
How Scarification, GA3 and Graphene Oxide Influence the In Vitro Establishment and Development of Strelitzia
 , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
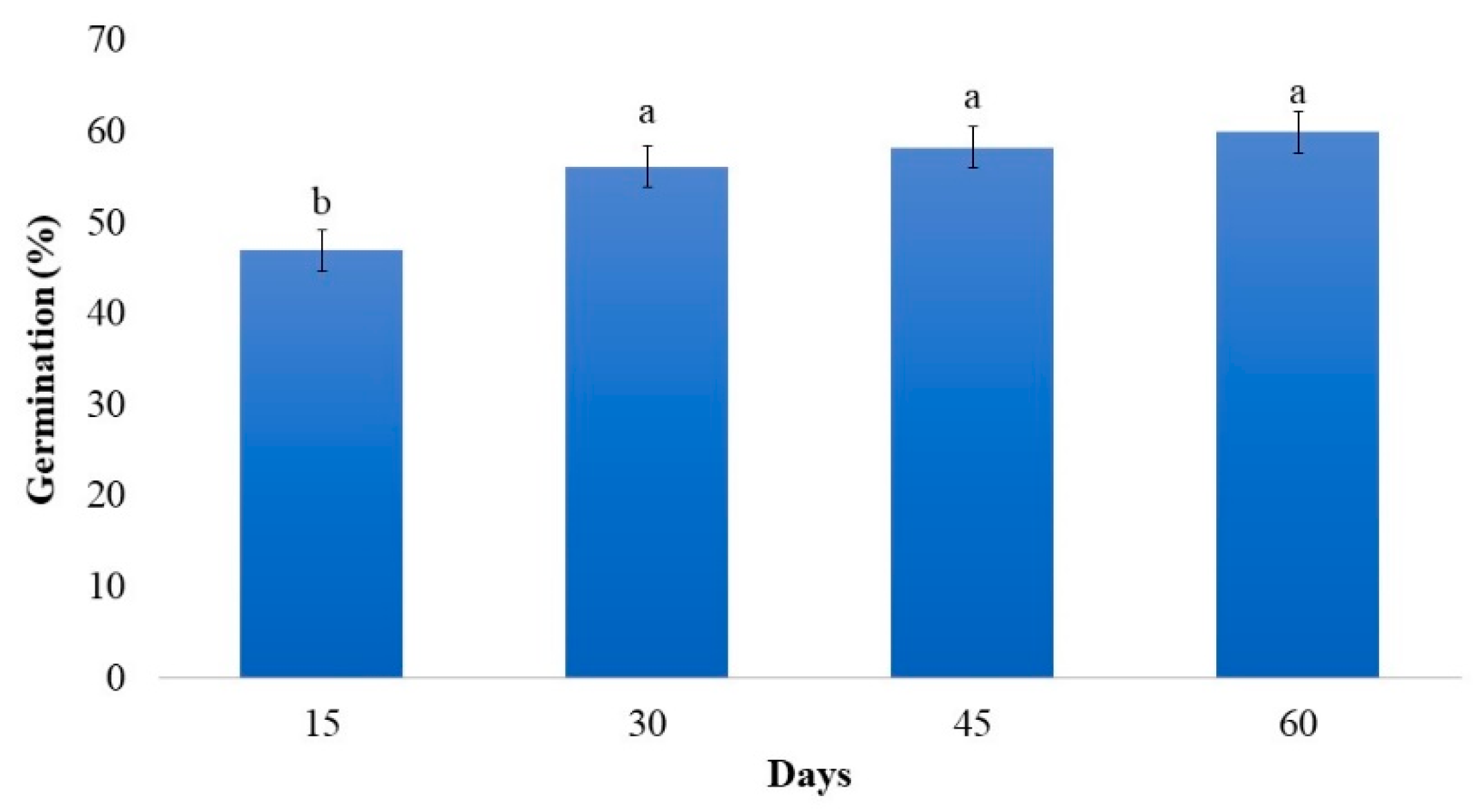
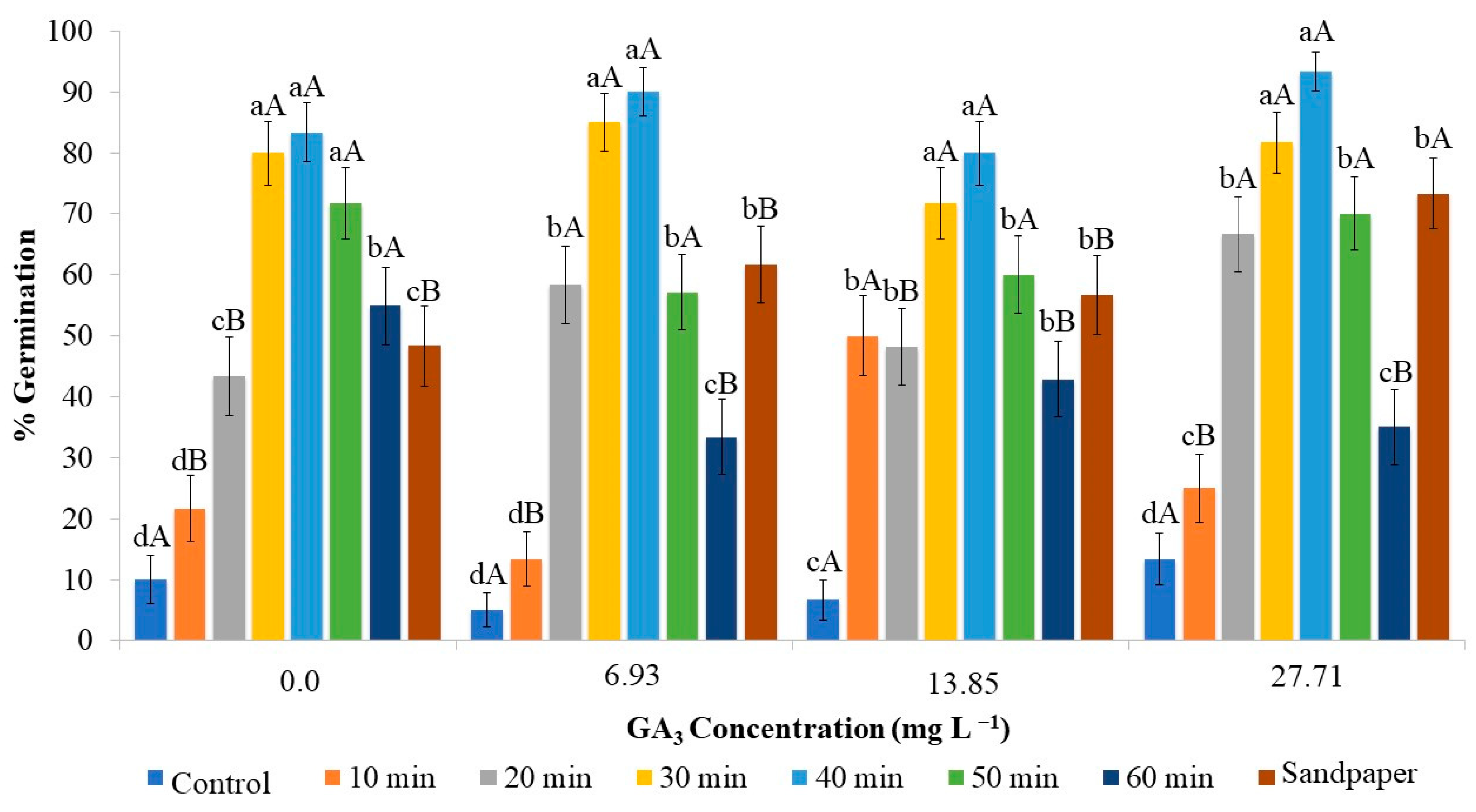
2.1. Effect of Gibberellin Associated with Scarification on In Vitro Germination of Strelitzia Seeds
2.2. Analysis of Images of Strelitzia Seeds Subjected to Chemical and Physical Scarification
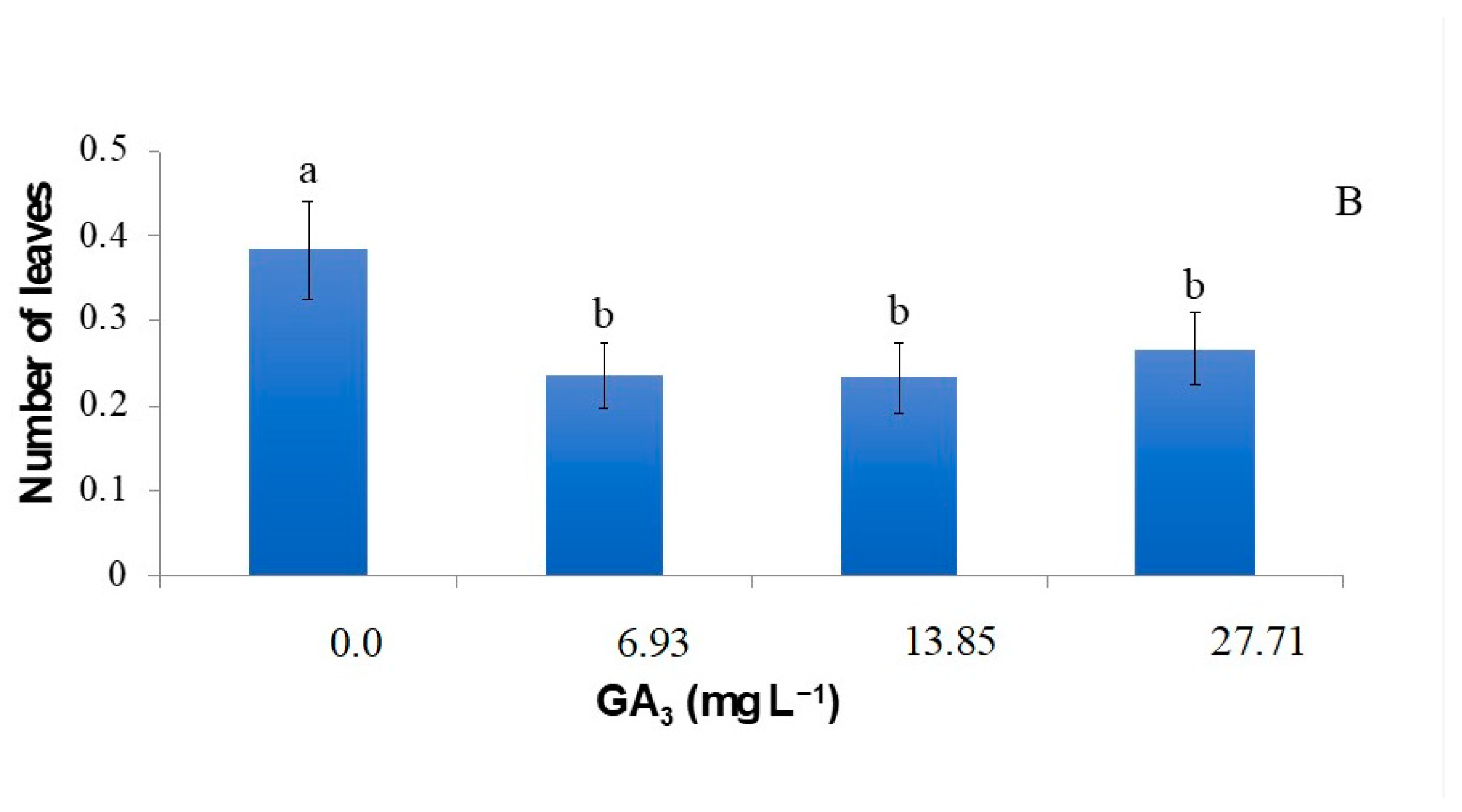
2.3. Effect of Graphene Oxide on the In Vitro Growth of Strelitzia Seedlings
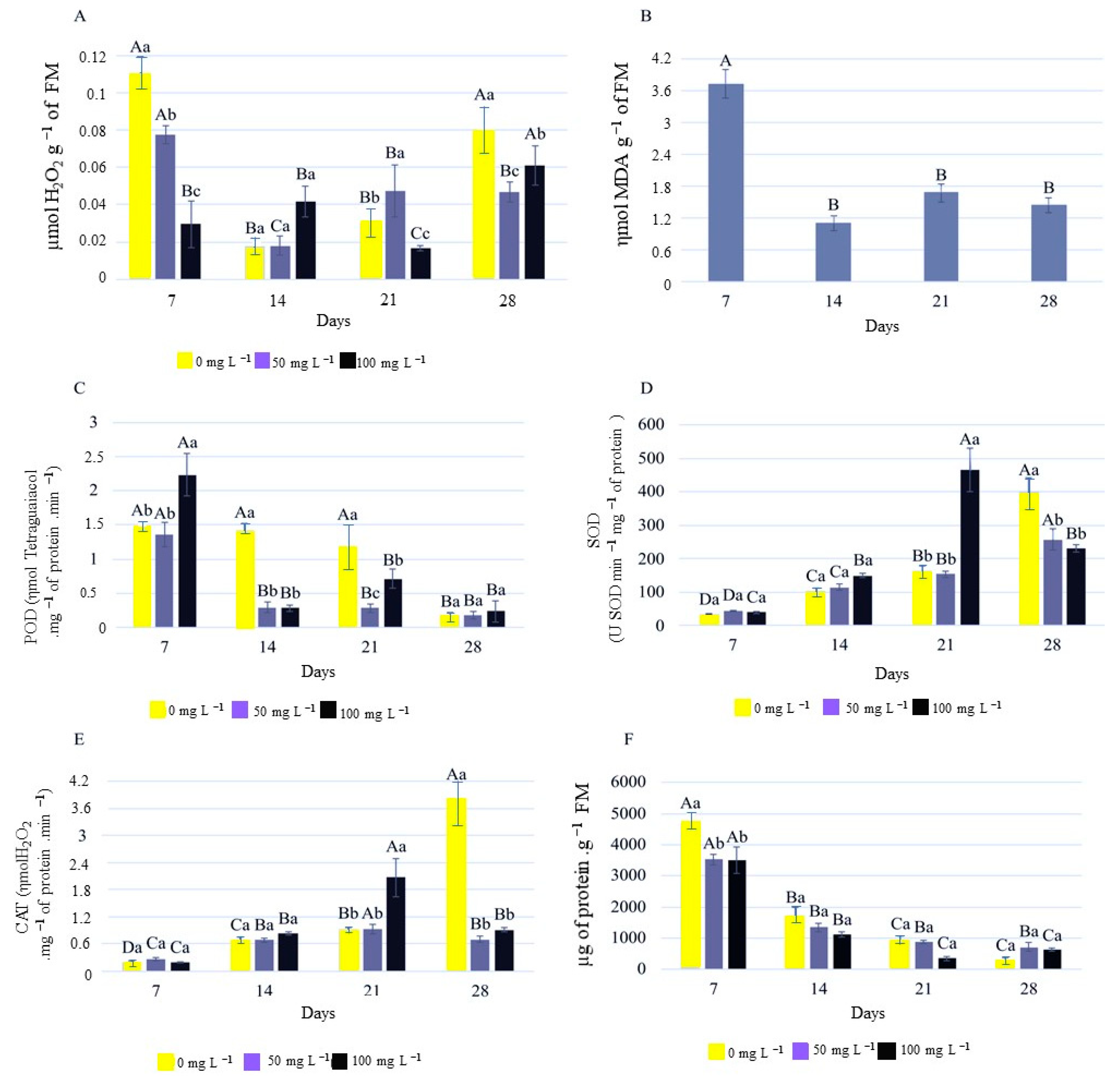
2.4. Biochemical Analysis
3. Material and Methods
3.1. Effect of Gibberellin Associated with Scarification on In Vitro Germination of Strelitzia Seeds
3.1.1. Effect of Gibberellin Associated with Seed Scarification on In Vitro Germination
3.1.2. Characterization of Seed Integrity after the Process of Chemical and Physical Scarification
3.2. Effect of Graphene Oxide on In Vitro Culture
3.2.1. Biochemical Analysis
3.2.2. Quantification of Hydrogen Peroxide (H2O2) and Lipid Peroxidation
3.2.3. Antioxidant Enzyme Activity: Catalase (CAT), Peroxidase (POD), and Superoxide Dismutase (SOD)
3.3. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paiva, P.D.O.; Almeida, E.F.A. Produção de Flores de Corte; UFLA: Lavras, Brazil, 2012; 678p. [Google Scholar]
- Paiva, P.D.O.; Pasqual, M.; Paiva, R. Controle de oxidação no cultivo in vitro de embriões de estrelícia (Strelitzia reginae). Rev. Bras. Hortic. Ornam. 2007, 13, 107–112. [Google Scholar] [CrossRef]
