

Expression of RsPORB Is Associated with Radish Root Color

Abstract
:1. Introduction
2. Results
2.1. Chlorophyll Accumulation Determines the Skin and Flesh Color of Green Radish Roots
2.2. RsPORB Is Highly Expressed in Green Radish Roots
2.3. RsPORB Gene Has Highly Conserved Motifs for Chlorophyll Biosynthesis
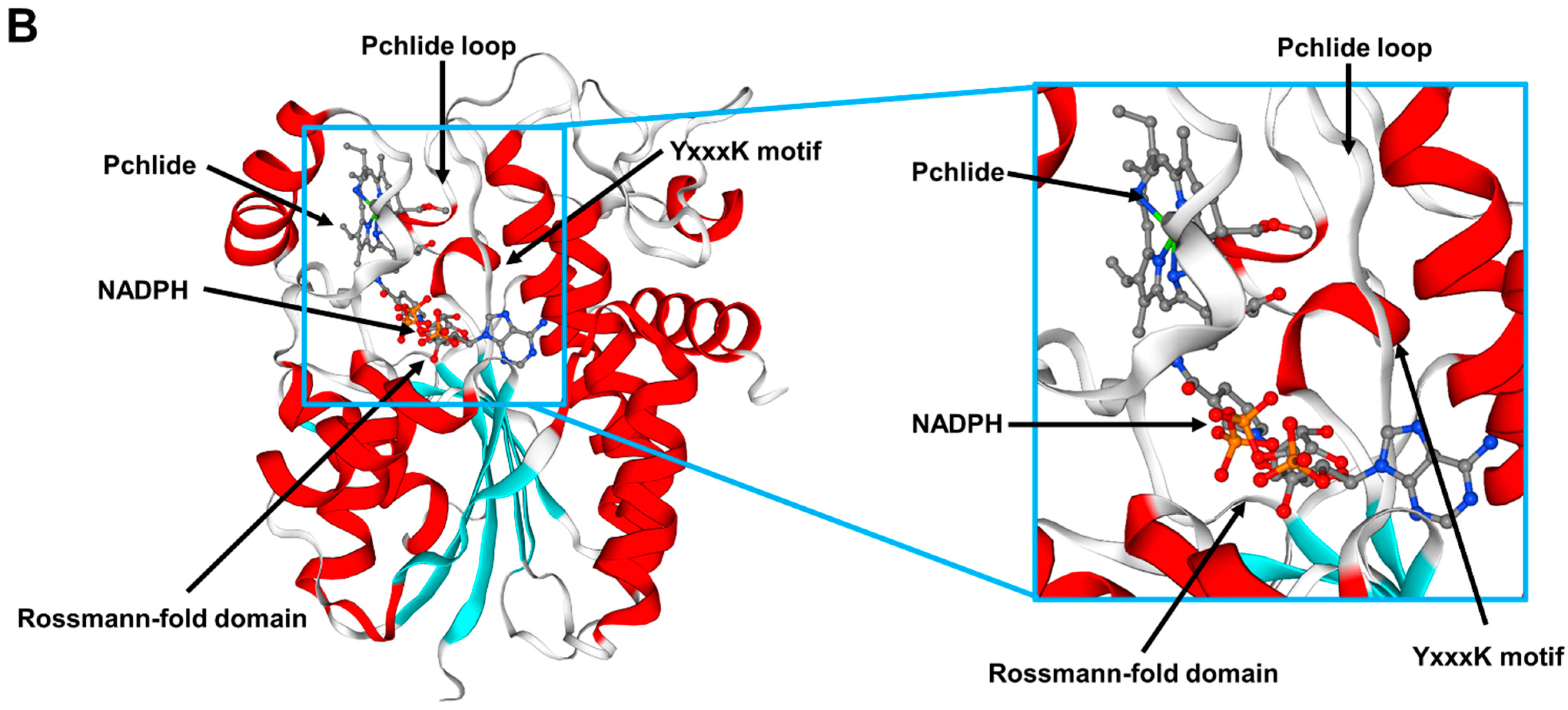
2.4. RsPORB Is Involved in Chlorophyll Biosynthesis
2.5. Promoter Region of RsPORB between Green and White Radish Was Variable
2.6. Variation of RsPORB Promoter Was Associated with Its Expression Level
3. Discussion
3.1. Expression of RsPORB Activates Chlorophyll Biosynthesis in Radish Roots
3.2. Promoter Variation of RsPORB Affects Its Transcript Level
4. Materials and Methods
4.1. Plant Materials
4.2. Measurement of Chlorophyll Contents
4.3. Total RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.4. Sequence Analysis of Promoter and Coding DNA Sequences of RsPORB
4.5. Construction of RsPORB and RsPOR VIGS Vectors and Agrobacterium-Mediated Infiltration
4.6. Promoter Activation Assay in Radish Root Protoplasts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tripathy, B.C.; Oelmüller, R. Reactive oxygen species generation and signaling in plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pospíšil, P. Production of reactive oxygen species by photosystem II as a response to light and temperature stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, H.; Zeng, T.; Zhao, Y.; Luo, T.; Deng, H.; Meng, C.; Luo, J.; Wang, C. Identification of chlorophyll metabolism-and photosynthesis-related genes regulating green flower color in chrysanthemum by integrative transcriptome and weighted correlation network analyses. Genes 2021, 12, 449. [Google Scholar] [CrossRef]
- Masuda, T.; Takamiya, K. Novel insig hts into the enzymology, regulation and physiological functions of light-dependent protochlorophyllide oxidoreductase in angiosperms. Photosynth. Res. 2004, 81, 1–29. [Google Scholar] [CrossRef]
- Scrutton, N.S.; Groot, M.L.; Heyes, D.J. Excited state dynamics and catalytic mechanism of the light-driven enzyme protochlorophyllide oxidoreductase. Phys. Chem. Chem. Phys. 2012, 14, 8818–8824. [Google Scholar] [CrossRef] [PubMed]
- Reinbothe, C.; El Bakkouri, M.; Buhr, F.; Muraki, N.; Nomata, J.; Kurisu, G.; Fijita, Y.; Reinbothe, S. Chlorophyll biosynthesis: Spotlight on protochlorophyllide reduction. Trends Plant Sci. 2010, 15, 614–624. [Google Scholar] [CrossRef]
- Reinbothe, S.; Reinbothe, C.; Holtorf, H.; Apel, K. Two NADPH: Protochlorophyllide oxidoreductases in barley: Evidence for the selective disappearance of PORA during the light-induced greening of etiolated seedlings. Plant Cell 1995, 7, 1933–1940. [Google Scholar] [CrossRef]
- Masuda, T.; Fusada, N.; Oosawa, N.; Takamatsu, K.; Yamamoto, Y.Y.; Ohto, M.; Nakamura, K.; Goto, K.; Shinata, D.; Shirano, Y.; et al. Functional analysis of isoforms of NADPH:protochlorophyllide oxidoreductase (POR), PORB and PORC, in Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 963–974. [Google Scholar] [CrossRef] [Green Version]
