
The Effects of Foliar Supplementation of Silicon on Physiological and Biochemical Responses of Winter Wheat to Drought Stress during Different Growth Stages
 , , and
, , and
Abstract
:1. Introduction
2. Results
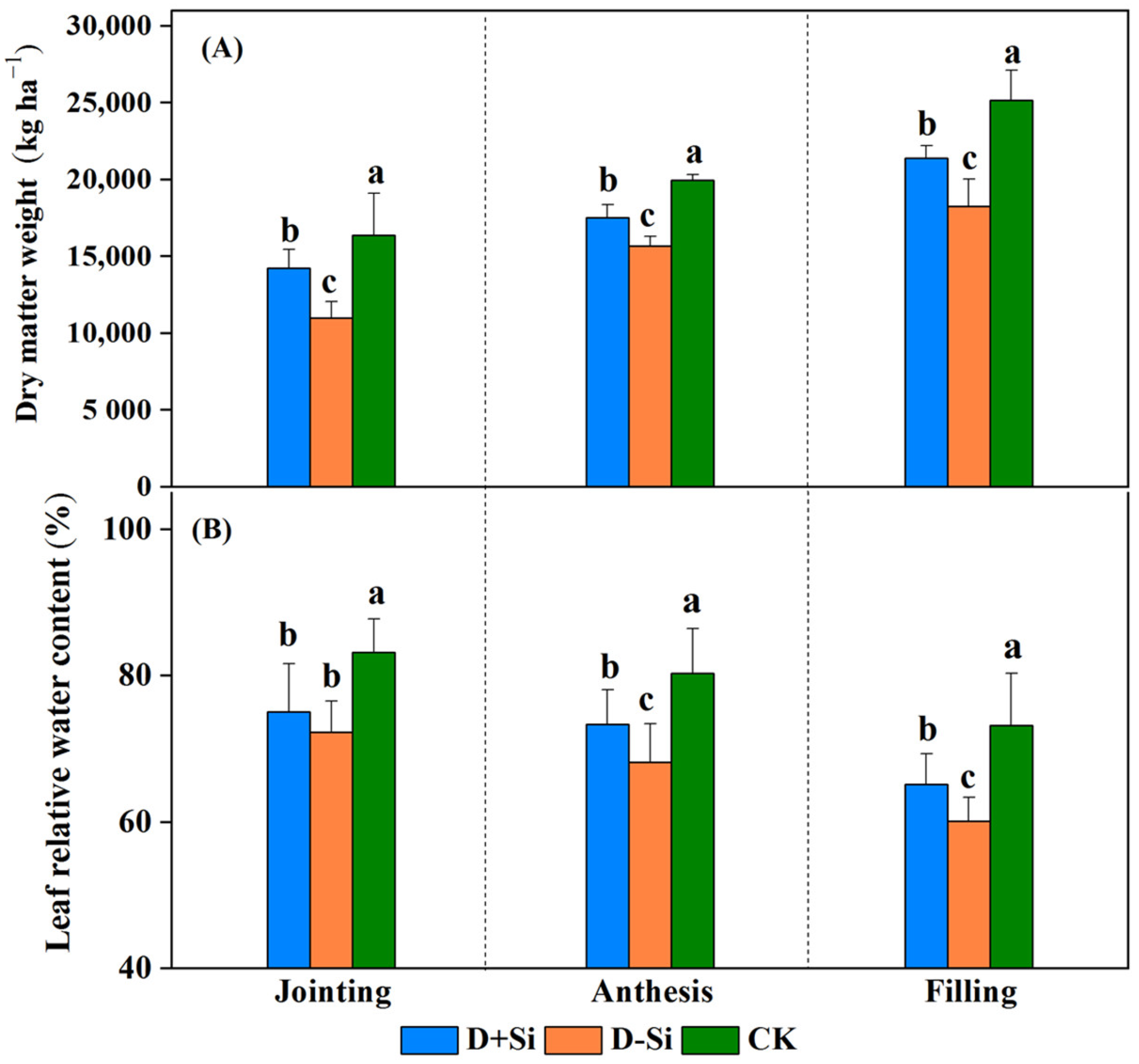
2.1. Plant Growth
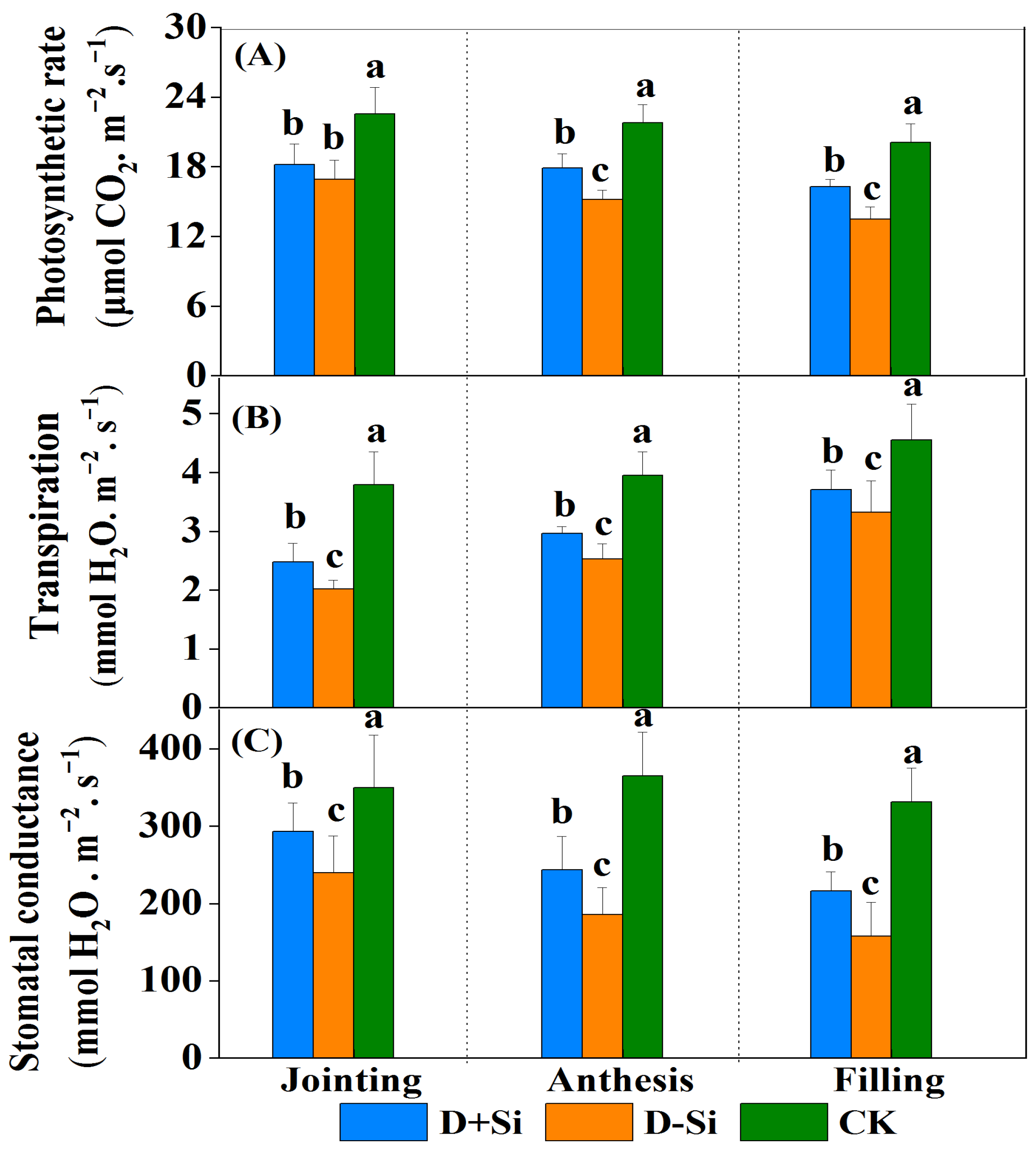
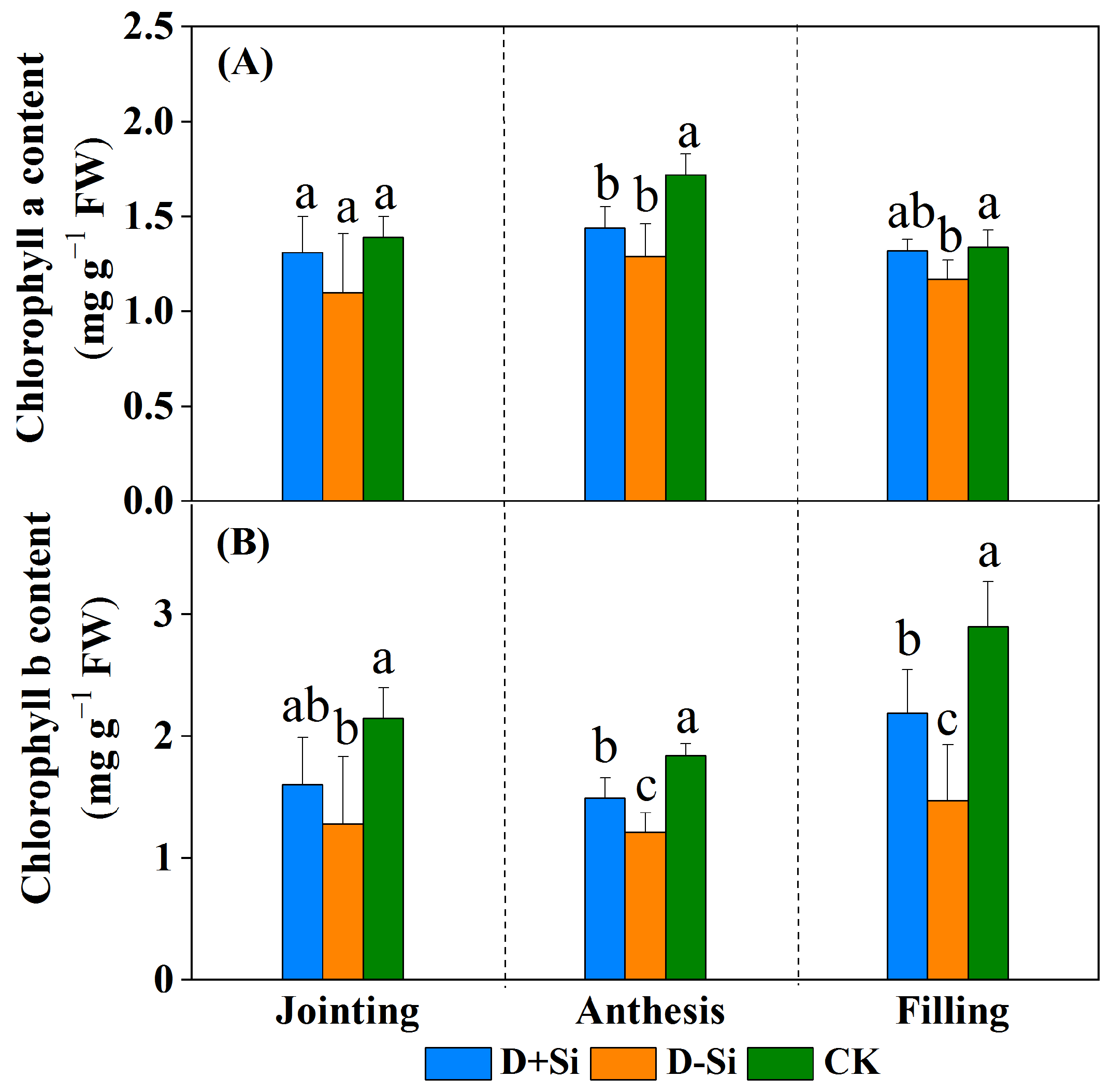
2.2. Photosynthesis and Chlorophyll Content
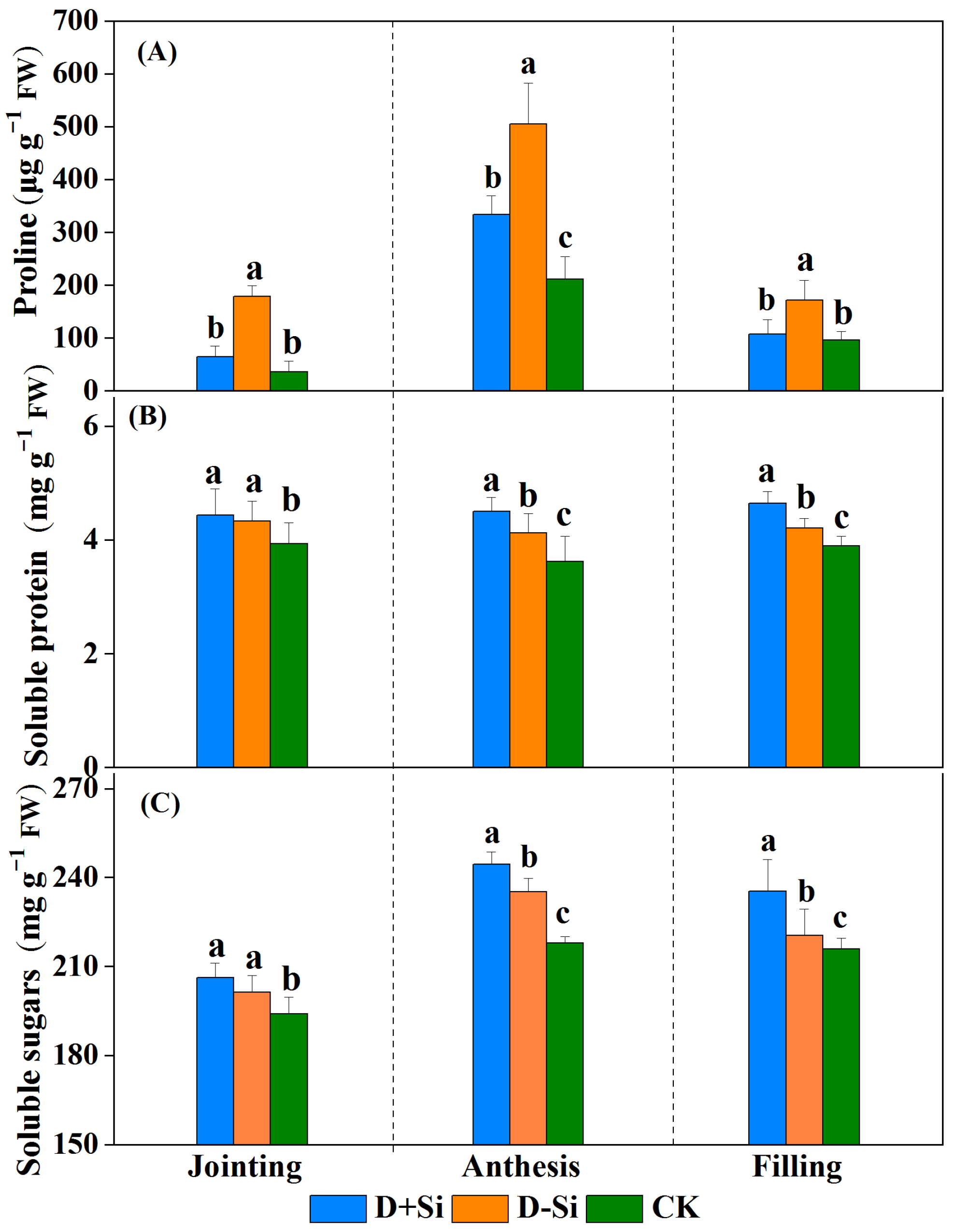
2.3. Osmotic Solute Content
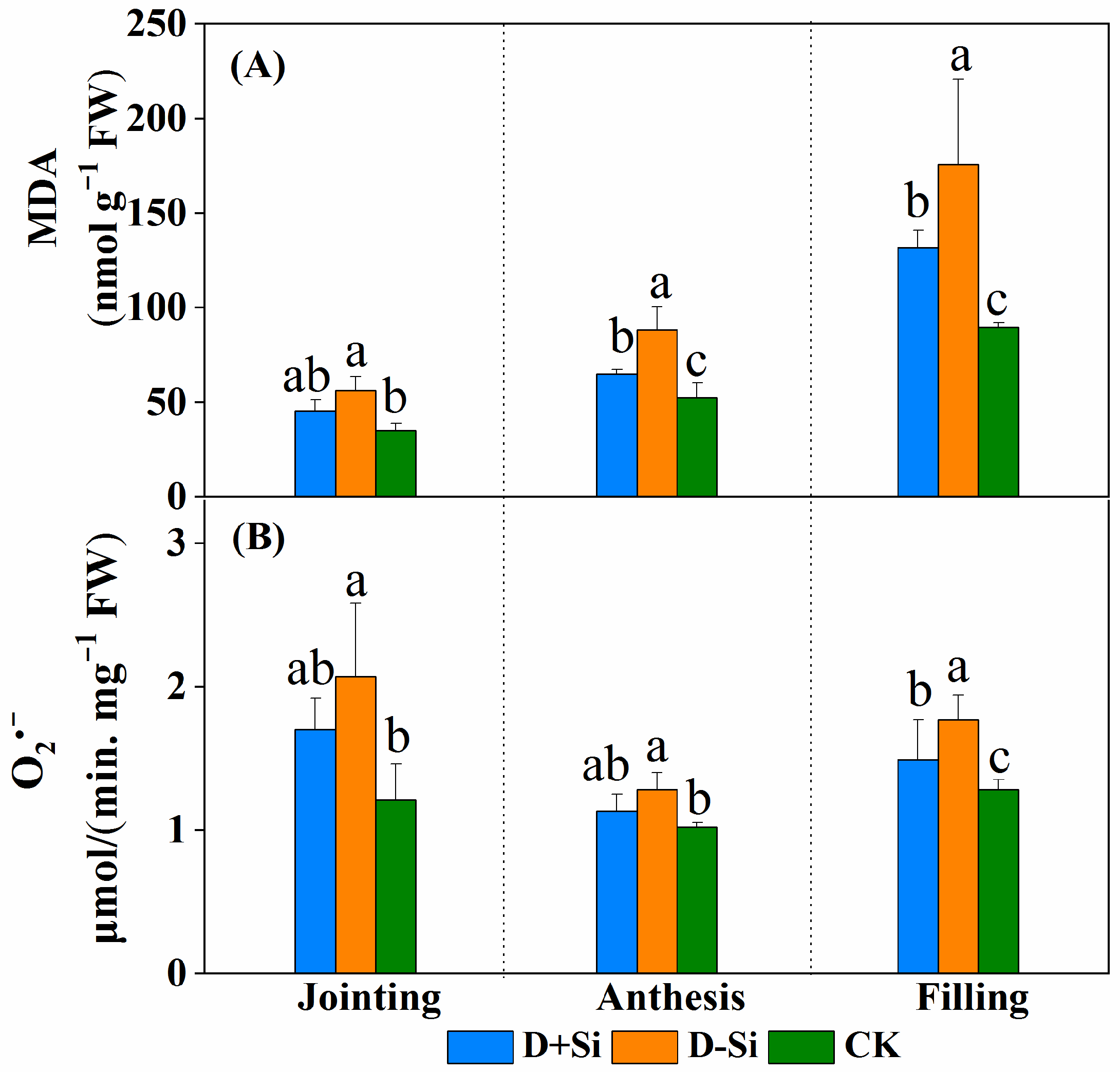
2.4. Superoxide Radicals and MDA Content
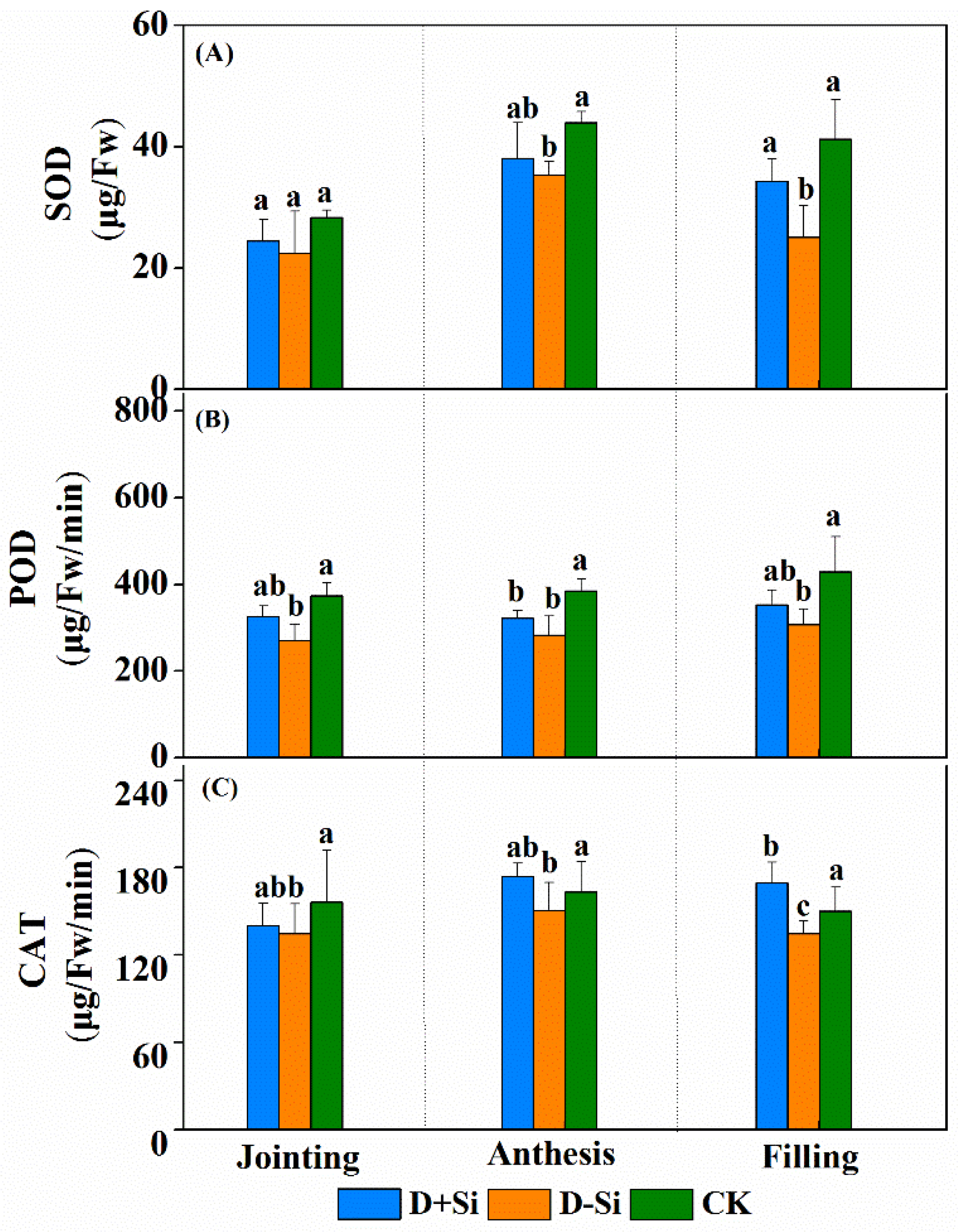
2.5. Antioxidant Enzyme Activity
2.6. Grain Yield
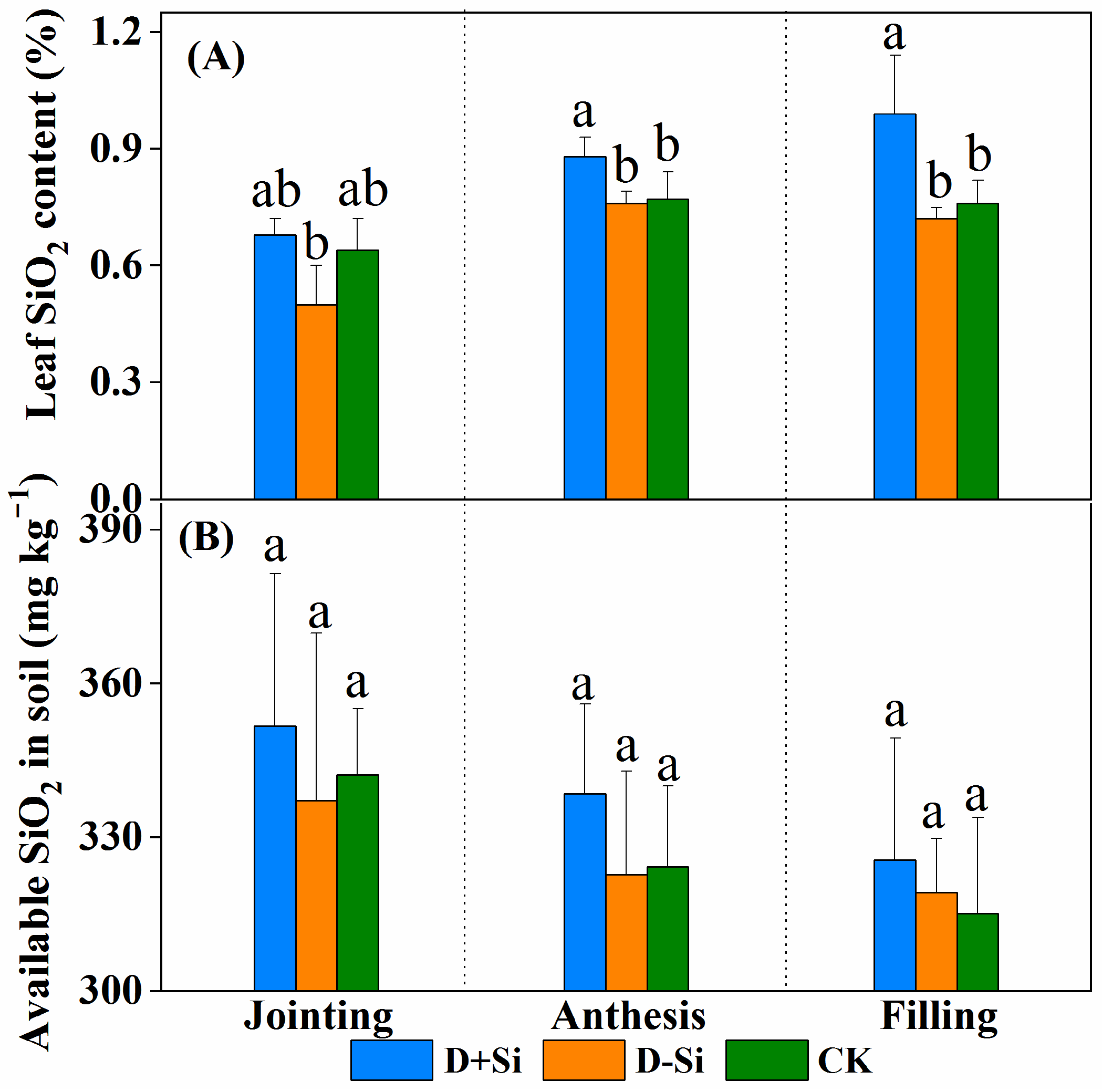
2.7. Si concentration in Plant Tissues and Soil
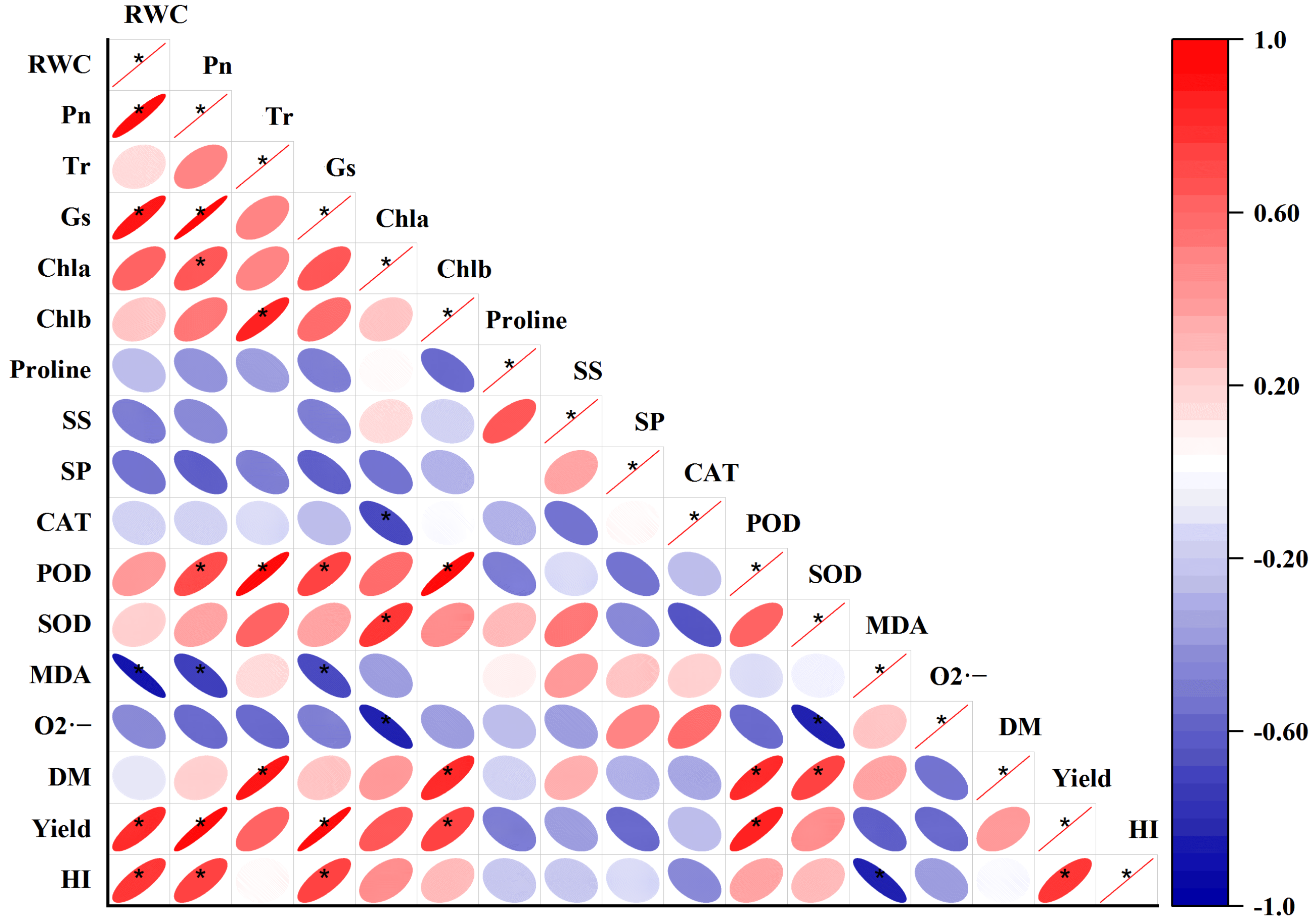
2.8. Correlation Analysis
3. Discussion
3.1. Effect of Si on Plant Growth, and Photosynthesis under Water Stress
3.2. Effect of Si on Osmotic Adjustment and Antioxidant Defense under Water Stress
3.3. Effect of Si on Wheat Biomass and Yield under Water Stress
4. Materials and Methods
4.1. Experimental Site and Treatments
4.2. Sampling and Measurements
- Leaf samples
- Dry weight of above biomass and grain yield
- Photosynthetic rate, transpiration rate and stomatal conductance
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ault, T.R. On the essentials of drought in a changing climate. Science 2020, 368, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Mukarram, M.; Choudhary, S.; Kurjak, D.; Petek, A.; Khan, M.M.A. Drought: Sensing, signalling, effects and tolerance in higher plants. Physiol. Plantarum. 2021, 172, 1291–1300. [Google Scholar] [CrossRef]