- Taniguchi, M.; Paiva, P.D.O.; Silva, D.P.C.; Nery, F.C.; Paiva, R.; Reis, M.V. Morpho-anatomical and biochemical characterization of Strelitzia reginae seeds and embryos. Ornam. Hortic. 2022, 28, 314–322. [Google Scholar] [CrossRef]
- Barbosa, J.G.; Alvarenga, E.M.; Dias, D.C.F.S.; Vieira, A.N. Efeito da escarificação ácida e de diferentes temperaturas na qualidade fisiológica de sementes de Strelitzia reginae. Rev. Bras. Sementes 2005, 27, 71–77. [Google Scholar] [CrossRef]
- Paiva, P.D.O.; Paiva, R.; Pasqual, M.; Paiva, L.V. Estabelecimento in vitro de estrelícia (Strelitzia reginae Banks.). Ciência Agrotecnologia 2004, 28, 1031–1037. [Google Scholar] [CrossRef]
- Marcos Filho, J. Fisiologia de Sementes de Plantas Cultivadas; FEALQ: Piracicaba, Brazil, 2005; 495p. [Google Scholar]
- Carvalho, F.J.; Carvalho, M.; Cunha, J.A. Dormancy breaking methods for bird of paradise seeds. Rev. Magistra 2020, 31, 799–804. [Google Scholar]
- Garcia, A.S.; Branquinho, E.G.A.; Menuchi, A.C.T.P.; Erlacher, K.C.; Domingues, M.C.S. Efeito de reguladores vegetais na germinação e desenvolvimento da Strelitzia reginae. Thesis 2006, 5, 161–176. [Google Scholar]
- Kumar, S.S.; Sane, A.; Reddy, M.L.; Sahijram, L.; Rao, A.V.D.D. Effect of growth regulators on seed germination of bird of paradise (Strelitzia reginae L.). Bull. Environ. Pharmacol. Life Sci. 2018, 7, 101–104. [Google Scholar]
- Figueiredo, J.R.M.; Paiva, P.D.O.; Silva, D.P.C.; Paiva, R.; Souza, R.R.; Reis, M.V. Effectiveness of high cytokinin levels on Strelitzia micropropagation. Acta Hortic. 2021, 1, 35–40. [Google Scholar] [CrossRef]