- Paddock, T.N.; Mason, M.E.; Lima, D.F.; Armstrong, G.A. Arabidopsis protochlorophyllide oxidoreductase A (PORA) restores bulk chlorophyll synthesis and normal development to a porB porC double mutant. Plant Mol. Biol. 2010, 72, 445–457. [Google Scholar] [CrossRef]
- Holtorf, H.; Reinbothe, S.; Reinbothe, C.; Bereza, B.; Apel, K. Two routes of chlorophyllide synthesis that are differentially regulated by light in barley (Hordeum vulgare L.). Proc. Natl. Acad. Sci. USA 1995, 92, 3254–3258. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, R.M.P.; Perez, R.L. Raphanus sativus (Radish): Their chemistry and biology. Sci. World J. 2004, 4, 811. [Google Scholar] [CrossRef] [PubMed]
- Park, N.I.; Xu, H.; Li, X.; Jang, I.H.; Park, S.; Ahn, G.H.; Lim, Y.P.; Kim, S.J.; Park, S.U. Anthocyanin accumulation and expression of anthocyanin biosynthetic genes in radish (Raphanus sativus). J. Agric. Food Chem. 2011, 59, 6034–6039. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Ki, W.; Kim, N.S.; Park, S.Y.; Kim, J.K.; Park, S.U. Metabolic Profiling of White and Green Radish Cultivars (Raphanus sativus). Horticulturae 2022, 8, 310. [Google Scholar] [CrossRef]
- Li, Y.Y.; Han, M.; Wang, R.H.; Gao, M.G. Comparative transcriptome analysis identifies genes associated with chlorophyll levels and reveals photosynthesis in green flesh of radish root. PLoS ONE 2021, 16, e0252031. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, Y.; Zhang, X.; Sun, Y.; Wang, H.; Song, J.; Li, X. Transcriptome analyses reveal key genes involved in skin color changes of ‘Xinlimei’radish root. Plant Physiol. Biochem. 2019, 139, 528–539. [Google Scholar] [CrossRef]
- Ohmiya, A.; Sasaki, K.; Nashima, K.; Oda-Yamamizo, C.; Hirashima, M.; Sumitomo, K. Transcriptome analysis in petals and leaves of chrysanthemums with different chlorophyll levels. BMC Plant Biol. 2017, 17, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, C.T.; Kim, S.H.; Song, G.; Kim, D.; Paek, N.C. Two NADPH: Protochlorophyllide Oxidoreductase (POR) Isoforms Play Distinct Roles in Environmental Adaptation in Rice. Rice 2017, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Gabruk, M.; Stecka, A.; Strzałka, W.; Kruk, J.; Strzałka, K.; Mysliwa-Kurdziel, B. Photoactive protochlorophyllide-enzyme complexes reconstituted with PORA, PORB and PORC proteins of A. thaliana: Fluorescence and catalytic properties. PLoS ONE 2015, 10, e0116990. [Google Scholar] [CrossRef] [Green Version]
- Bray, J.E.; Marsden, B.D.; Oppermann, U. The human short-chain dehydrogenase/reductase (SDR) superfamily: A bioinformatics summary. Chem. Biol. Interact. 2009, 178, 99–109. [Google Scholar] [CrossRef]
