
Biocontrol Potential of an Endophytic Pseudomonas poae Strain against the Grapevine Trunk Disease Pathogen Neofusicoccum luteum and Its Mechanism of Action
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
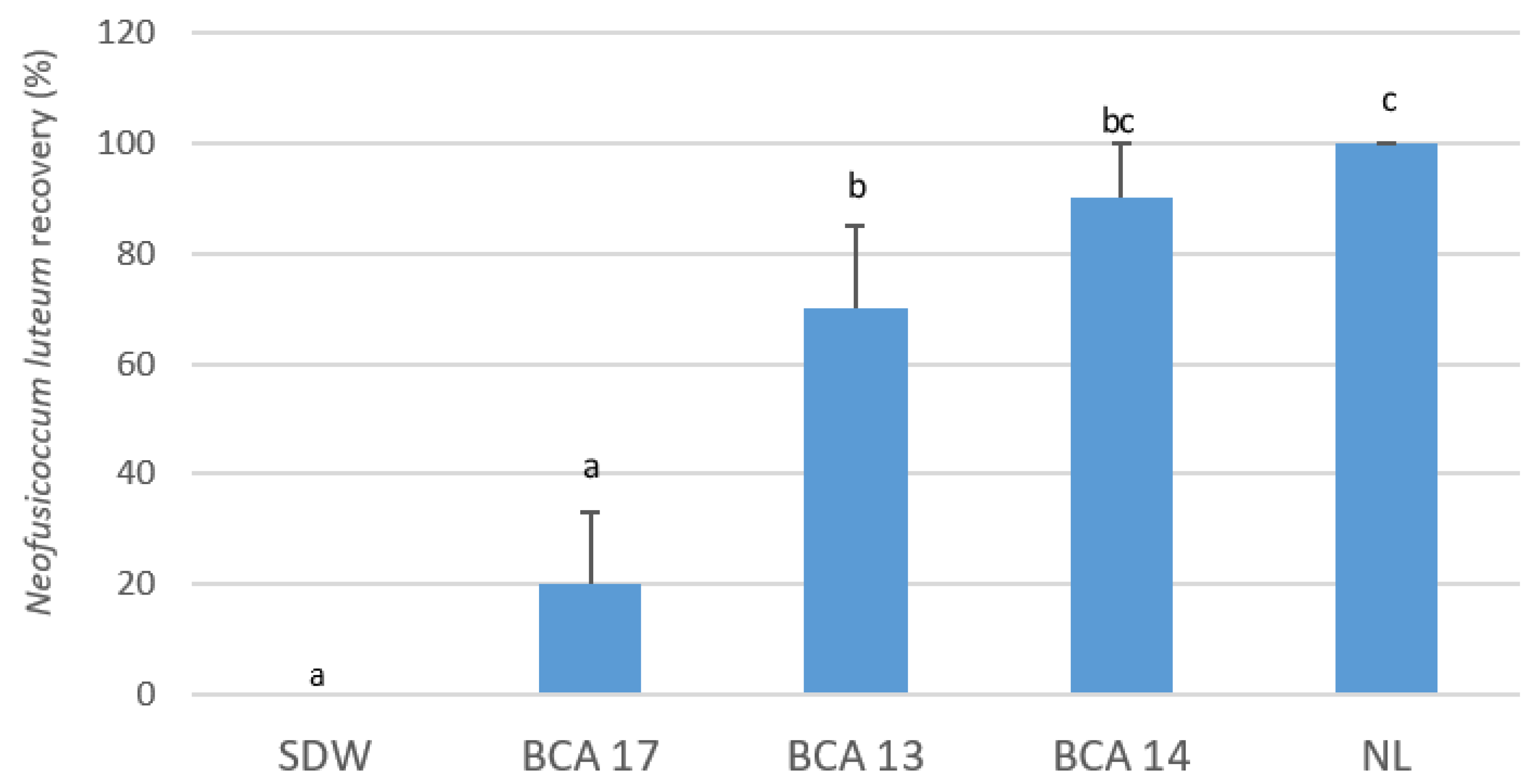
2.1. Suppression of Neofusicoccum luteum Infection in Planta by Pseudomonas Strains
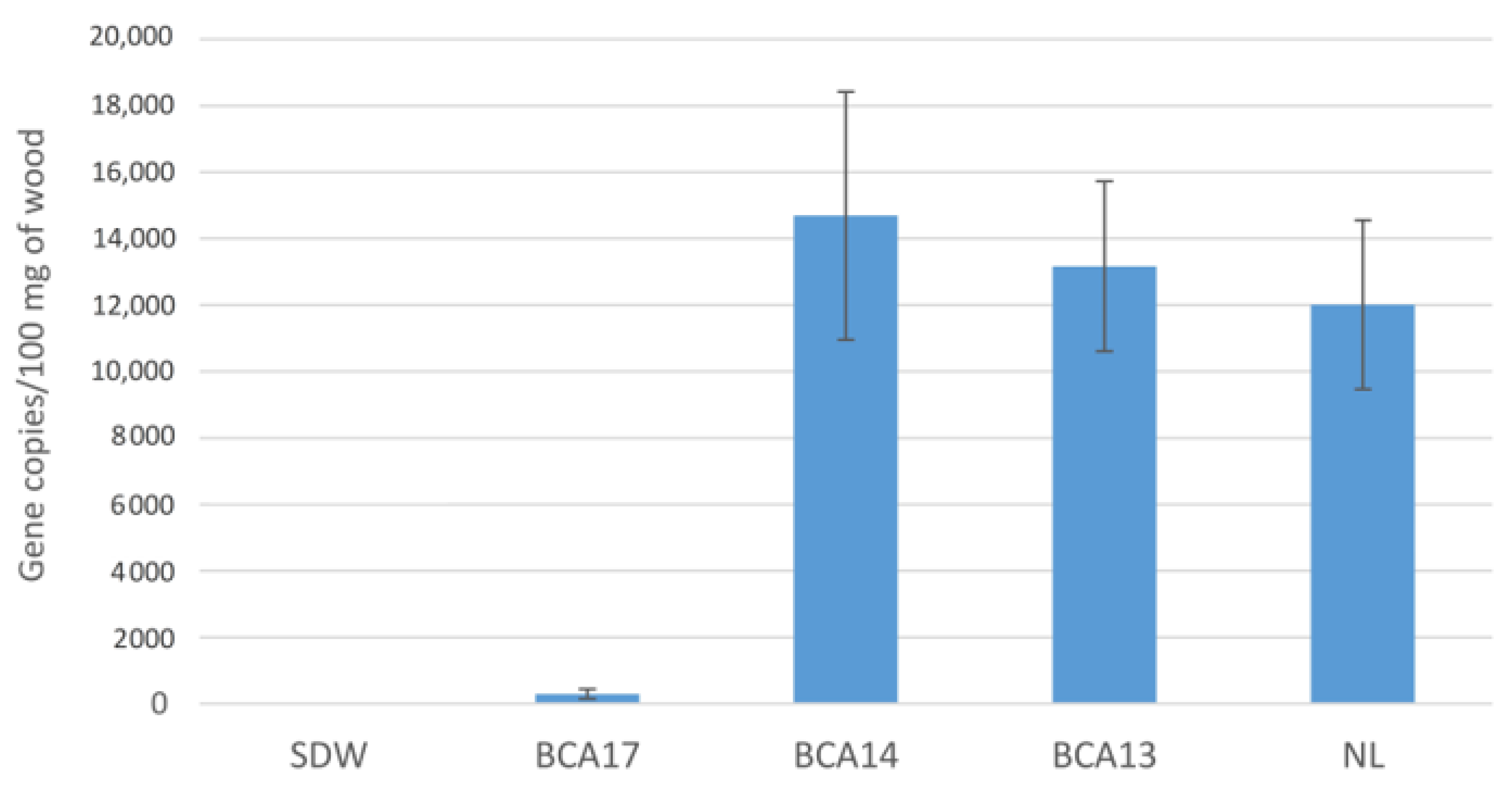
Assessment of N. luteum Infections by qPCR
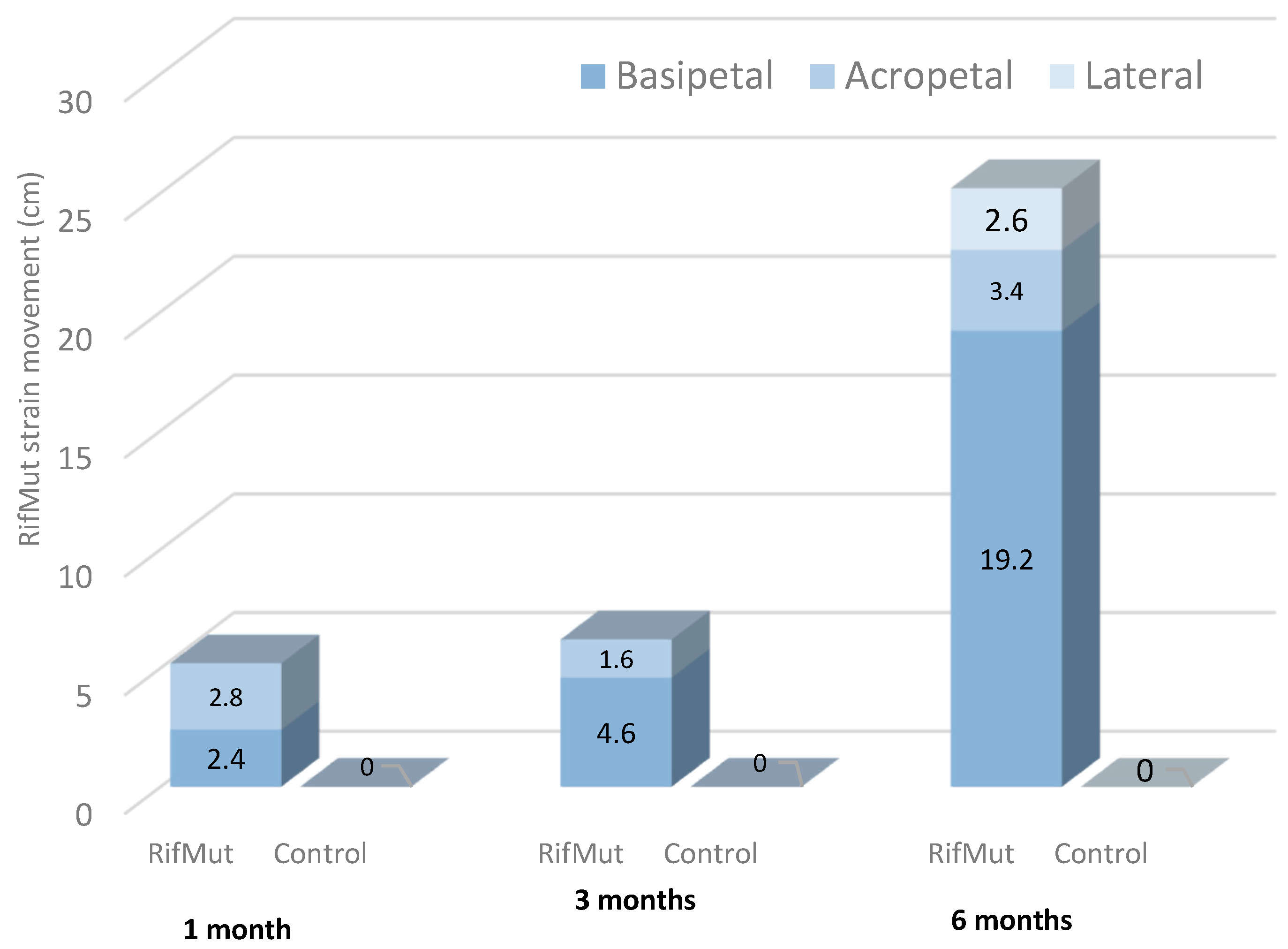
2.2. Pseudomonas Colonization and Establishment in Grapevine Tissues
2.2.1. Pre-Planting Application of Pseudomonas RifMut Strain to Canes
2.2.2. Post-Planting Application of Pseudomonas
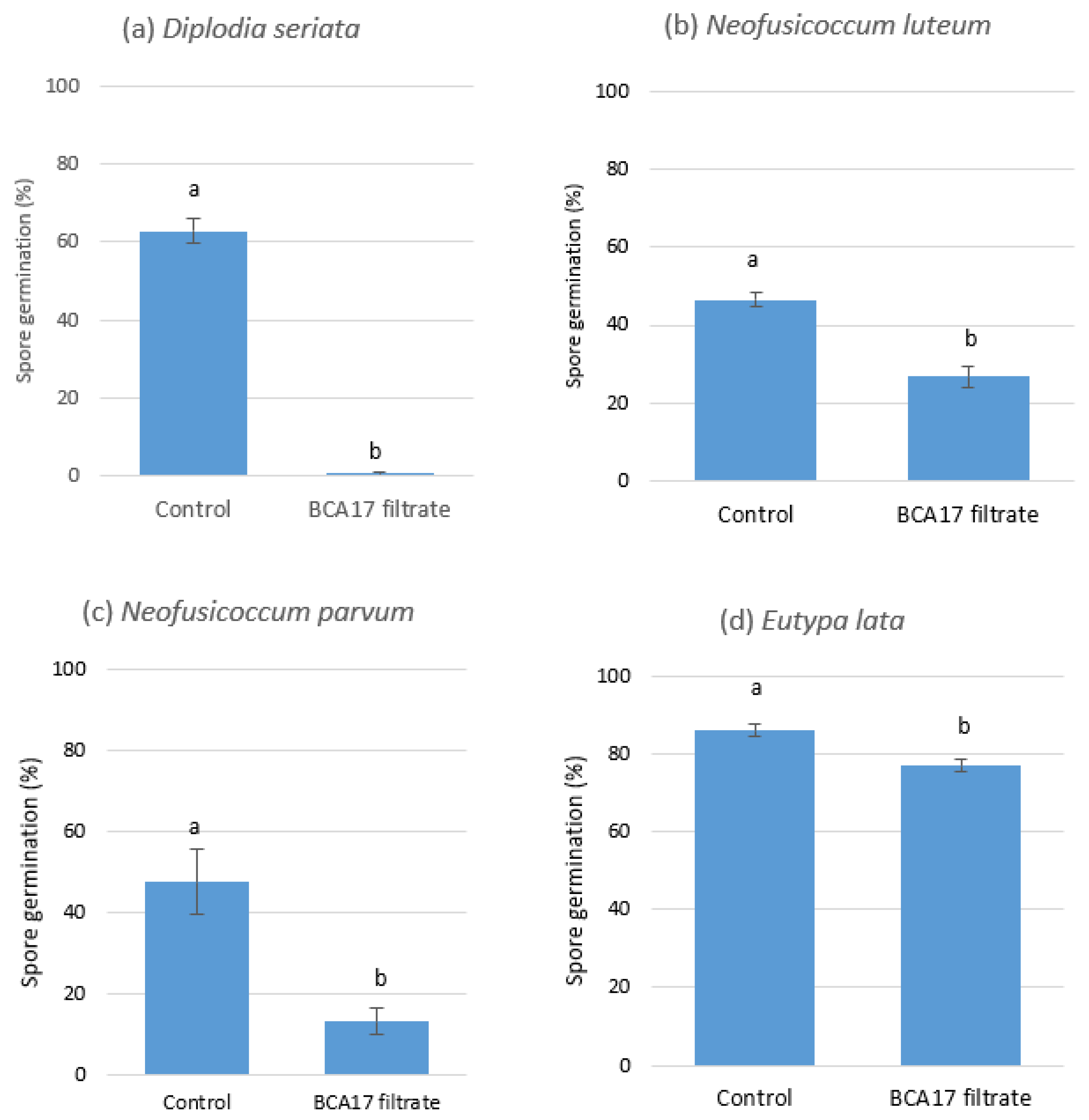
2.3. Bioactivity of P. poae BCA17 Culture Filtrate on Mycelial Growth and Spore Germination of GTD Pathogens
2.3.1. Effect of BCA17 Culture Filtrate on Mycelial Growth
2.3.2. Effect of BCA17 Culture Filtrate on Spore Germination
2.4. Identification of Secondary Metabolite Biosynthetic Gene Clusters
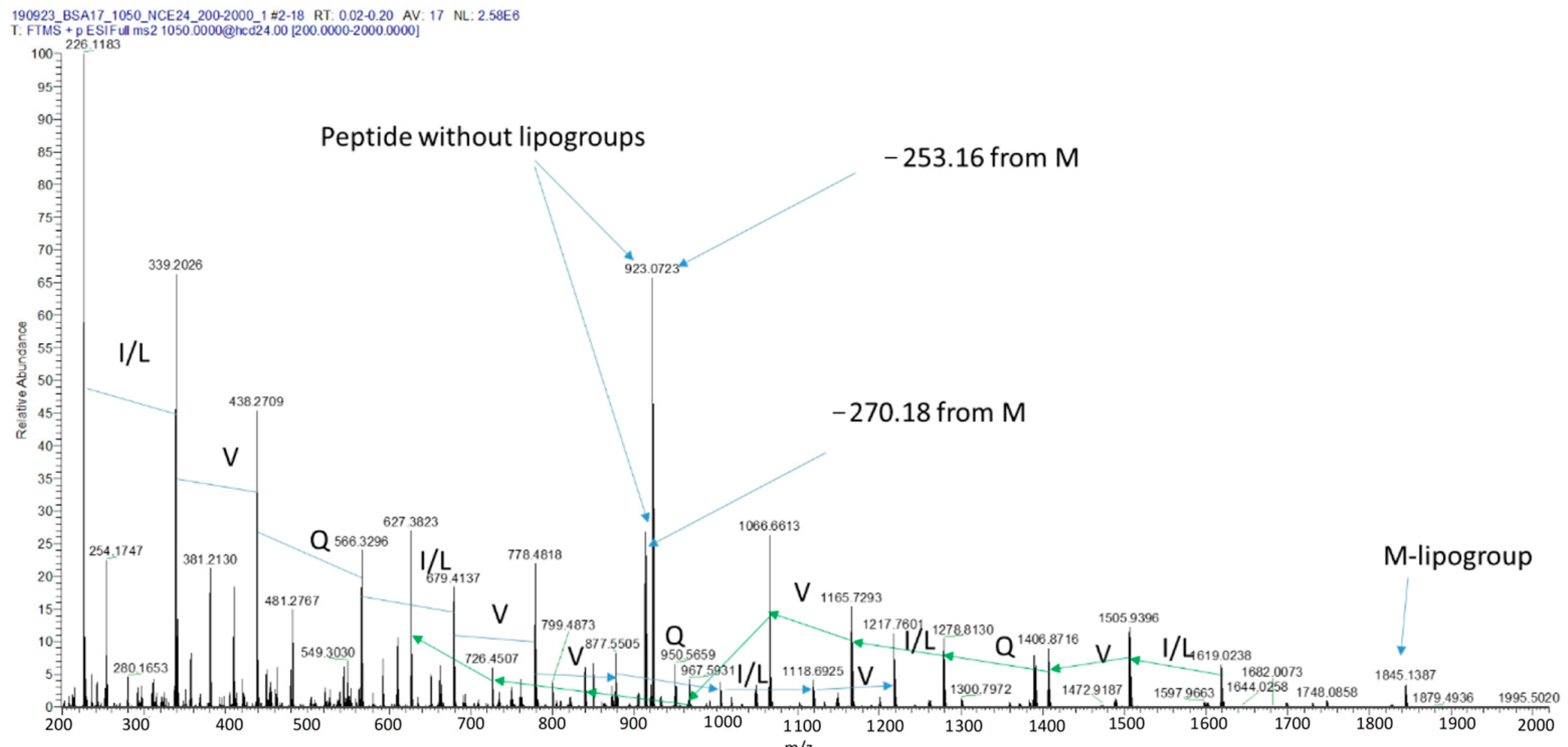
2.5. Identification of the Bioactive Lipopeptides Produced by BCA17
3. Materials and Methods
3.1. Plant Materials
3.2. Bacterial Strains, Culture Conditions, Inoculum Preparation
3.3. Fungal Isolates
3.4. Challenge Inoculation
3.4.1. Detached Canes
3.4.2. Potted Grapevines
- N = the calculated number of β-tubulin gene copies in 100 mg of wood sample
- g = the mean number of gene copies detected by qPCR
- d = the total gDNA extracted from 100 mg of wood (100 µL)
- c = DNA concentration (µL)
- t = the amount of DNA template (5 µL) in one reaction
3.5. Pseudomonas Colonization and Establishment in Grapevine Tissues
3.5.1. Production of Rifampicin-Resistant Strain