- Figueiredo, J.R.M.; Paiva, P.D.O.; Silva, D.P.C.; Paiva, R.; Souza, R.R.; Reis, M.V. Temperature and GA3 on ROS and cytogenetic stability during in vitro cultivation of strelitzia zygotic embryos. Ciencia Agrotecnologia 2021, 45, e020220. [Google Scholar] [CrossRef]
- Figueiredo, J.R.M.; Paiva, P.D.O.; Silva, D.P.C.; Paiva, R.; Mesquita, R.; Souza, R.R.; Reis, M.V. Anatomical changes and cytogenetic stability in bird of paradise plants after zygotic embryo cryopreservation by desiccation method. Vitr. Cell. Dev. Biol.-Plant 2021, 54, 272–280. [Google Scholar] [CrossRef]
- Bezerra, G.A.; Gabriel, A.V.M.D.; Mariano, E.D.; Cardoso, J.C. In vitro culture and greenhouse acclimatization of Oncidium varicosum (Orchidaceae) with microorganisms isolated from its roots. Ornam. Hortic. 2019, 25, 407–416. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal, 6th ed.; Artmed: Porto Alegre, Brazil, 2017. [Google Scholar]
- Shang, Y.; Hasan, M.K.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of nanotechnology in plant growth and crop protection: A review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef]
- Park, S.; Choi, K.S.; Kim, S.; Gwon, Y.; Kim, J. Graphene oxide-assisted promotion of plant growth and stability. Nanomaterials 2020, 10, 758. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xu, F.; Liu, Q.; Chen, M.; Liu, X.; Wang, Y.; Sun, Y.; Zhang, L. Nanomaterials and plants: Positive effects, toxicity and the remediation of metal and metalloid pollution in soil. Sci. Total Environ. 2019, 662, 414–421. [Google Scholar] [CrossRef]
- Dideikin, A.T.; Vul, A.Y. Graphene oxide and derivatives: The place in graphene family. Front. Phys. 2019, 6, 149. [Google Scholar] [CrossRef]
