
Exogenous Melatonin Regulates Physiological Responses and Active Ingredient Levels in Polygonum cuspidatum under Drought Stress

 ,
,  and
and 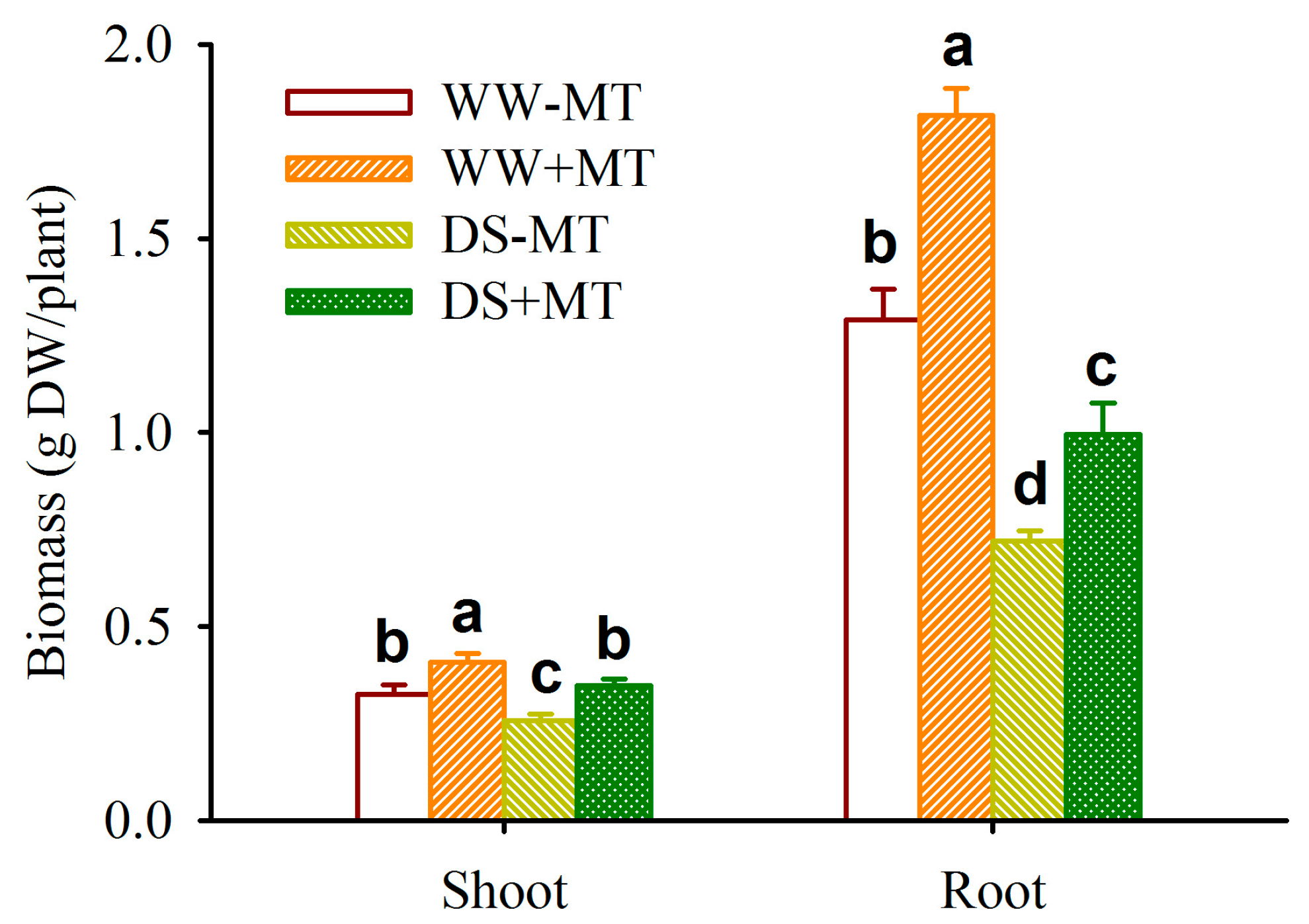{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Response of Plant Biomass
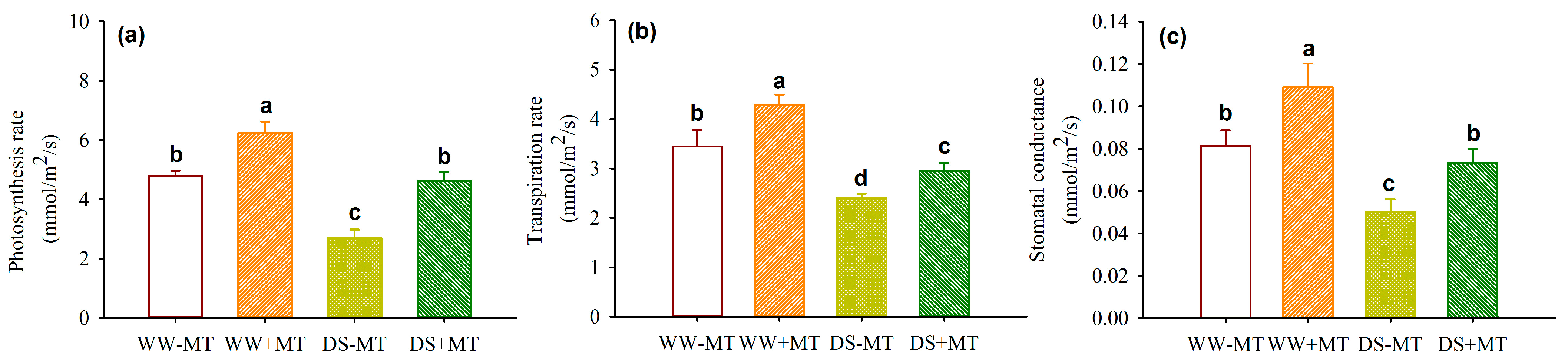
2.2. Response of Leaf Gas Exchange and Water Potential
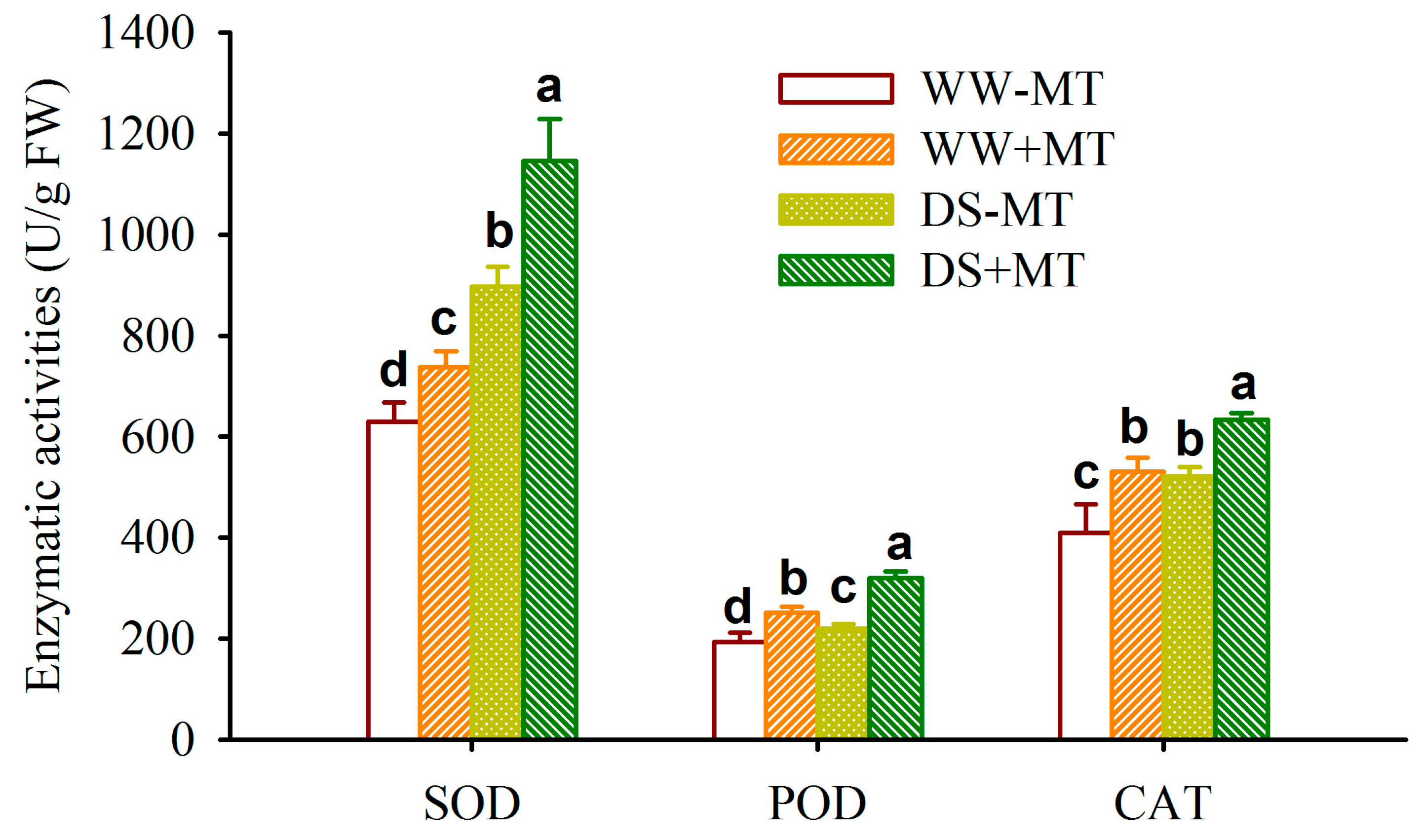
2.3. Response of Root Antioxidant Enzyme Activities
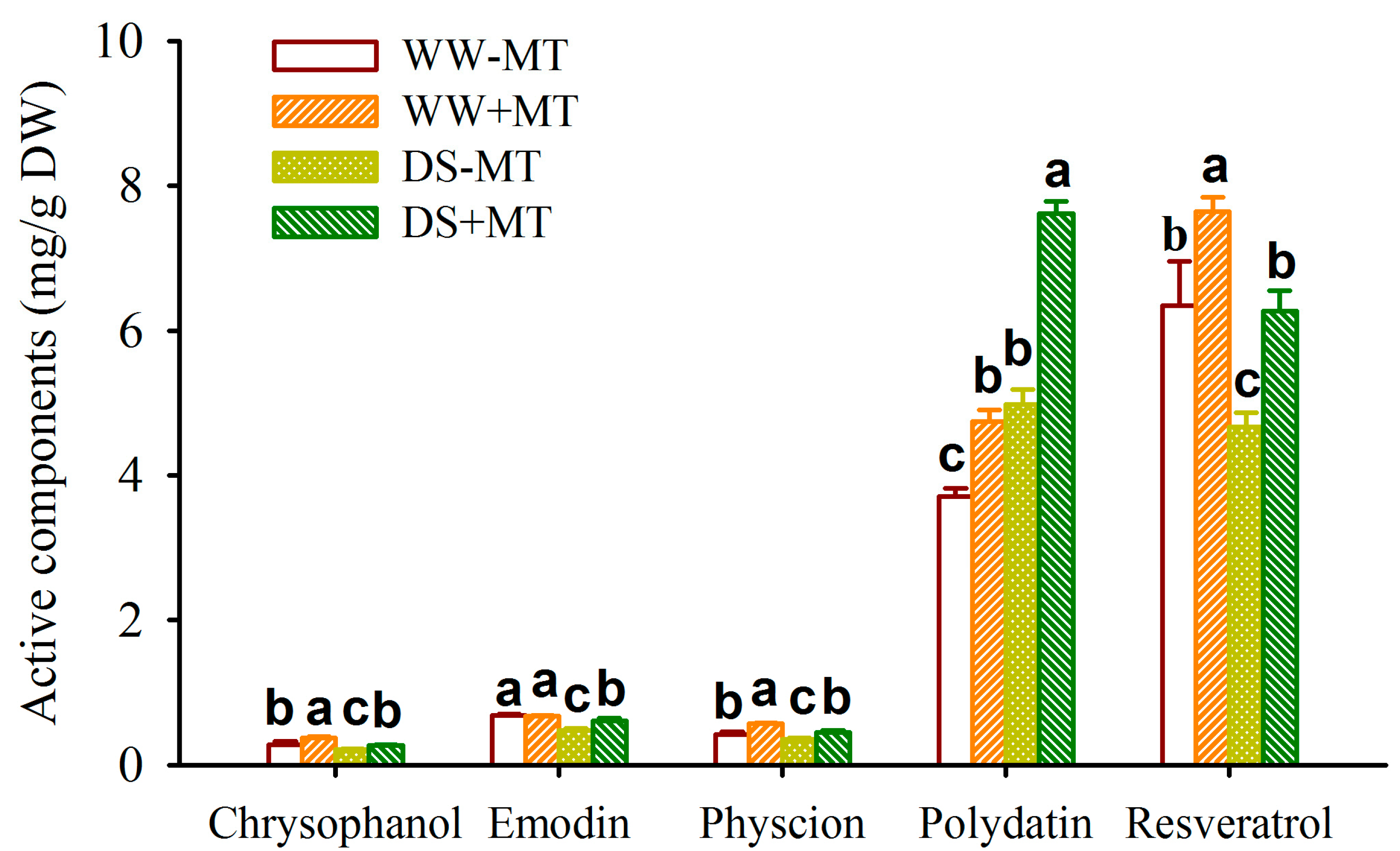
2.4. Response of Root Active Substance Concentrations
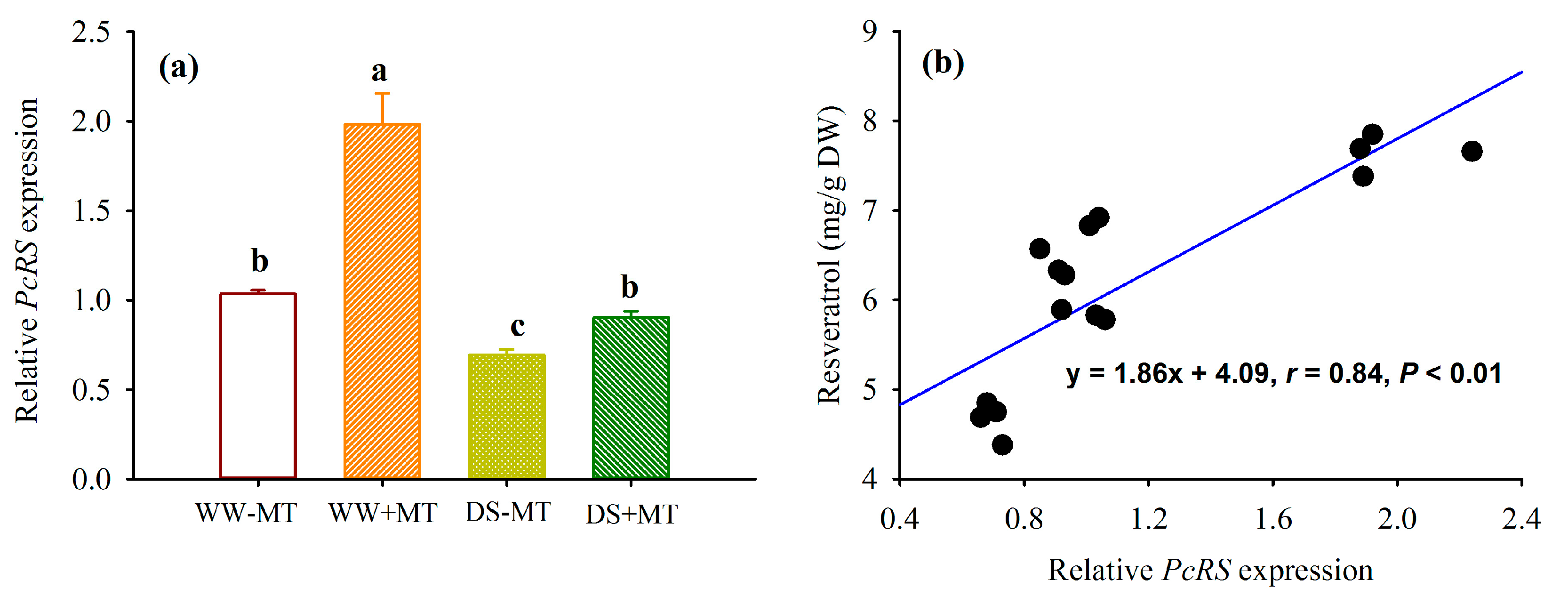
2.5. Response of Root PcRS Gene Expression
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Culture and Treatment Application
5.2. Experimental Design
5.3. Determinations of Plant Biomass, Leaf Gas Exchange, and Leaf Water Potential
5.4. Determinations of Antioxidant Enzyme Activities in Leaves
5.5. Determinations of Active Components in Roots
5.6. Determinations of Gene Expressions in Roots
5.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, W.H.; Bi, J.W.; Zhao, Y.J.; Qiu, S.Y.; Madalin, P.; Oliver, B.; Christina, R.; Ju, R.T.; Wu, J.H.; Liao, Z.Y.; et al. Latitudinal variation and influence factors of two secondary metabolites in Reynoutria japonica leaves. J. Fudan Univ. (Nat. Sci.) 2022, 61, 249–259. [Google Scholar]