3.5.2. Pre-Planting Application of RifMut Strain
3.5.3. Post-Planting Application of RifMut Strain
3.6. Elucidation of Antagonistic Mechanisms of P. poae BCA17 against GTD Pathogens
3.6.1. Bioactivity of P. poae-Derived Culture Filtrates on Mycelial Growth and Spore Germination of GTD Pathogens
Effect of Culture Filtrates on Mycelial Growth
Effect of Culture Filtrates on Spore Germination
3.6.2. Identification of Secondary Metabolite Biosynthetic Gene Clusters
3.6.3. Extraction and Identification of Bioactive Lipopeptides
3.7. Data Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef]
- Gómez-Lama Cabanás, C.; Schiliró, E.; Valverde-Corredor, A.; Mercado-Blanco, J. The biocontrol endophytic bacterium Pseudomonas fluorescens PICF7 induces systemic defense responses in aerial tissues upon colonization of olive roots. Front. Microbiol. 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Rybakova, D.; Cernava, T.; Köberl, M.; Liebminger, S.; Etemadi, M.; Berg, G. Endophytes-assisted biocontrol: Novel insights in ecology and the mode of action of Paenibacillus. Plant Soil 2015, 405, 125–140. [Google Scholar] [CrossRef]
- Hong, C.E.; Park, J.M. Endophytic bacteria as biocontrol agents against plant pathogens: Current state-of-the-art. Plant Biotechnol. Rep. 2016, 10, 353–357. [Google Scholar] [CrossRef]
- Baker, K.F. Evolving concepts of biological control of plant pathogens. Annu. Rev. Phytopathol. 1987, 25, 67–85. [Google Scholar] [CrossRef]
- Kotze, C.; Van Niekerk, J.; Mostert, L.; Halleen, F.; Fourie, P. Evaluation of biocontrol agents for grapevine pruning wound protection against trunk pathogen infection. Phytopathol. Mediterr. 2011, 50, S247–S263. [Google Scholar]