- Buhr, F.; El Bakkouri, M.; Valdez, O.; Pollmann, S.; Lebedev, N.; Reinbothe, S.; Reinbothe, C. Photoprotective role of NADPH: Protochlorophyllide oxidoreductase A. Proc. Natl. Acad. Sci. USA 2008, 105, 12629–12634. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.C.; Melo, A.A.; Kruk, J.; Frost, A.; Gabruk, M. Photocatalytic LPOR forms helical lattices that shape membranes for chlorophyll synthesis. Nat. Plants 2021, 7, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Godwin, A.R.F.; Taylor, S.; Hardman, S.J.O.; Jowitt, T.A.; Johannissen, L.O.; Hay, S.; Baldock, C.; Heyes, D.J.; Scrutton, N.S. Dual role of the active site ‘lid’ regions of protochlorophyllide oxidoreductase in photocatalysis and plant development. FEBS J. 2021, 288, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Menon, B.R.; Hardman, S.J.; Scrutton, N.S.; Heyes, D.J. Multiple active site residues are important for photochemical efficiency in the light-activated enzyme protochlorophyllide oxidoreductase (POR). J. Photochem. Photobiol 2016, 161, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Nawkar, G.M.; Kang, C.H.; Maibam, P.; Park, J.H.; Jung, Y.J.; Chae, H.B.; Chi, Y.H.; Jung, I.J.; Kim, W.Y.; Yun, D.J.; et al. HY5, a positive regulator of light signaling, negatively controls the unfolded protein response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 2084–2089. [Google Scholar] [CrossRef] [Green Version]
- Ruta, V.; Longo, C.; Lepri, A.; De Angelis, V.; Occhigrossi, S.; Costantino, P.; Vittorioso, P. The DOF transcription factors in seed and seedling development. Plants 2020, 9, 218. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.H.; Kim, S.; Lee, H.A.; Choi, D.; Yeom, S.I. Genome-wide analysis of Dof transcription factors reveals functional characteristics during development and response to biotic stresses in pepper. Sci. Rep. 2016, 6, 33332. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.; Lee, S.; Kim, T.H.; Lee, J.H.; Park, J.; Lee, J.; Lee, J.Y.; Cho, L.H.; Choi, J.Y.; Lee, W.; et al. Natural variations at the Stay-Green gene promoter control lifespan and yield in rice cultivars. Nat. Commun. 2020, 11, 2819. [Google Scholar] [CrossRef]
- Liu, W.; Liu, S.; Zhang, K.; Xie, M.; Sun, H.; Huang, X.; Zhang, L.; Li, M. Chlorophyllase is transcriptionally regulated by CsMYB308/CsDOF3 in young leaves of tea plant. Hortic. Plant J. 2022. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, J.H.; Kim, J.; Kim, J.; Lee, U.; Song, I.J.; Kim, J.H.; Lee, H.Y.; Nam, H.G.; Lim, P.O. The RAV1 transcription factor positively regulates leaf senescence in Arabidopsis. J. Exp. Bot. 2010, 61, 3947–3957. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, G.A.; Runge, S.; Frick, G.; Sperling, U.; Apel, K. Identification of NADPH:protochlorophyllide oxidoreductases A and B: A branched pathway for light-dependent chlorophyll biosynthesis in Arabidopsis thaliana. Plant Physiol. 1995, 108, 1505–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frick, G.; Su, Q.; Apel, K.; Armstrong, G.A. An Arabidopsis porB porC double mutant lacking light-dependent NADPH: Protochlorophyllide oxidoreductases B and C is highly chlorophyll-deficient and developmentally arrested. Plant J. 2003, 35, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Bang, W.Y.; Jeong, I.S.; Kim, D.W.; Im, C.H.; Ji, C.; Hwang, S.M.; Kim, S.W.; Son, Y.S.; Jeong, J.; Shiina, T.; et al. Role of Arabidopsis CHL27 protein for photosynthesis, chloroplast development and gene expression profiling. Plant Cell Physiol. 2008, 49, 1350–1363. [Google Scholar] [CrossRef] [PubMed]