- Du, K.X.; Jiang, Y.B.; Chen, J.; Mu, W.L. Study on antioxidant effect of Polygonum cuspidatum and various plant extracts. Oral Care Ind. 2022, 32, 46–49. [Google Scholar]
- Chen, J. Preliminary Study on Key Breeding Techniques of Polygonum cuspidatum. Master’s Thesis, Chengdu University of TCM, Chengdu, China, 2012; pp. 1–47. [Google Scholar]
- Sun, R.-T.; Zhang, Z.-Z..; Liu, M.-Y.; Feng, X.-C.; Zhou, N.; Feng, H.-D.; Hashem, A.; Abd_Allah, E.F.; Harsonowati, W.; Wu, Q.-S. Arbuscular mycorrhizal fungi and phosphorus supply accelerate main medicinal component production of Polygonum cuspidatum. Front. Microbiol. 2022, 13, 1006140. [Google Scholar] [CrossRef]
- Sun, R.-T.; Zhang, Z.-Z.; Feng, X.-C.; Zhou, N.; Feng, H.-D.; Liu, Y.-M.; Harsonowati, W.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.-S. Endophytic fungi accelerate leaf physiological activity and resveratrol accumulation in Polygonum cuspidatum by up-regulating expression of associated genes. Agronomy 2022, 12, 1220. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmad, W.; Reiter, R.J.; Niu, M.; Hameed, S. Melatonin: Current status and future perspectives in plant science. Front. Plant Sci. 2016, 6, 1230. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2012, 63, 577–597. [Google Scholar] [CrossRef]