- Ferreira, J.H.S.; Matthee, F.N.; Thomas, A.C. Biological control of Eutypa lata on grapevine by an antagonistic strain of Bacillus subtilis. Phytopathology 1991, 81, 283–287. [Google Scholar] [CrossRef]
- Schmidt, C.S.; Lorenz, D.; Wolf, G.A. Biological control of the grapevine dieback fungus Eutypa lata I: Screening of bacterial antagonists. J. Phytopathol. 2001, 149, 427–435. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Abou-Mansour, E.; Courteaux, B.; Rabenoelina, F.; Clément, C.; Fontaine, F.; Aziz, A. Bacillus subtilis PTA-271 counteracts Botryosphaeria dieback in grapevine, triggering immune responses and detoxification of fungal phytotoxins. Front. Plant Sci. 2019, 10, 25. [Google Scholar] [CrossRef]
- Leal, C.; Richet, N.; Guise, J.-F.; Gramaje, D.; Armengol, J.; Fontaine, F.; Trotel-Aziz, P. Cultivar contributes to the beneficial effects of Bacillus subtilis PTA-271 and Trichoderma atroviride SC1 to protect grapevine against Neofusicoccum parvum. Front. Microbiol. 2021, 12, 726132. [Google Scholar] [CrossRef]
- Álvarez-Pérez, J.M.; González-García, S.; Cobos, R.; Olego, M.Á.; Ibañez, A.; Díez-Galán, A.; Garzón-Jimeno, E.; Coque, J.J.R. Use of endophytic and rhizosphere actinobacteria from grapevine plants to reduce nursery fungal graft infections that lead to young grapevine decline. Appl. Environ. Microbiol. 2017, 83, e01564-17. [Google Scholar] [CrossRef]
- Martínez-Diz, M.D.P.; Díaz-Losada, E.; Andrés-Sodupe, M.; Bujanda, R.; Maldonado-González, M.M.; Ojeda, S.; Yacoub, A.; Rey, P.; Gramaje, D. Field evaluation of biocontrol agents against black-foot and Petri-diseases of grapevine. Pest Manag. Sci. 2021, 77, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Arseneault, T.; Goyer, C.; Filion, M. Biocontrol of potato common scab is associated with high Pseudomonas fluorescens LBUM223 populations and phenazine-1-carboxylic acid biosynthetic transcript accumulation in the potato geocaulosphere. Phytopathology 2016, 106, 963–970. [Google Scholar] [CrossRef]
- Hultberg, M.; Bengtsson, T.; Liljeroth, E. Late blight on potato is suppressed by the biosurfactant-producing strain Pseudomonas koreensis 2.74 and its biosurfactant. BioControl 2010, 55, 543–550. [Google Scholar] [CrossRef]
- D’aes, J.; Hua, G.K.H.; De Maeyer, K.; Pannecoucque, J.; Forrez, I.; Ongena, M.; Dietrich, L.E.P.; Thomashow, L.S.; Mavrodi, D.V.; Höfte, M. Biological control of Rhizoctonia root rot on bean by phenazine- and cyclic lipopeptide-producing Pseudomonas CMR12a. Phytopathology 2011, 101, 996–1004. [Google Scholar] [CrossRef]
- Solanki, M.K.; Singh, R.K.; Srivastava, S.; Kumar, S.; Kashyap, P.L.; Srivastava, A.K.; Arora, D.K. Isolation and characterization of siderophore producing antagonistic rhizobacteria against Rhizoctonia solani. J. Basic Microbiol. 2014, 54, 585–597. [Google Scholar] [CrossRef]
- Bardas, G.A.; Lagopodi, A.L.; Kadoglidou, K.; Tzavella-Klonari, K. Biological control of three Colletotrichum lindemuthianum races using Pseudomonas chlororaphis PCL1391 and Pseudomonas fluorescens WCS365. Biol. Control 2009, 49, 139–145. [Google Scholar] [CrossRef]
- Aiello, D.; Restuccia, C.; Stefani, E.; Vitale, A.; Cirvilleri, G. Postharvest biocontrol ability of Pseudomonas synxantha against Monilinia fructicola and Monilinia fructigena on stone fruit. Biocatal. Agric. Biotechnol. 2019, 13, 46–52. [Google Scholar] [CrossRef]