- Xiao, D.P.; Tao, F.L. Contributions of cultivars, management and climate change to winter wheat yield in the North China Plain in the past three decades. Eur. J. Agron. 2014, 52, 112–122. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought stress in wheat during flowering and grain-filling periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Wang, J.Y.; Xiong, Y.C.; Li, F.M.; Siddique, K.H.M.; Turner, N.C. Effects of drought stress on morphophysiological traits, biochemical characteristics, yield, and yield components in different ploidy wheat: A meta-analysis. Adv. Agron. 2017, 143, 139–173. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etesamia, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotox Environ. Safe 2018, 147, 881–896. [Google Scholar] [CrossRef]
- Malik, M.A.; Wani, A.H.; Mir, S.H.; Ul Rehman, I.; Tahir, I.; Ahmad, P.; Rashid, I. Elucidating the role of silicon in drought stress tolerance in plants. Plant Physiol. Bioch. 2021, 165, 187–195. [Google Scholar] [CrossRef]
- Wang, Y.W.; Zhang, B.B.; Jiang, D.X.; Chen, G.X. Silicon improves photosynthetic performance by optimizing thylakoid membrane protein components in rice under drought stress. Environ. Exp. Bot. 2019, 158, 117–124. [Google Scholar] [CrossRef]
- Meunier, J.D.; Barboni, D.; Anwar-ul-Haq, M.; Levard, C.; Chaurand, P.; Vidal, V.; Grauby, O.; Huc, R.; Laffont-Schwob, I.; Rabier, J. Effect of phytoliths for mitigating water stress in durum wheat. New Phytol. 2017, 215, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, D.F.; Qin, A.Z.; Liu, Z.D.; Duan, A.W.; Xiao, J.F.; Zhang, J.Y.; Liu, Z.G.; Zhao, B.; Liu, Z.J. Silicon-mediated physiological and agronomic responses of maize to drought stress imposed at the vegetative and reproductive stages. Agronomy 2020, 10, 1136. [Google Scholar] [CrossRef]