- Li, C.; Tan, D.X.; Liang, D.; Chang, C.; Jia, D.F.; Ma, F.W. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging and stomatal behavior in two Malus species under drought stress. J. Exp. Bot. 2015, 66, 669–680. [Google Scholar] [CrossRef]
- Sadak, M.S.; Abdalla, A.M.; Abd Elhamid, E.M.; Ezzo, M.I. Role of melatonin in improving growth, yield quantity and quality of Moringa oleifera L. plant under drought stress. Bull. Natl. Res. Cent. 2020, 44, 18. [Google Scholar] [CrossRef]
- Antoniou, C.; Chatzimichail, G.; Xenofontos, R.; Pavlou, J.J.; Panagiotou, E.; Christou, A.; Fotopoulos, V. Melatonin systemically ameliorates drought stress-induced damage in Medicago sativa plants by modulating nitro-oxidative homeostasis and proline metabolism. J. Pineal Res. 2017, 62, e12401. [Google Scholar] [CrossRef]
- Liang, B.; Ma, C.; Zhang, Z.; Wei, Z.; Gao, T.; Zhao, Q.; Ma, F.; Li, C. Long-term exogenous application of melatonin improves nutrient uptake fluxes in apple plants under moderate drought stress. Environ. Exp. Bot. 2018, 155, 650–661. [Google Scholar] [CrossRef]
- Cao, L.; Jin, X.; Zhang, Y.; Zhang, M.; Wang, Y. Transcriptomic and metabolomic profiling of melatonin treated soybean (Glycine max L.) under drought stress during grain filling period through regulation of secondary metabolite biosynthesis pathways. PLoS ONE 2020, 15, e0239701. [Google Scholar] [CrossRef] [PubMed]