- Weller, D.M.; Cook, R.J. Suppression of take-all of wheat by seed treatments with fluorescent pseudomonads. Phytopathology 1983, 73, 463–469. [Google Scholar] [CrossRef]
- Thomashow, L.S.; Weller, D.M. Role of a phenazine antibiotic from Pseudomonas fluorescens in biological control of Gaeumannomyces graminis var. tritici. J. Bacteriol. 1988, 170, 3499–3508. [Google Scholar] [CrossRef]
- Keel, C.; Schnider, U.; Maurhofer, M.; Voisard, C.; Laville, J.; Burger, U.; Wirthner, P.; Haas, D.; Défago, G. Suppression of root diseases by Pseudomonas fluorescens CHA0: Importance of the bacterial secondary metabolite 2,4-diacetylphloroglucinol. Mol. Plant-Microbe Interact. 1992, 5, 4–13. [Google Scholar] [CrossRef]
- Ait Barka, E.; Gognies, S.; Nowak, J.; Audran, J.C.; Belarbi, A. Inhibitory effect of endophyte bacteria on Botrytis cinerea and its influence to promote grapevine growth. Biol. Control 2002, 24, 135–142. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Couderchet, M.; Biagianti, S.; Aziz, A. Characterization of new biocontrol agents Acinetobacter, Bacillus, Pantoea and Pseudomonas spp, 2008. mediating grapevine resistance against Botrytis cinerea. Environ. Exp. Bot. 2008, 64, 21–32. [Google Scholar] [CrossRef]
- Ziedan, E.H.E.; El-Mohamedy, R.S.R. Application of Pseudomonas fluorescens for controlling root-rot disease of grapevine. Res. J. Agric. Biol. Sci. 2008, 4, 346–353. [Google Scholar]
- Khmel, I.A.; Sorokina, T.A.; Lemanova, N.B.; Lipasova, V.A.; Metlitski, O.Z.; Burdeinaya, T.V.; Chernin, L.S. Biological control of crown gall in grapevine and raspberry by two Pseudomonas spp. with a wide spectrum of antagonistic activity. Biocontrol Sci. Technol. 1998, 8, 45–57. [Google Scholar] [CrossRef]
