
Invasive and Native Plants Differentially Respond to Exogenous Phosphorus Addition in Root Growth and Nutrition Regulated by Arbuscular Mycorrhizal Fungi
 ,
,
Abstract
:1. Introduction
2. Results
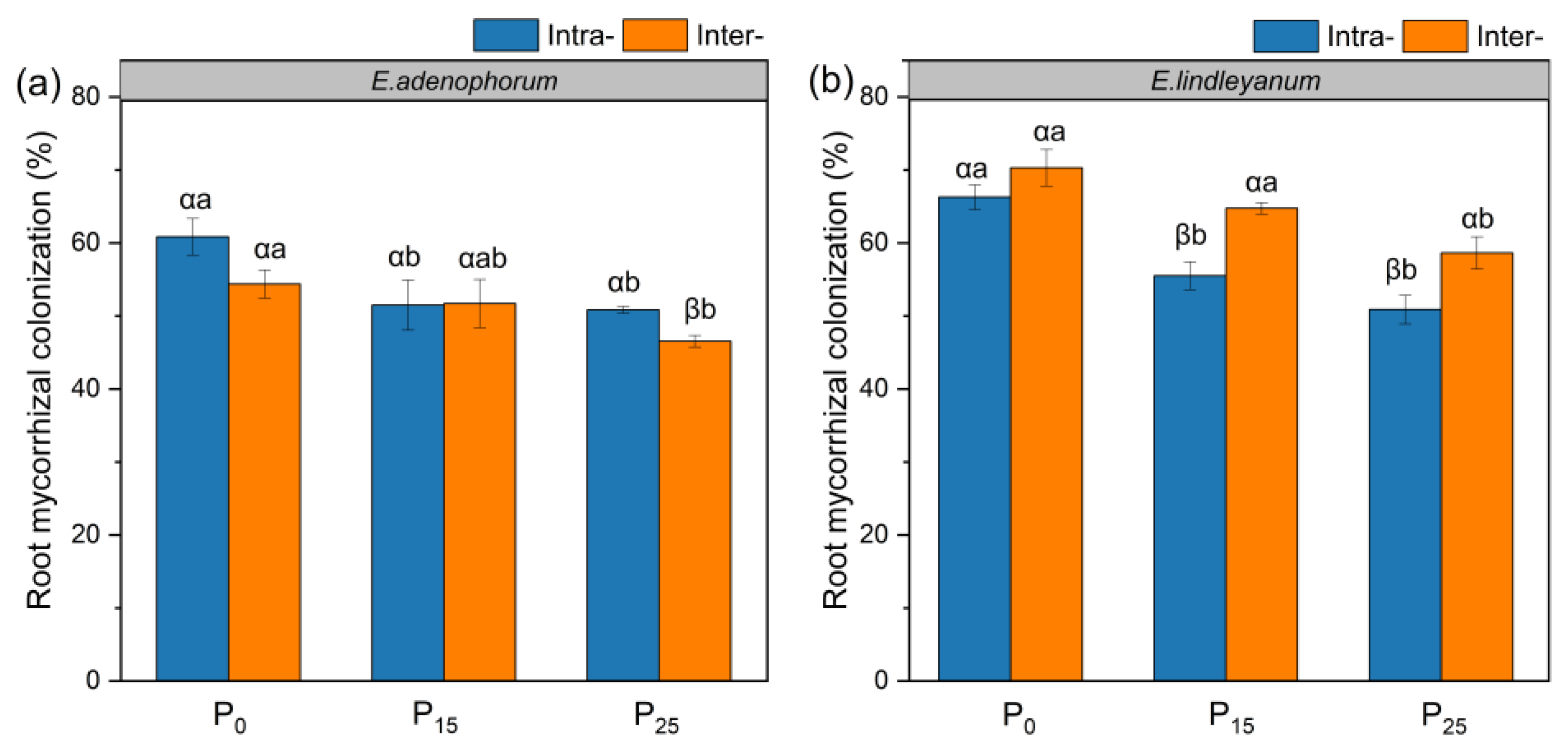
2.1. The Root Mycorrhizal Colonization of Alien Plant E. adenophorum and Native Plant E. lindleyanum
2.2. The Root Biomass of Alien Plant E. adenophorum and Native Plant E. lindleyanum
2.3. The Root Traits of Alien Plant E. adenophorum and Native Plant E. lindleyanum
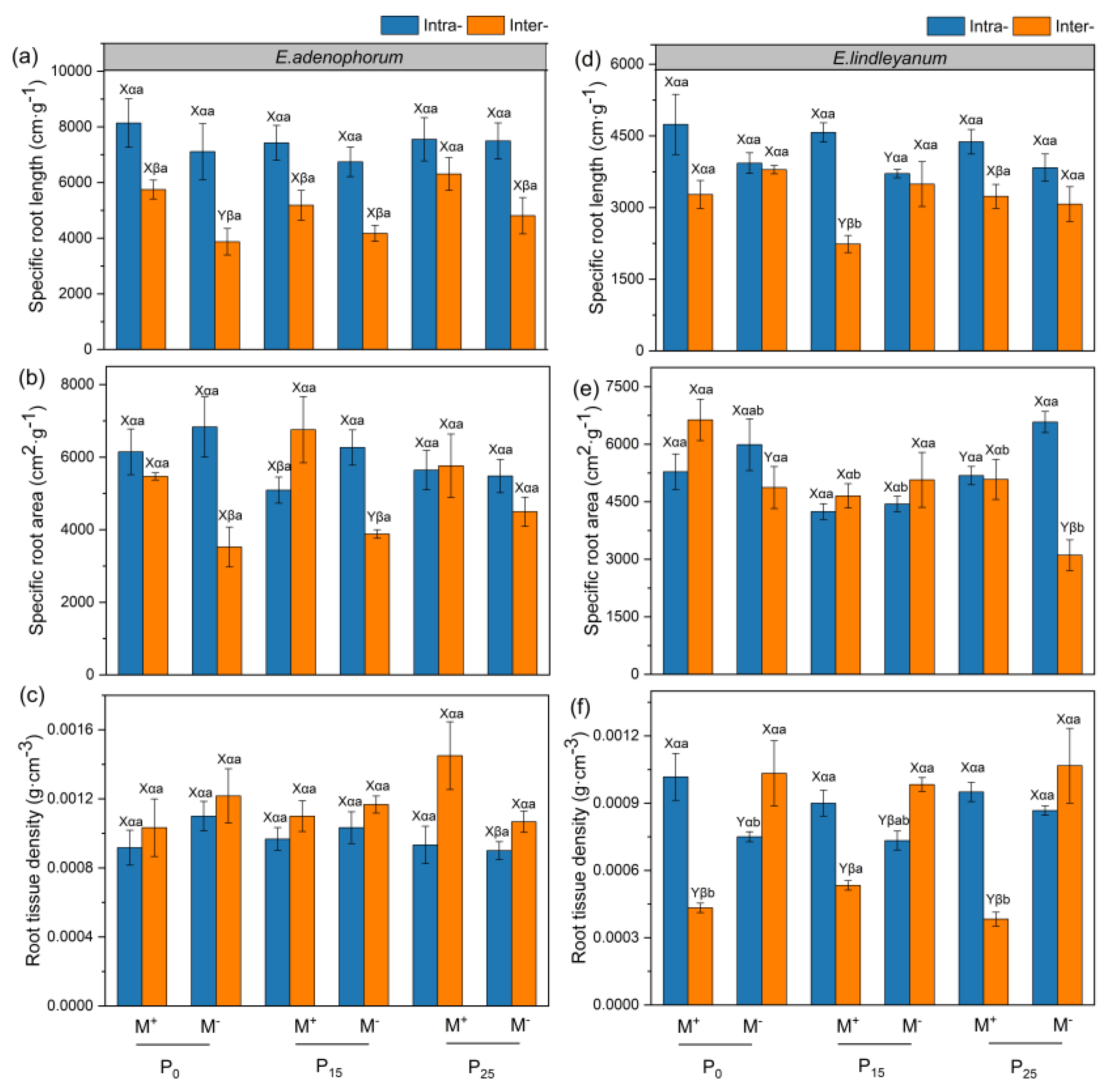
2.4. The Specific Root Traits of Alien Plant E. adenophorum and Native Plant E. lindleyanum
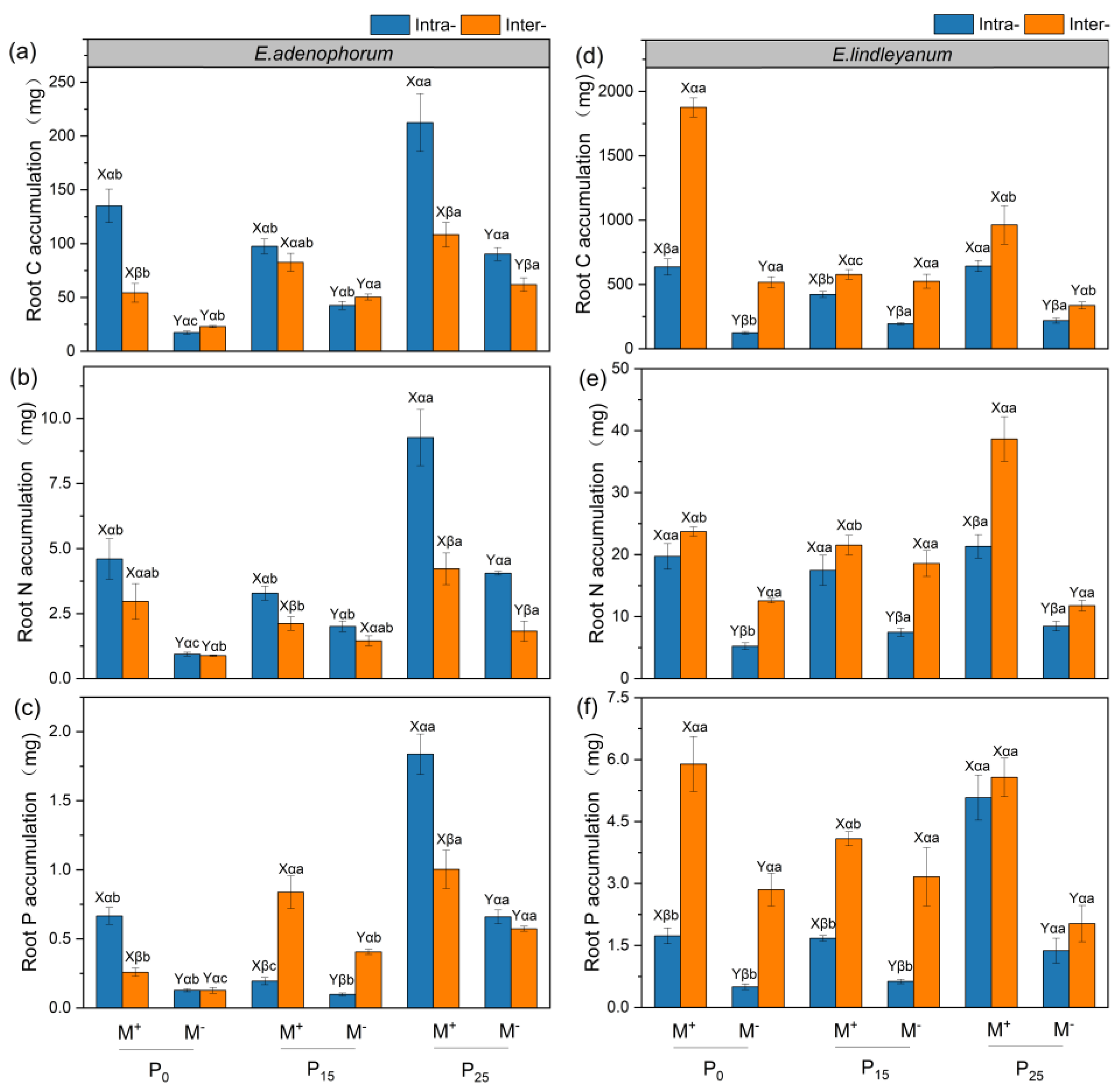
2.5. The Root C, N, and P Accumulation of Alien Plant E. adenophorum and Native Plant E. lindleyanum
3. Discussion
3.1. AMF Differently Affected the Root Growth and Nutrition of Alien and Native Plants
3.2. The Competition Differently Affects the Root Growth and Nutrition of Alien and Native Plants
3.3. Root Traits of Alien and Native Plants Respond Differently to Exogenous P Addition
3.4. The Interaction of AMF and Exogenous P Addition Differently Affect the Alien and Native Plants
4. Materials and Methods
4.1. Experimental Material
4.2. Experiment Design
4.3. Measurements and Calculations
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ni, M.; Liu, Y.; Chu, C.; Xu, H.; Fang, S. Fast seedling root growth leads to competitive superiority of invasive plants. Biol. Invasions 2018, 20, 1821–1832. [Google Scholar] [CrossRef]
- Zhang, F.; Li, Q.; Yerger, E.H.; Chen, X.; Shi, Q.; Wan, F. AM fungi facilitate the competitive growth of two invasive plant species, Ambrosia artemisiifolia and Bidens pilosa. Mycorrhiza 2018, 28, 703–715. [Google Scholar] [CrossRef]
- Xia, T.; Wang, Y.; He, Y.; Wu, C.; Shen, K.; Tan, Q.; Kang, L.; Guo, Y.; Wu, B.; Han, X. An invasive plant experiences greater benefits of root morphology from enhancing nutrient competition associated with arbuscular mycorrhizae in karst soil than a native plant. PLoS ONE 2020, 15, e0234410. [Google Scholar] [CrossRef] [PubMed]
- Dawson, W. Release from below-ground enemies and shifts in root traits as interrelated drivers of alien plant invasion success: A hypothesis. Ecol. Evol. 2015, 5, 4505–4516. [Google Scholar] [CrossRef] [PubMed]