- Camargo, M.S.; Honaine, M.F.; Osterrieth, M.; Bozza, N.G.; Silva, V.D.; Benvenuto, M.L.; Silva, M.D. Silicon fertilization increases gas-exchange and biomass by silicophytolith deposition in the leaves of contrasting drought-tolerant sugarcane cultivars under well-watered conditions. Plant Soil. 2021, 466, 581–595. [Google Scholar] [CrossRef]
- Saja-Garbarz, D.; Libik-Konieczny, M.; Fellner, M.; Jurczyk, B.; Janowiak, F. Silicon-induced alterations in the expression of aquaporins and antioxidant system activity in well-watered and drought-stressed oilseed rape. Plant Physiol. Bioch. 2022, 174, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Biju, S.; Fuentes, S.; Gupta, D. Silicon improves seed germination and alleviates drought stress in lentil crops by regulating osmolytes, hydrolytic enzymes and antioxidant defense system. Plant Physiol. Bioch. 2017, 119, 250–264. [Google Scholar] [CrossRef]
- Helaly, M.N.; El-Hoseiny, H.; El-Sheery, N.I.; Rastogi, A.; Kalaji, H.M. Regulation and physiological role of silicon in alleviating drought stress of mango. Plant Physiol. Bioch. 2017, 118, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, Y.; Han, W.H.; Feng, R.; Hu, Y.H.; Guo, J.; Gong, H.J. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Front. Plant Sci. 2016, 7, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hameed, A.; Sheikh, M.A.; Jamil, A.; Basra, S.M.A. Seed priming with sodium silicate enhances seed germination and seedling growth in wheat (Triticum aestivum L.) under water deficit stress induced by polyethylene glycol. Pak. J. Life Soc. Sci. 2013, 11, 19–24. [Google Scholar]