- Bell, C.R.; Dickie, G.A.; Harvey, W.L.G.; Chan, J.W.Y.F. Endophytic bacteria in grapevine. Can. J. Microbiol. 1995, 41, 46–53. [Google Scholar] [CrossRef]
- West, E.R.; Cother, E.J.; Steel, C.C.; Ash, G.J. The characterization and diversity of bacterial endophytes of grapevine. Can. J. Microbiol. 2010, 56, 209–216. [Google Scholar] [CrossRef]
- Deyett, E.; Roper, M.C.; Ruegger, P.; Yang, J.; Borneman, J.; Rolshausen, P.E. Microbial landscape of the grapevine endosphere in the context of Pierce’s disease. Phytobiomes J. 2017, 1, 138–149. [Google Scholar] [CrossRef]
- Niem, J.M.; Billones-Baaijens, R.; Stodart, B.; Savocchia, S. Diversity profiling of grapevine microbial endosphere and antagonistic potential of endophytic Pseudomonas against Grapevine trunk diseases. Front. Microbiol. 2020, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Weller, D.M. Biological control of soilborne plant pathogens in the rhizosphere with bacteria. Annu. Rev. Phytopathol. 1988, 26, 379–407. [Google Scholar] [CrossRef]
- Keel, C.; Wirthner, P.H.; Oberhänsli, T.H.; Voisard, C.; Burger, P.U.; Haas, D.; Défago, G. Pseudomonads as antagonists of plant pathogens in the rhizosphere–role of the antibiotic 2,4 diacetylphloroglucinol in the suppression of black root-rot of tobacco. Symbiosis 1990, 9, 327–341. [Google Scholar]
- Raaijmakers, J.; Weller, D.M. Natural plant protection by 2, 4 diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils. Mol. Plant-Microbe Interact. 1998, 11, 144–152. [Google Scholar] [CrossRef]
- Ramette, A.; Moënne-Loccoz, Y.; Défago, G. Prevalence of fluorescent pseudomonads producing antifungal phloroglucinols and/or hydrogen cyanide in soils naturally suppressive or conducive to tobacco black root rot. FEMS Microbiol. Ecol. 2003, 44, 35–43. [Google Scholar] [CrossRef]