- Jafari, M.; Shahsavar, A. The effect of foliar application of melatonin on changes in secondary metabolite contents in two citrus species under drought stress conditions. Front. Plant Sci. 2021, 12, 692735. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Naz, S.; Altaf, M.M.; Khan, L.U.; Tiwari, R.K.; Lal, M.K.; Shahid, M.A.; Kumar, R.; et al. Melatonin improves drought stress tolerance of tomato by modulating plant growth, root architecture, photosynthesis, and antioxidant defense system. Antioxidants 2022, 11, 309. [Google Scholar] [CrossRef]
- Tomann, P.; Paus, R.; Millar, S.E.; Scheidereit, C.; Schmidt-Ullrich, R. LHX2 is a direct NF-κB target gene that promotes primary hair follicle placode down-growth. Development 2016, 143, 1512. [Google Scholar] [CrossRef]
- Hura, T.; Hura, K.; Grzesiak, M.; Rzepka, A. Effect of long-term drought stress on leaf gas exchange and fluorescence parameters in C3 and C4 plants. Acta Physiol. Plant. 2007, 29, 103–113. [Google Scholar] [CrossRef]
- Zhang, P.P.; Gao, Y.; Wang, X.L.; Liu, Y.R.; Liang, X.L.; Zhang, X. Effects of melatonin on seedlings morphology and physiological characteristics of lettuce under PEG stress. North Hortic. 2022, 10, 1–8. [Google Scholar]
- Dai, L.; Li, J.; Harmens, H.; Zheng, X.; Zhang, C. Melatonin enhances drought resistance by regulating leaf stomatal behaviour, root growth and catalase activity in two contrasting rapeseed (Brassica napus L.) genotypes. Plant Physiol. Biochem. 2020, 149, 86–95. [Google Scholar] [CrossRef]