- Liu, P.; Yin, L.N.; Deng, X.P.; Wang, S.W.; Tanaka, K.; Zhang, S.Q. Aquaporin-mediated increase in root hydraulic conductanceis involved in silicon-induced improved root water uptake under osmotic stress in Sorghum bicolor L. J. Exp. Bot. 2014, 65, 4747–4756. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.Q.; Guo, L.F.; Liu, L.W. Exogenous silicon alleviates drought stress in maize by improving growth, photosynthetic and antioxidant metabolism. Env. Exp. Bot. 2022, 201, 104974. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [Green Version]
- Côté-Beaulieu, C.; Chain, F.; Menzies, J.G.; Kinrade, S.D.; Bélanger, R.R. Absorption of aqueous inorganic and organic silicon compounds by wheat and their effect on growth and powdery mildew control. Environ. Exp. Bot. 2009, 65, 155–161. [Google Scholar] [CrossRef]
- Zhu, Y.X.; Gong, H.J. Beneficial effects of silicon on salt and drought tolerance in plants. Agron Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Maghsoudi, K.; Emam, Y.; Ashraf, M. Foliar application of silicon at different growth stages alters growth and yield of selected wheat cultivars. J. Plant Nutr. 2016, 39, 1194–1203. [Google Scholar] [CrossRef]
- Wasaya, A.; Hassan, J.; Yasir, T.A.; Ateeq, M.; Raza, M.A. Foliar application of silicon improved physiological indicators, yield attributes, and yield of pearl millet (Pennisetum glaucum L.) under terminal drought stress. J. Soil Sci. Plant Nutr. 2022, 22, 4458–4472. [Google Scholar] [CrossRef]
- Zhang, C.H.; Moutinho-Pereira, J.M.; Correia, C.; Coutinho, J.; Goncalves, A.; Guedes, A.; Gomes-Laranjo, J. Foliar application of Sili-K (R) increases chestnut (Castanea Spp.) growth and photosynthesis, simultaneously increasing susceptibility to water deficit. Plant Soil. 2013, 365, 211–225. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress, effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Savant, N.K.; Korndörfer, G.H.; Datnoff, L.E.; Snyder, G.H. Silicon nutrition and sugarcane production: A review. J. Plant Nutr. 1999, 22, 1853–1903. [Google Scholar] [CrossRef]