- Bull, C.T.; Weller, D.M.; Thomashow, L.S. Relationship between root colonization and suppression of Gaeumannomyces graminis var. tritici by Pseudomonas fluorescence strain 2-79. Phytopathology 1991, 81, 954–959. [Google Scholar] [CrossRef]
- Mazzola, M.; Cook, R.J.; Thomashow, L.S.; Weller, D.M.; Pierson, L.S. Contribution of phenazine antibiotic biosynthesis to the ecological competence of fluorescent pseudomonads in soil habitats. Appl. Environ. Microbiol. 1992, 58, 2616–2624. [Google Scholar] [PubMed]
- Raaijmakers, J.M.; Vlami, M.; de Souza, J.T. Antibiotic production by bacterial biocontrol agents. Antonie Leeuwenhoek. 2002, 81, 537–547. [Google Scholar] [CrossRef]
- Scher, F.M.; Baker, R. Effect of Pseudomonas putida and a synthetic iron chelator on induction of soil suppressiveness to Fusarium wilt pathogens. Phytopathology 1982, 72, 1567–1573. [Google Scholar] [CrossRef]
- Ahl, P.; Voisard, C.; Défago, G. Iron bound–siderophores, cyanic acid, and antibiotics involved in suppression of Thielaviopsis basicola by a Pseudomonas fluorescens strains. J. Phytopathol. 1986, 116, 121–134. [Google Scholar] [CrossRef]
- Becker, O.; Cook, R.J. Role of siderophores in suppression of Pythium species and production of increased growth response of wheat by fluorescent pseudomonads. Phytopathology 1988, 78, 778–782. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Pseudomonas siderophores: A mechanism explaining disease suppressive soils. Curr. Microbiol. 