- Morris, T.L.; Esler, K.J.; Barger, N.N.; Jacobs, S.M.; Cramer, M.D. Ecophysiological traits associated with the competitive ability of invasive Australian acacias. Divers. Distrib. 2011, 17, 898–910. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Martin, C.E.; Falasco, M.R.; James, J.J. Variation in resource acquisition and utilization traits between native and invasive perennial forbs. Am. J. Bot. 2008, 95, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, A.; Stevens, C.J.; Peltzer, D.A.; Ostle, N.J.; Orwin, K.H. Below-ground competition drives invasive plant impact on native species regardless of nitrogen availability. Oecologia 2018, 186, 577–587. [Google Scholar] [CrossRef]
- Van, D.; Bradford, M.A.; Brinkman, E.P.; Van, D.; Veen, G. Where, when and how plant–soil feedback matters in a changing world. Funct. Ecol. 2016, 30, 1109–1121. [Google Scholar]
- Péret, B.; Clément, M.; Nussaume, L.; Desnos, T. Root developmental adaptation to phosphate starvation: Better safe than sorry. Trends Plant Sci. 2011, 16, 442–450. [Google Scholar] [CrossRef]
- Kumar, A.; Shahbaz, M.; Blagodatskaya, E.; Pausch, J.; Sabine, S.J. Root trait plasticity and plant nutrient acquisition in phosphorus limited soil. J. Plant Nutr. Soil Sci. 2019, 182, 945–952. [Google Scholar] [CrossRef]
- Betekhtina, A.A.; Ronzhina, D.A.; Ivanova, L.A.; Malygin, M.V.; Ivanov, L.A. Relative growth rate and its components in invasive species Heracleum sosnowskyi and congeneric native species H. sibiricum. Russ. J. Biol. Invasions 2019, 10, 5–11. [Google Scholar] [CrossRef]
- Reich, P.B. The worldwide ‘fast–slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Sánchez-Calderón, L.; Chacon-López, A.; Pérez-Torres, C.-A.; Herrera-Estrella, L. Phosphorus: Plant Strategies to Cope with Its Scarcity. In Cell Biology of Metals and Nutrients; Hell, R., Mendel, R.-R., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 173–198. [Google Scholar]
- Seabloom, E.W.; Borer, E.T.; Buckley, Y.M.; Cleland, E.E.; Davies, K.F.; Firn, J.; Harpole, W.S.; Hautier, Y.; Lind, E.M.; MacDougall, A.S. Plant species’ origin predicts dominance and response to nutrient enrichment and herbivores in global grasslands. Nat. Commun. 2015, 6, 7710. [Google Scholar] [CrossRef] [PubMed]
- Akin Fajiye, M.; Schmidt, A.C.; Fraser, L.H. Soil nutrients and variation in biomass rather than native species richness influence introduced plant richness in a semi-arid grassland. Basic Appl. Ecol. 2021, 53, 62–73. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, R.; Huangfu, C.; Jiang, I.N.; Tu, C.; Yang, D. Effects of applying phosphorus and fungicide on the growth and physiological characteristics of Flaveria bidentis. Chin. J. Ecol. 2012, 31, 2810. [Google Scholar]