- Yin, L.N.; Wang, S.W.; Liu, P.; Wang, W.H.; Cao, D.; Deng, X.P. Silicon-mediated changes in polyamine and 1-aminocyclopropane-1-carboxylic acid are involved in silicon-induced drought resistance in Sorghum bicolor. Plant Physiol. Bioch. 2014, 80, 268–277. [Google Scholar] [CrossRef]
- Haghighi, T.M.; Saharkhiz, M.J.; Ramezanian, A.; Zarei, M. The use of silicon and mycorrhizal fungi to mitigate changes in licorice leaf micromorphology, chlorophyll fluorescence, and rutin content under water-deficit conditions. Plant Physiol. Bioch. 2023, 197, 107662. [Google Scholar] [CrossRef]
- Mastalerczuk, G.; Borawska-Jarmulowicz, B.; Darkalt, A. Changes in the physiological and morphometric characteristics and biomass distribution of forage grasses growing under conditions of drought and silicon application. Plants 2023, 12, 16. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Qin, H.; Ding, H.; Li, Y.; Guo, T. Silicon application alleviates drought stress in wheat through transcriptional regulation of multiple antioxidant defense pathways. J. Plant Growth Regul. 2016, 35, 1–10. [Google Scholar] [CrossRef]
- Gong, H.J.; Chen, K.M. The regulatory role of silicon on water relations, photosynthetic gas exchange, and carboxylation activities of wheat leaves in field drought conditions. Acta. Physiol. Plant. 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Xu, L.; Islam, F.; Ali, B.; Pei, Z.F.; Li, J.J.; Ghani, M.A.; Zhou, W.J. Silicon and water-deficit stress differentially modulate physiology and ultrastructure in wheat (Triticum aestivum L.). 