- He, Y.; Hu, R.; Zhong, Y.; Zhao, X.; Chen, Q.; Zhu, H. Graphene oxide as a water transporter promoting germination of plants in soil. Nano Res. 2018, 11, 1928–1937. [Google Scholar] [CrossRef]
- Gao, C.; Zhang, G.; Liu, Y. Effects of graphene oxide exposure on germination and seedling growth of rice and wheat. Hans J. Soil Sci. 2019, 07, 251–261. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, J.; Li, C.; Guo, Q.; Hou, X.; Zhao, C.; Wang, Y.; Chen, C.; Wang, Q. Effects of graphene oxide on the growth and photosynthesis of the emergent plant Iris pseudacorus. Plants 2023, 12, 1738. [Google Scholar] [CrossRef]
- Li, F.; Sun, C.; Li, X.; Yu, X.; Luo, C.; Shen, Y.; Qu, S. The effect of graphene oxide on adventitious root formation and growth in apple. Plant Physiol. Biochem. 2018, 129, 122–129. [Google Scholar] [CrossRef]
- Gao, M.; Xu, Y.; Chang, X.; Dong, Y.; Song, Z. Effects of foliar application of graphene oxide on cadmium uptake by lettuce. J. Hazard. Mater. 2020, 398, 122859. [Google Scholar] [CrossRef]
- Yamburov, M.S.; Astafurova, T.P.; Zharnakhova, E.Y.; Romanova, S.B.; Smolina, V.M.; Khotskova, L.V. Reproductive biology of Strelitzia nicolai and Strelitzia reginae in the conditions of a greenhouse. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 309–316. [Google Scholar]
- Martín, I.; Guerrero, M. Effect of sulphuric acid scarification on seed accessions of cluster clover (Trifolium glomeratum) stored in a genebank. Seed Sci. Technol. 2014, 42, 293–299. [Google Scholar] [CrossRef]