- Gangappa, S.N.; Botto, J.F. The multifaceted roles of HY5 in plant growth and development. Mol. Plant 2016, 9, 1353–1365. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Obayashi, T.; Masuda, T. Role of the G-box element in regulation of chlorophyll biosynthesis in Arabidopsis roots. Plant Signal. Behav. 2012, 7, 922–926. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Bai, X.; Cheng, N.; Xiao, J.; Li, X.; Xing, Y. Wide Grain 7 increases grain width by enhancing H3K4me3 enrichment in the OsMADS1 promoter in rice (Oryza sativa L.). Plant J. 2020, 102, 517–528. [Google Scholar] [CrossRef]
- Wu, X.; Liang, Y.; Gao, H.; Wang, J.; Zhao, Y.; Hua, L.; Yuan, Y.; Wang, A.; Zhang, X.; Liu, J.; et al. Enhancing rice grain production by manipulating the naturally evolved cis-regulatory element-containing inverted repeat sequence of OsREM20. Mol. Plant 2021, 14, 997–1011. [Google Scholar] [CrossRef]
- Kim, D.H.; Park, S.; Lee, J.Y.; Ha, S.H.; Lee, J.G.; Lim, S.H. A rice B-Box protein, OsBBX14, finely regulates anthocyanin biosynthesis in rice. Int. J. Mol. Sci. 2018, 19, 2190. [Google Scholar] [CrossRef] [Green Version]
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll a and b in N, N-dimethylformamide and 80% acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Lee, J.; Rhee, J.; Lee, J.Y.; Lim, S.H. Loss of the R2R3 MYB Transcription Factor RsMYB1 Shapes Anthocyanin Biosynthesis and Accumulation in Raphanus sativus. Int. J. Mol. Sci. 2021, 22, 10927. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.H.; Kim, D.H.; Lee, J.Y. RsTTG1, a WD40 Protein, Interacts with the bHLH Transcription Factor RsTT8 to Regulate Anthocyanin and Proanthocyanidin Biosynthesis in Raphanus sativus. Int. J. Mol. Sci. 2022, 23, 11973. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motif | Sequence | Position (from ATG codon) | Recognized by | |
|---|---|---|---|---|
| G1 | W1 | |||
| CCAAT | CCAAT | −835 to −839, −310 to −314, −273 to −277 | −308 to −312, −271 to −275 | HY5 |
| Dof | AAAG | −757 to −760, −660 to −663, −333 to −336 | −797 to −800, −374 to −377, −367 to −370, −361 to −364, −355 to −358, −331 to −334 | Dof |
| CAACA | CAACA | −575 to −579 | −872 to −876, −622 to −626, −447 to −451 | RAV1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-H.; Lim, S.-H.; Lee, J.-Y. Expression of RsPORB Is Associated with Radish Root Color. Plants 2023, 12, 2214. https://doi.org/10.3390/plants12112214
Kim D-H, Lim S-H, Lee J-Y. Expression of RsPORB Is Associated with Radish Root Color. Plants. 2023; 12(11):2214. https://doi.org/10.3390/plants12112214
Chicago/Turabian StyleKim, Da-Hye, Sun-Hyung Lim, and Jong-Yeol Lee. 2023. "Expression of RsPORB Is Associated with Radish Root Color" Plants 12, no. 11: 2214. https://doi.org/10.3390/plants12112214