3 Biotech 2017, 273, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.J.; Chen, K.M.; Zhao, Z.G.; Chen, G.C.; Zhou, W.J. Effects of silicon on defense of wheat against oxidative stress under drought at different developmental stages. Biologia. Plant. 2008, 52, 592–596. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Env. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Ali, E.F. Evaluation of proline functions in saline conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar] [CrossRef]
- Parveen, A.; Liu, W.; Hussain, S.; Asghar, J.; Perveen, S.; Xiong, Y. Silicon priming regulates morpho-physiological growth and oxidative metabolism in maize under drought stress. Plants 2019, 8, 431. [Google Scholar] [CrossRef] [Green Version]
- Pei, Z.F.; Ming, D.F.; Liu, D.; Wan, G.L.; Geng, X.X.; Gong, H.J.; Zhou, W.J. Silicon improves the tolerance to water-deficit stress induced by polyethylene glycol in wheat (Triticum aestivum L.) seedlings. J. Plant Growth Regul. 2010, 29, 106–115. [Google Scholar] [CrossRef]
- Maillard, A.; Ali, N.; Schwarzenberg, A.; Jamois, F.; Yvin, J.C.; Hosseini, S.A. Silicon transcriptionally regulates sulfur and ABA metabolism and delays leaf senescence in barley under combined sulfur deficiency and osmotic stress. Environ. Exp. Bot. 2018, 155, 394–410. [Google Scholar] [CrossRef]
- Sonobe, K.; Hattori, T.; An, P.; Tsuji, W.; Eneji, E.; Tanaka, K.; Inanaga, S. Diurnal variations in photosynthesis, stomatal conductance and leaf water relation in sorghum grown with or without silicon under water stress. J. Plant Nutr. 2009, 32, 433–442. [Google Scholar] [CrossRef]