- Rausch, C.; Bucher, M. Molecular mechanisms of phosphate transport in plants. Planta 2002, 216, 23–37. [Google Scholar] [CrossRef]
- Balemi, T.; Negisho, K. Management of soil phosphorus and plant adaptation mechanisms to phosphorus stress for sustainable crop production: A review. J. Soil Sci. Plant Nut. 2012, 12, 547–562. [Google Scholar] [CrossRef]
- Shen, J.; Yuan, L.; Zhang, J.; Li, H.; Bai, Z.; Chen, X.; Zhang, W.; Zhang, F. Phosphorus dynamics: From soil to plant. Plant Physiol. 2011, 156, 997–1005. [Google Scholar] [CrossRef]
- Lugli, L.F.; Andersen, K.M.; Aragão, L.E.; Cordeiro, A.L.; Cunha, H.F.; Fuchslueger, L.; Meir, P.; Mercado, L.M.; Oblitas, E.; Quesada, C.A. Multiple phosphorus acquisition strategies adopted by fine roots in low-fertility soils in Central Amazonia. Plant Soil. 2020, 450, 49–63. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Strack, D.; Fester, T.; Hause, B.; Schliemann, W.; Walter, M.H. Review Paper: Arbuscular Mycorrhiza: Biological, Chemical, and Molecular Aspects. J. Chem. Ecol. 2003, 29, 1955–1979. [Google Scholar] [CrossRef]
- Yongchun, S.; Xiaolin, L.I.; Gu, F.; Fusuo, Z.; Peter, C. Rapid assessment of acid phosphatase activity in the mycorrhizosphere and in arbuscular mycorrhizal fungal hyphae. Sci. Bull. Engl. Version 2000, 5, 1187–1191. [Google Scholar]
- Ferrol, N.; Azcón-Aguilar, C.; Pérez-Tienda, J. Review: Arbuscular mycorrhizas as key players in sustainable plant phosphorus acquisition: An overview on the mechanisms involved. Plant Sci. 2019, 280, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A.; Jakobsen, I. Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol. 2003, 133, 16–20. [Google Scholar] [CrossRef]
- Cheng, J.K.; Yue, M.F.; Yang, H.R.; Chen, B.M.; Xin, G.R. Do arbuscular mycorrhizal fungi help the native species Bidens biternata resist the invasion of Bidens alba? Plant Soil 2019, 444, 443–455. [Google Scholar] [CrossRef]
- Sun, D.; Yang, X.; Wang, Y.; Fan, Y.; Ding, P.; Song, X.E.; Yuan, X.; Yang, X. Stronger mutualistic interactions with arbuscular mycorrhizal fungi help Asteraceae invaders outcompete the phylogenetically related natives. New Phytol. 2022, 236, 1487–1496. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Tu, C.; Chen, X.; Hu, S. Arbuscular mycorrhizal fungi enhance P uptake and alter plant morphology in the invasive plant Microstegium vimineum. Biol. Invasions 2014, 16, 1083–1093. [Google Scholar] [CrossRef]
- Busso, M.A.; Busso, M. Arbuscular mycorrhizal fungi and common mycorrhizal networks benefit plants through morphological, physiological and productive traits and soil quality. Lilloa 2022, 59, 301–317. [Google Scholar] [CrossRef]
- Penuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; Van Der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef]
- López-Bucio, J.; Cruz-Ramırez, A.; Herrera-Estrella, L. The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 2003, 6, 280–287. [Google Scholar] [CrossRef]
- Zhang, F.-J.; Li, Q.; Chen, F.-X.; Xu, H.-Y.; Wan, F.-H. Arbuscular mycorrhizal fungi facilitate growth and competitive ability of an exotic species Flaveria bidentis. Soil Biol. Biochem. 2017, 115, 275–284. [Google Scholar] [CrossRef]
- Řezáčová, V.; Řezáč, M.; Gryndlerová, H.; Wilson, G.W.; Michalová, T. Arbuscular mycorrhizal fungi favor invasive Echinops sphaerocephalus when grown in competition with native Inula conyzae. Sci. Rep. 2020, 10, 20287. [Google Scholar] [PubMed]
- Alvarez Flores, R.; Winkel, T.; Nguyen Thi Truc, A.; Joffre, R. Root foraging capacity depends on root system architecture and ontogeny in seedlings of three Andean Chenopodium species. Plant Soil 2014, 380, 415–428. [Google Scholar] [CrossRef]
- Chandrasekaran, M. Arbuscular mycorrhizal fungi mediated enhanced biomass, root morphological traits and nutrient uptake under drought stress: A meta-analysis. J. Fungi 2022, 8, 660. [Google Scholar] [CrossRef] [PubMed]
- Martín Robles, N.; Lehmann, A.; Seco, E.; Aroca, R.; Rillig, M.C.; Milla, R. Impacts of domestication on the arbuscular mycorrhizal symbiosis of 27 crop species. New Phytol. 2018, 218, 322–334. [Google Scholar] [PubMed]
- Maherali, H. Is there an association between root architecture and mycorrhizal growth response? New Phytol. 2014, 204, 192–200. [Google Scholar] [CrossRef]
- Qi, S.; He, F.; Wang, J.; Li, Q.; Dai, Z.; Du, D. Effects of arbuscular mycorrhizal fungi on the growth and the competition of an invasive plant Wedelia trilobata. Microbiol. China 2020, 47, 3801–3810. [Google Scholar]
- Shen, K.; Cornelissen, J.H.C.; Wang, Y.; Wu, C.; He, Y.; Ou, J.; Tan, Q.; Xia, T.; Kang, L.; Guo, Y. AM fungi alleviate phosphorus limitation and enhance nutrient competitiveness of invasive plants via mycorrhizal networks in karst areas. Front. Ecol. Evol. 2020, 8, 125. [Google Scholar]
- Jabborova, D.; Annapurna, K.; Paul, S.; Kumar, S.; Saad, H.A.; Desouky, S.; Ibrahim, M.F.; Elkelish, A. Beneficial features of biochar and arbuscular mycorrhiza for improving spinach plant growth, root morphological traits, physiological properties, and soil enzymatic activities. J. Fungi 2021, 7, 571. [Google Scholar] [CrossRef]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity 2020, 12, 370. [Google Scholar]