- Afonso, M.V.; Paranhos, J.T.; Tabaldi, L.A.; Soriani, H.H. Germinação in vitro de sementes e parâmetros morfofisiológicos de microestacas de Tabernaemontana catharinensis A. DC. Iheringia Série Botânica 2018, 73, 39–45. [Google Scholar] [CrossRef]
- Silva, B.M.S.; Silva, C.O.; Môro, F.V.; Vieira, R.D. Seed anatomy and water uptake and their relation to seed dormancy of Ormosia paraensis Ducke. J. Seed Sci. 2018, 40, 237–245. [Google Scholar] [CrossRef]
- Brancalion, P.H.S.; Mondo, V.H.V.; Novembre, A.D.L.C. Escarificação química para a superação da dormência de sementes de saguaraji-vermelho (Colubrina glandulosa Perk.—Rhamnaceae). Rev. Árvore 2011, 35, 119–124. [Google Scholar] [CrossRef]
- Moura, L.C.; Titon, M.; Fernandes, J.S.C.; Santana, R.C. Germinação in vitro e aclimatação de plântulas de sucupira-preta (Bowdichia virgilioides Kunth.). Biosci. J. 2014, 30, 678–687. [Google Scholar]
- Paiva, P.D.O.; Cardoso, M.G.; Pasqual, M.; Paiva, R. Identificação de compostos liberados no meio de cultura pelo processo de oxidação em cultivo in vitro de estrelícia. Ciência Agrotecnologia 2000, 24, 50–55. [Google Scholar]
- Cho, J.S.; Lee, C.H. Effect of germination and water absorption on scarification and stratification of kousa dogwood seed. Hortic. Environ. Biotech. 2018, 59, 335–344. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, W.; Fu, X.; Liu, A.; Cao, J.; Liu, J. Graphene oxide, a novel nanomaterial as soil water retention agent, dramatically enhances drought stress tolerance in soybean plants. Front. Plant Sci. 2022, 13, 810905. [Google Scholar] [CrossRef]