- Hu, H.; Wang, X.; Wu, Z.; Chen, M.; Chai, T.; Wang, H. Overexpression of the Polygonum cuspidatum PcDREB2A gene encoding a DRE-Binding transcription factor enhances the drought tolerance of transgenic Arabidopsis thaliana. J. Plant Biol. 2022, 65, 505–515. [Google Scholar] [CrossRef]
- Pieri, C.; Marra, M.; Moroni, F.; Recchioni, R.; Marcheselli, F. Melatonin: A peroxyl radical scavenger more effective than vitamin E. Life Sci. 1994, 55, 271–276. [Google Scholar] [CrossRef]
- Sun, R.-T.; Feng, X.-C.; Zhang, Z.-Z.; Zhou, N.; Feng, H.-D.; Liu, Y.-M.; Hashem, A.; Al-Arjani, A.-B.F.; Abd_Allah, E.F.; Wu, Q.-S. Root endophytic fungi regulate changes in sugar and medicinal compositions of Polygonum cuspidatum. Front. Plant Sci. 2022, 13, 818909. [Google Scholar] [CrossRef]
- Xie, W.; Hao, Z.; Zhou, X.; Jiang, X.; Xu, L.; Wu, S.; Zhao, A.; Zhang, X.; Chen, B. Arbuscular mycorrhiza facilitates the accumulation of glycyrrhizin and liquiritin in Glycyrrhiza uralensis under drought stress. Mycorrhiza 2018, 28, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Jayalakshmi, P.; Devika, P.T. Assessment of in vitro antioxidant activity study of polydatin. J. Pharmacogn. Phytochem. 2019, 8, 55–58. [Google Scholar]