- Fortunel, C.; Fine, P.V.; Baraloto, C. Leaf, stem and root tissue strategies across 758 N eotropical tree species. Funct. Ecol. 2012, 26, 1153–1161. [Google Scholar] [CrossRef]
- Li, Q.; Umer, M.; Guo, Y.; Shen, K.; Xia, T.; Xu, X.; Han, X.; Ren, W.; Sun, Y.; Wu, B. Karst soil patch heterogeneity with gravels promotes plant root development and nutrient utilization associated with arbuscular mycorrhizal fungi. Agronomy 2022, 12, 1063. [Google Scholar] [CrossRef]
- Ren, G.-Q.; Li, Q.; Li, Y.; Li, J.; Adomako, M.O.; Dai, Z.-C.; Li, G.-L.; Wan, L.-Y.; Zhang, B.; Zou, C.B. The enhancement of root biomass increases the competitiveness of an invasive plant against a co-occurring native plant under elevated nitrogen deposition. Flora 2019, 261, 151486. [Google Scholar] [CrossRef]
- Wang, Y.J.; Chen, D.; Yan, R.; Yu, F.H.; van Kleunen, M. Invasive alien clonal plants are competitively superior over co-occurring native clonal plants. Perspect Plant Ecol. 2019, 40, 125484. [Google Scholar] [CrossRef]
- Young, S.L.; Barney, J.N.; Kyser, G.B.; Jones, T.S.; DiTomaso, J.M. Functionally similar species confer greater resistance to invasion: Implications for grassland restoration. Restor. Ecol. 2009, 17, 884–892. [Google Scholar] [CrossRef]
- Bunn, R.A.; Ramsey, P.W.; Lekberg, Y. Do native and invasive plants differ in their interactions with arbuscular mycorrhizal fungi? A meta-analysis. J. Ecol. 2015, 103, 1547–1556. [Google Scholar] [CrossRef]
- Ostonen, I.; Rosenvald, K.; Helmisaari, H.S.; Godbold, D.; Parts, K.; Uri, V.; Lõhmus, K. Morphological plasticity of ectomycorrhizal short roots in Betula sp and Picea abies forests across climate and forest succession gradients: Its role in changing environments. Front. Ecol. Evol. 2013, 4, 335. [Google Scholar] [CrossRef]
- Colom, S.M.; Baucom, R.S. Below-ground competition can influence the evolution of root traits. Am. Nat. 2020, 195, 577–590. [Google Scholar] [CrossRef]
- Danieli-Silva, A.; Uhlmann, A.; Vicente-Silva, J.; Stürmer, S.L. How mycorrhizal associations and plant density influence intra-and inter-specific competition in two tropical tree species: Cabralea canjerana (Vell.) Mart. and Lafoensia pacari A. St.-Hil. Plant Soil 2010, 330, 185–193. [Google Scholar] [CrossRef]
- Sielaff, A.C.; Polley, H.W.; Fuentes-Ramirez, A.; Hofmockel, K.; Wilsey, B.J. Mycorrhizal colonization and its relationship with plant performance differs between exotic and native grassland plant species. Biol. Invasions 2019, 21, 1981–1991. [Google Scholar] [CrossRef]