- He, Y.; Wei, H.M.; Liu, S.J.; Xu, Y.C.; Zhu, Z.Y.; Yan, H.J.; Li, X.; Tian, Z.H. Growth response of Oryza sativa seedlings to graphene oxide and its variability among genotypes. Biol. Plant. 2021, 65, 39–46. [Google Scholar] [CrossRef]
- Chen, L.; Yang, S.; Liu, Y.; Mo, M.; Guan, X.; Huang, L.; Sun, C.; Yang, S.-T.; Chang, X.-L. Toxicity of graphene oxide to naked oats (Avena sativa L.) in hydroponic and soil cultures. RSC Adv. 2018, 8, 15336–15343. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Huang, J.; Lin, X.; Wang, Y.; Cao, C.; Qian, X. Performance of constructed wetlands with different water level for treating graphene oxide wastewater: Characteristics of plants and microorganisms. J. Environ. Manag. 2023, 334, 117432. [Google Scholar] [CrossRef] [PubMed]
- Konieczny, R.; Banaś, A.K.; Surówka, E.; Michalec, Ż.; Miszalski, Z.; Libik-Konieczny, M. Pattern of antioxidant enzyme activities and hydrogen peroxide content during developmental stages of rhizogenesis from hypocotyl explants of Mesembryanthemum crystallinum L. Plant Cell Rep. 2014, 33, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.R.; Paiva, P.D.O.; Souza, A.R.; Silva, R.R.; Silva, D.P.C.; Reis, M.V.; Paiva, R. Morpho-anatomical changes and antioxidant enzyme activity during the acclimatization of Genipa americana. Acta Physiol. Plant. 2021, 43, 93. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, M.S.; Zulfiqar, F.; Alam, M.M.; Fujita, M. Regulation of ROS metabolism in plants under environmental stress: A Review of recent experimental evidence. Int. J. Mol. Sci. 2020, 18, 8695. [Google Scholar] [CrossRef]