- Thorne, S.J.; Hartley, S.E.; Maathuis, F.J.M. Is silicon a panacea for alleviating drought and salt stress in crops? Front. Plant Sci. 2020, 11, 1221. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, Y.; Yao, H.J.; Wu, J.W.; Sun, H. Silicon improves seed germination and alleviates oxidative stress of bud seedlings in tomato under water deficit stress. Plant Physiol. Bioch. 2014, 78, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Q.; Zhang, Q.; Sun, P.; Song, C.Q. Impact of droughts on winter wheat yield in different growth stages during 2001-2016 in eastern China. Int. J. Disaster Risk Sci. 2018, 9, 376–391. [Google Scholar] [CrossRef] [Green Version]
- Haider, I.; Raza, M.A.S.; Iqbal, R.; Aslam, M.U.; Habib-ur-Rahman, M.; Raja, S.; Khan, M.T.; Aslam, M.M.; Waqas, M.; Ahmad, S. Potential effects of biochar application on mitigating the drought stress implications on wheat (Triticum aestivum L.) under various growth stages. J. Saudi. Chem. Soc. 2020, 24, 974–981. [Google Scholar] [CrossRef]
- Bukhari, M.A.; Ahmad, Z.; Ashraf, M.Y.; Afzal, M.; Nawaz, F.; Nafees, M.; Jatoi, W.N.; Malghani, N.A.; Shah, A.N.; Manan, A. Silicon mitigates drought stress in wheat (Triticum aestivum L.) through improving photosynthetic pigments, biochemical and yield characters. Silicon 2021, 13, 4757–4772. [Google Scholar] [CrossRef]
- Lavinsky, A.O.; Detmann, K.C.; Reis, J.V.; Avila, R.T.; Sanglard, M.L.; Pereira, L.F.; Sanglard, L.M.V.P.; Rodrigues, F.A.; Araujo, W.L.; DaMatta, F.M. Silicon improves rice grain yield and photosynthesis specifically when supplied during the reproductive growth stage. J. Plant Physiol. 2016, 206, 125–132. [Google Scholar] [CrossRef]
- Ning, D.; Qin, A.; Duan, A.; Xiao, J.; Zhang, J.; Liu, Z.; Liu, Z.; Zhao, B.; Liu, Z. Deficit irrigation combined with reduced N-fertilizer rate can mitigate the high nitrous oxide emissions from Chinese drip-fertigated maize field. Glob. Ecol. Conserv. 2019, 20, e00803. [Google Scholar] [CrossRef]