- Treseder, K.K. The extent of mycorrhizal colonization of roots and its influence on plant growth and phosphorus content. Plant Soil. 2013, 371, 1–13. [Google Scholar] [CrossRef]
- Luo, B.; Ma, P.; Nie, Z.; Zhang, X.; He, X.; Ding, X.; Feng, X.; Lu, Q.; Ren, Z.; Lin, H. Metabolite profiling and genome-wide association studies reveal response mechanisms of phosphorus deficiency in maize seedling. Plant J. 2019, 97, 947–969. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, P.; Herrmann, L.; Lesueur, D.; Robin, A.; Trap, J.; Waithaisong, K.; Plassard, C. Impact of roots, microorganisms and microfauna on the fate of soil phosphorus in the rhizosphere. Ann. Plant Rev. Online 2015, 48, 375–407. [Google Scholar]
- Nahar, K.; Bovill, B.; McDonald, G. Mycorrhizal colonization in bread wheat varieties differing in their response to phosphorus. J. Plant Nutr. 2020, 44, 29–45. [Google Scholar] [CrossRef]
- Wyatt, G.A.; Kiers, E.T.; Gardner, A.; West, S.A. A biological market analysis of the plant-mycorrhizal symbiosis. Evolution 2014, 68, 2603–2618. [Google Scholar] [CrossRef] [PubMed]
- Marler, M.J.; Zabinski, C.A.; Callaway, R.M. Mycorrhizae indirectly enhance competitive effects of an invasive forb on a native bunchgrass. Ecology 1999, 80, 1180–1186. [Google Scholar] [CrossRef]
- Harpole, W.S. Resource-ratio theory and the control of invasive plants. Plant Soil 2006, 280, 23–27. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Harpole, W.S.; Reichman, O.; Tilman, D. Invasion, competitive dominance, and resource use by exotic and native California grassland species. Proc. Natl. Acad. Sci. USA 2003, 100, 13384–13389. [Google Scholar] [CrossRef]
- Zheng, Y.L.; Burns, J.H.; Wang, R.F.; Feng, Y.L. Identity recognition and the invasion of exotic plant. Flora 2021, 280, 151828. [Google Scholar] [CrossRef]
- Huang, Q.Q.; Shen, Y.D.; Li, X.X.; Li, S.L.; Fan, Z.W. Invasive Eupatorium catarium and Ageratum conyzoides benefit more than does a common native plant from nutrient addition in both competitive and non-competitive environments. Ecol. Res. 2016, 31, 145–152. [Google Scholar] [CrossRef]
- Pang, J.; Ryan, M.H.; Tibbett, M.; Cawthray, G.R.; Siddique, K.H.; Bolland, M.D.; Denton, M.D.; Lambers, H. Variation in morphological and physiological parameters in herbaceous perennial legumes in response to phosphorus supply. Plant Soil 2010, 331, 241–255. [Google Scholar] [CrossRef]