- Pandi-Perumal, S.R.; Trakht, I.; Srinivasan, V.; Spence, D.W.; Maestroni, G.J.; Zisapel, N.; Cardinali, D.P. Physiological effects of melatonin: Role of melatonin receptors and signal transduction pathways. Prog. Neurobiol. 2008, 85, 335–353. [Google Scholar] [CrossRef] [PubMed]
- Kolář, J.; Macháčková, I. Melatonin in higher plants: Occurrence and possible functions. J. Pineal Res. 2005, 39, 333–341. [Google Scholar] [CrossRef]
- Liu, Z.; Zhuang, C.; Sheng, S.; Shao, L.; Zhao, W.; Zhao, S. Overexpression of a resveratrol synthase gene (PcRS) from Polygonum cuspidatum in transgenic Arabidopsis causes the accumulation of trans-piceid with antifungal activity. Plant Cell Rep. 2011, 30, 2027–2036. [Google Scholar] [CrossRef]
- Thomzik, J.E.; Stenzel, K.; Stocker, R.; Schreier, P.H.; Hain, R.; Stahl, D.J. Synthesis of a grapevine phytoalexin in transgenic tomatoes (Lycopersicon esculentum Mill.) conditions resistance against Phytophthora infestans. Physiol. Mol. Plant Pathol. 1997, 51, 265–278. [Google Scholar] [CrossRef]
- Durak, I. A methodological approach to superoxide dismutase (SOD) activity assay based on inhibition of nitroblue tetrazolium (NBT) reduction. Clin. Chim. Acta 1993, 214, 103–104. [Google Scholar] [CrossRef]
- Rong, Z.-Y.; Jiang, D.-J.; Cao, J.-L.; Hashem, A.; Abd_Allah, E.F.; Alsayed, M.F.; Harsonowati, W.; Wu, Q.-S. Endophytic fungus Serendipita indica accelerates ascorbate-glutathione cycle of white clover in response to water stress. Front. Microbiol. 2022, 13, 967851. [Google Scholar] [CrossRef]