- Bailly, C. The signaling role of ROS in the regulation of seed germination and dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef]
- Kondamudi, R.; Sumar, K.; Swamy, K.N.; Murthy, K.S.R. Role of antioxidant enzymes in acclimatization of Ceropegia spiralis Wt., C. pusilla Wt. and Arn. and C. juncea Roxb. Int. J. Adv. Res. Bot. 2017, 3, 19–27. [Google Scholar]
- Gonçalves, S.; Martins, N.; Romano, A. Physiological traits and oxidative stress markers during acclimatization of micropropagated plants from two endangered Plantago species: P. algarbiensis Samp. and P. almogravensis Franco. Vitr. Cell. Dev. Biol.-Plant 2017, 53, 249–255. [Google Scholar] [CrossRef]
- Sairam, R.K.; Deshmukh, P.S.; Saxena, D.C. Role of antioxidante systems in wheat genotype tolerance to water stress. Biol. Plant. 1998, 41, 387–394. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Guo, X.; Zhao, J.; Wang, R.; Zhang, H.; Xing, B.; Naeem, M.; Yao, T.; Li, R.; Xu, R.; Zhang, Z.; et al. Effects of graphene oxide on tomato growth in different stages. Plant Physiol. Biochem. 2021, 162, 447–455. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1978; pp. 302–310. [Google Scholar]
- Biemelt, S.; Keetman, U.; Albrecht, G. Re-aeration following hypoxia or anoxia leads to activation of the antioxidative defense system in roots of wheat seedlings. Plant Physiol. 1998, 116, 651–658. [Google Scholar] [CrossRef]
- Havir, E.A.; Mchale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.-C.; Kao, C.H. Enhanced peroxidase activity in rice leaves in response to excess iron, copper and zinc. Plant Sci. 2000, 158, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Ferreira, D.F. Sisvar: A guide for its bootstrap procedures in multiple comparisons. Ciência Agrotecnologia 2014, 38, 109–112. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Color | Scarification | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | 10 min | 20 min | 30 min | 40 min | 50 min | 60 min | Sand | |
| Black | 88.72 | 98.89 | 98.88 | 98.41 | 95.24 | 76.51 | 59.59 | 83.42 |
| Sky blue | 1.12 | 0.07 | 0 | 0 | 0 | 0 | 0 | 1.32 |
| Gray (light) | 0 | 0 | 0 | 0 | 0 | 0.05 | 0.13 | 2.56 |
| Red | 0 | 0 | 0.48 | 0.47 | 0.58 | 0.96 | 1.86 | 0 |
| Orange | 0.74 | 0.26 | 0.15 | 0.78 | 2.23 | 17.32 | 30.28 | 1.38 |
| Pink | 0 | 0 | 0.16 | 0.05 | 0 | 0 | 0 | 0 |
| Cyan | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paiva, P.D.d.O.; Silva, D.P.C.d.; Silva, B.R.d.; Sousa, I.P.d.; Paiva, R.; Reis, M.V.d. How Scarification, GA3 and Graphene Oxide Influence the In Vitro Establishment and Development of Strelitzia. Plants 2023, 12, 2142. https://doi.org/10.3390/plants12112142
Paiva PDdO, Silva DPCd, Silva BRd, Sousa IPd, Paiva R, Reis MVd. How Scarification, GA3 and Graphene Oxide Influence the In Vitro Establishment and Development of Strelitzia. Plants. 2023; 12(11):2142. https://doi.org/10.3390/plants12112142
Chicago/Turabian StylePaiva, Patrícia Duarte de Oliveira, Diogo Pedrosa Correa da Silva, Bruna Raphaella da Silva, Israela Pimenta de Sousa, Renato Paiva, and Michele Valquíria dos Reis. 2023. "How Scarification, GA3 and Graphene Oxide Influence the In Vitro Establishment and Development of Strelitzia" Plants 12, no. 11: 2142. https://doi.org/10.3390/plants12112142