- Hodge, A.; Berta, G.; Doussan, C.; Merchan, F.; Crespi, M. Plant root growth, architecture and function. Plant Soil 2009, 321, 153–187. [Google Scholar] [CrossRef]
- Xiang, Y.; Deng, Q.; Duan, H.; Guo, Y. Effects of biochar application on root traits: A meta-analysis. GCB Bioenergy 2017, 9, 1563–1572. [Google Scholar] [CrossRef]
- Penuelas, J.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Sardans, J. Anthropogenic global shifts in biospheric N and P concentrations and ratios and their impacts on biodiversity, ecosystem productivity, food security, and human health. Glob. Change Biol. 2020, 26, 1962–1985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Leng, Z.; Wu, Y.; Jia, H.; Yan, C.; Wang, X.; Ren, G.; Wu, G.; Li, J. Interaction between nitrogen, phosphorus, and invasive alien plants. Sustainability 2022, 14, 746. [Google Scholar] [CrossRef]
- Lannes, L.S.; Karrer, S.; Teodoro, D.A.; Bustamante, M.M.; Edwards, P.J.; Olde Venterink, H. Species richness both impedes and promotes alien plant invasions in the Brazilian Cerrado. Sci. Rep. 2020, 10, 11365. [Google Scholar] [CrossRef]
- Cordell, D.; White, S. Peak phosphorus: Clarifying the key issues of a vigorous debate about long-term phosphorus security. Sustainability 2011, 3, 2027–2049. [Google Scholar] [CrossRef]
- National Soil Census Office. Chinese Soil; China Agriculture Press: Beijing, China, 1998; p. 1254. (In Chinese) [Google Scholar]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Bao, S. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specific Root Traits | Treatments | df | E. adenophorum | E. lindleyanum | ||
|---|---|---|---|---|---|---|
| F | P | F | P | |||
| Specific root length | M | 1 | 2.569 | 0.114 | 0.835 | 0.364 |
| C | 1 | 31.377 | 0.000 *** | 13.612 | 0.000 *** | |
| P | 2 | 0.938 | 0.397 | 11.365 | 0.000 *** | |
| M × C | 1 | 0.643 | 0.426 | 0.032 | 0.858 | |
| M × P | 2 | 0.668 | 0.517 | 4.637 | 0.013 * | |
| C × P | 2 | 0.732 | 0.485 | 0.803 | 0.453 | |
| M × C × P | 2 | 0.212 | 0.809 | 5.453 | 0.007 ** | |
| Specific root area | M | 1 | 2.349 | 0.131 | 0.369 | 0.546 |
| C | 1 | 16.627 | 0.000 *** | 1.056 | 0.308 | |
| P | 2 | 1.356 | 0.265 | 9.109 | 0.000 *** | |
| M × C | 1 | 3.166 | 0.080 | 33.673 | 0.000 *** | |
| M × P | 2 | 0.008 | 0.992 | 0.465 | 0.631 | |
| C × P | 2 | 3.129 | 0.051 | 10.872 | 0.000 *** | |
| M × C × P | 2 | 0.780 | 0.463 | 0.510 | 0.603 | |
| Root tissue density | M | 1 | 0.206 | 0.651 | 36.481 | 0.000 *** |
| C | 1 | 11.177 | 0.001 ** | 1.052 | 0.309 | |
