [39] |
| M. arenaria | HarA | 30,308 | 163.75 | 46,509 | 82.2 | 30.3 | [44] |
| M. enterolobii | L30 | 31,051 | 162.97 | 46,090 | NA | 30.2 | [44] |
| M. graminicola | IARI | 10,196 | 38.19 | 4304 | - | 23.1 | [46] |
| Effector Gene/Protein | Nematode Species | Cellular Localization in Nematode | Cellular Localization in Plant | Function in Parasitism | Ref. |
|---|---|---|---|---|---|
| Gr-pel-2 | Globodera rostochiensis | Subventral esophageal glands | Apoplast | Pectatelyases (cell-wall-degrading and migration) | [93] |
| Mi-PEL 3/Pectate lyase | Meloidogyne incognita | Subventral glands | Apoplast | Protein degradation and cell wall modification | [25] |
| Bx-crt-1 | Bursaphelenchus xylophilus | Esophageal gland | - | Calreticulin calcium-binding protein, cell-to-cell trafficking, and differentiation of NF cells. | [94] |
| Mi-CRT/Calreticulin | M. incognita | Subventral esophageal gland cells | Apoplast | Overproduction in plant cells increases plant resistance to RKNs | [95] |
| Mj-FAR-1/Fatty acid and retinol binding protein | M. javanica | Cuticle | Apoplasm | Manipulates the lipid-based signaling | [96] |
| Mj-eng-3/Beta-1,4-Endoglucanase | M. javanica | Subventral glands | Apoplasm | Degrades the cellulose of plant cell walls | [97] |
| MjTTL5 | M. javanica | Subventral gland | Plastids | Encodes a transthyretin-like protein that may suppress host defenses | [98] |
| Rs-CRT | Radopholus similis | Esophageal glands, gonads, and intestines of juveniles | - | Essential for reproduction and pathogenicity | [99] |
| Misp12 | M. incognita | Dorsal esophageal gland | Cytoplasm | Participates in the maintenance of giant cells during parasitism | [100] |
| MiMsp40 | M. incognita | Subventral esophageal gland cells | Cytoplasm and nucleus | Suppresses ETI-associated cell death | [73] |
| HaEXPB2 | Heterodera avenae | Subventral esophageal glands | Apoplast | Involvement in successful compatibility Interaction J2s | [101] |
| MeTCTP | M. enterolobii | Dorsal gland | Cytoplasm | Suppresses programmed cell death in host plants | [102] |
| MgGPP | M. graminicola | Subventral esophageal gland cells | Nucleus | Suppresses host defenses and enhances nematode parasitism | [103] |
| MiSGCR1 | M. incognita | Dorsal gland | Cytoplasm and nucleus | Suppresses plant cell death | [89] |
| Mg16820 | M. graminicola | Subventral glands | Apoplast, cytoplasm, and nucleus | Suppresses both the PTI and ETI responses | [104] |
| MiISE6 | M. incognita | Esophageal glands | Nucleus | Suppresses programmed cell death in hosts | [105] |
| MiPDI1 | M. incognita | Secreted by the esophageal glands | Cytoplasm and nucleus | Increased susceptibility and facilitates parasitism | [106] |
| MiEFF1 | M. incognita | Esophageal glands | Nucleus | Interacts with cytosolic glyceraldehyde-3-phosphate dehydrogenases to promote parasitism | [107] |
| MiEFF18 | M. incognita | Salivary glands | Nucleus | Giant cell ontogenesis | [90] |
| Mi-ISC-1 | M. incognita | Subventral esophageal glands | Cytoplasm | Disrupts the isochorismate synthase pathway for SA biosynthesis | [108] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.; Chen, S.; Fatima, S.; Ahamad, L.; Siddiqui, M.A. Biotechnological Tools to Elucidate the Mechanism of Plant and Nematode Interactions. Plants 2023, 12, 2387. https://doi.org/10.3390/plants12122387
Khan A, Chen S, Fatima S, Ahamad L, Siddiqui MA. Biotechnological Tools to Elucidate the Mechanism of Plant and Nematode Interactions. Plants. 2023; 12(12):2387. https://doi.org/10.3390/plants12122387
Chicago/Turabian StyleKhan, Arshad, Shaohua Chen, Saba Fatima, Lukman Ahamad, and Mansoor Ahmad Siddiqui. 2023. "Biotechnological Tools to Elucidate the Mechanism of Plant and Nematode Interactions" Plants 12, no. 12: 2387. https://doi.org/10.3390/plants12122387