- Qin, A.; Ning, D.; Liu, Z.; Sun, B.; Zhao, B.; Xiao, J.; Duan, A. Insentek sensor: An alternative to estimate daily crop evapotranspiration for maize plants. Water 2019, 11, 25. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Wheat Yield (kg ha−1) | Straw Biomass (kg ha−1) | Harvest Index (%) |
|---|---|---|---|
| D-jointing+Si | 10,804 ± 354 ab | 17,180 ± 780 b | 38.6 ± 0.58 a |
| D-anthesis+Si | 10,175 ± 313 b | 16,965 ± 612 b | 37.5 ± 1.15 a |
| D-filling+Si | 10,421 ± 339 bc | 17,750 ± 864 ab | 37.0 ± 1.53 a |
| D-jointing-Si | 10,352 ± 116 bc | 17,198 ± 381 b | 37.6 ± 0.58 a |
| D-anthesis-Si | 9858 ± 694 bc | 16,755 ± 665 b | 37.0 ± 0.63 a |
| D-filling-Si | 9290 ± 430 c | 18,113 ± 1362 ab | 33.9 ± 1.53 b |
| CK | 11,450 ± 1119 a | 18,840 ± 223 a | 37.8 ± 2.08 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, D.; Zhang, Y.; Li, X.; Qin, A.; Huang, C.; Fu, Y.; Gao, Y.; Duan, A. The Effects of Foliar Supplementation of Silicon on Physiological and Biochemical Responses of Winter Wheat to Drought Stress during Different Growth Stages. Plants 2023, 12, 2386. https://doi.org/10.3390/plants12122386
Ning D, Zhang Y, Li X, Qin A, Huang C, Fu Y, Gao Y, Duan A. The Effects of Foliar Supplementation of Silicon on Physiological and Biochemical Responses of Winter Wheat to Drought Stress during Different Growth Stages. Plants. 2023; 12(12):2386. https://doi.org/10.3390/plants12122386
Chicago/Turabian StyleNing, Dongfeng, Yingying Zhang, Xiaojing Li, Anzhen Qin, Chao Huang, Yuanyuan Fu, Yang Gao, and Aiwang Duan. 2023. "The Effects of Foliar Supplementation of Silicon on Physiological and Biochemical Responses of Winter Wheat to Drought Stress during Different Growth Stages" Plants 12, no. 12: 2386. https://doi.org/10.3390/plants12122386