| P | 2 | 0.066 | 0.936 | 6.205 | 0.004 ** | |
| M × C | 1 | 1.242 | 0.270 | 45.208 | 0.000 *** | |
| M × P | 2 | 3.232 | 0.046 | 3.997 | 0.023 * | |
| C × P | 2 | 0.935 | 0.398 | 4.977 | 0.010 * | |
| M × C × P | 2 | 0.598 | 0.553 | 4.328 | 0.018 * | |
| Root Nutrient Accumulation | Treatments | df | E. adenophorum | E. lindleyanum | ||
|---|---|---|---|---|---|---|
| F | P | F | P | |||
| C accumulation | M | 1 | 119.019 | 0.000 *** | 233.892 | 0.000 *** |
| C | 1 | 33.756 | 0.000 *** | 148.242 | 0.000 *** | |
| P | 2 | 36.721 | 0.000 *** | 38.854 | 0.000 *** | |
| M × C | 1 | 24.193 | 0.000 *** | 16.552 | 0.000 *** | |
| M × P | 2 | 3.962 | 0.024 * | 45.709 | 0.000 *** | |
| C × P | 2 | 8.555 | 0.001 ** | 32.710 | 0.000 *** | |
| M × C × P | 2 | 2.424 | 0.097 | 19.514 | 0.000 *** | |
| N accumulation | M | 1 | 74.996 | 0.000 *** | 125.144 | 0.000 *** |
| C | 1 | 38.926 | 0.000 *** | 51.799 | 0.000 *** | |
| P | 2 | 31.699 | 0.000 *** | 8.525 | 0.001 ** | |
| M × C | 1 | 8.413 | 0.005 ** | 0.022 | 0.882 | |
| M × P | 2 | 6.313 | 0.003 ** | 12.032 | 0.000 *** | |
| C × P | 2 | 9.122 | 0.000 *** | 2.171 | 0.123 | |
| M × C × P | 2 | 0.679 | 0.511 | 8.623 | 0.001 ** | |
| P accumulation | M | 1 | 70.742 | 0.000 *** | 80.432 | 0.000 *** |
| C | 1 | 3.485 | 0.067 | 70.320 | 0.000 *** | |
| P | 2 | 61.702 | 0.000 *** | 6.248 | 0.003 ** | |
| M × C | 1 | 6.470 | 0.014 * | 1.294 | 0.260 | |
| M × P | 2 | 11.663 | 0.000 *** | 8.267 | 0.001 ** | |
| C × P | 2 | 24.941 | 0.000 *** | 10.007 | 0.000 *** | |
| M × C × P | 2 | 9.075 | 0.000 *** | 1.461 | 0.240 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Shen, K.; Xia, T.; He, Y.; Guo, Y.; Wu, B.; Han, X.; Yan, J.; Jiao, M. Invasive and Native Plants Differentially Respond to Exogenous Phosphorus Addition in Root Growth and Nutrition Regulated by Arbuscular Mycorrhizal Fungi. Plants 2023, 12, 2195. https://doi.org/10.3390/plants12112195
Yang X, Shen K, Xia T, He Y, Guo Y, Wu B, Han X, Yan J, Jiao M. Invasive and Native Plants Differentially Respond to Exogenous Phosphorus Addition in Root Growth and Nutrition Regulated by Arbuscular Mycorrhizal Fungi. Plants. 2023; 12(11):2195. https://doi.org/10.3390/plants12112195
Chicago/Turabian StyleYang, Xionggui, Kaiping Shen, Tingting Xia, Yuejun He, Yun Guo, Bangli Wu, Xu Han, Jiawei Yan, and Min Jiao. 2023. "Invasive and Native Plants Differentially Respond to Exogenous Phosphorus Addition in Root Growth and Nutrition Regulated by Arbuscular Mycorrhizal Fungi" Plants 12, no. 11: 2195. https://doi.org/10.3390/plants12112195