- Ma, W.-Y.; Qin, Q.-Y.; Zou, Y.-N.; Kuča, K.; Giri, B.; Wu, Q.-S.; Hashem, A.; Al-Arjani, A.-B.F.; Almutairi, K.F.; Abd_Allah, E.F.; et al. Arbuscular mycorrhiza induces low oxidative burst in drought-stressed walnut through activating antioxidant defense systems and heat shock transcription factor expression. Front. Plant Sci. 2022, 13, 1089420. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, R.-J.; Ye, M.-Y.; Liu, Y.; Wu, Q.-S.; Abd_Allah, E.F.; Zhou, N. Exogenous Melatonin Regulates Physiological Responses and Active Ingredient Levels in Polygonum cuspidatum under Drought Stress. Plants 2023, 12, 2141. https://doi.org/10.3390/plants12112141
Shi R-J, Ye M-Y, Liu Y, Wu Q-S, Abd_Allah EF, Zhou N. Exogenous Melatonin Regulates Physiological Responses and Active Ingredient Levels in Polygonum cuspidatum under Drought Stress. Plants. 2023; 12(11):2141. https://doi.org/10.3390/plants12112141
Chicago/Turabian StyleShi, Ru-Jie, Ming-Yan Ye, Yue Liu, Qiang-Sheng Wu, Elsayed Fathi Abd_Allah, and Nong Zhou. 2023. "Exogenous Melatonin Regulates Physiological Responses and Active Ingredient Levels in Polygonum cuspidatum under Drought Stress" Plants 12, no. 11: 2141. https://doi.org/10.3390/plants12112141