1980, 4, 317–320. [Google Scholar] [CrossRef]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.H.; Sørensen, J. Production of cyclic lipopeptides by Pseudomonas fluorescence strains in bulk soil and in the sugar beet rhizosphere. Appl. Environ. Microbiol. 2003, 69, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Martinez, E.P.; Osorio, J.A. Preliminary studies for the production of an active biosurfactant against Phytophthora infestans (Mont.) de Bary. Rev. Corpoica Cienc. Y Technol. Agropecu. 2007, 8, 5–16. [Google Scholar] [CrossRef]
- Tran, H.; Ficke, A.; Asiimwe, T.; Höfte, M.; Raaijmakers, J.M. Role of the cyclic lipopeptide massetolide A in biological control of Phytophthora infestans and in colonization of tomato plants by Pseudomonas fluorescens. New Phytol. 2007, 175, 731–742. [Google Scholar] [CrossRef]
- Hultberg, M.; Alsberg, T.; Khalil, S.; Alsanius, B. Suppression of disease in tomato infected by Pythium ultimum with a biosurfactant produced by Pseudomonas koreensis. BioControl 2010, 55, 435–444. [Google Scholar] [CrossRef]
- Stanghellini, M.E.; Miller, R.M. Biosurfactants: Their identity and potential efficacy in the biological control of zoosporic plant pathogens. Plant Dis. 1997, 81, 4–12. [Google Scholar] [CrossRef]
- Nielsen, T.H.; Christophersen, C.; Anthoni, U.; Sørensen, J. Viscosinamide, a new cyclic depsipeptide with surfactant and antifungal properties produced by Pseudomonas fluorescens DR54. J. Appl. Microbiol. 1999, 86, 80–90. [Google Scholar] [CrossRef]
- Voisard, C.; Keel, C.; Haas, D.; Défago, G. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. EMBO J. 1989, 8, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Lo Cantore, P.; Giorgio, A.; Lacobellis, N.S. Bioactivity of volatile organic compounds produced by Pseudomonas tolaasii. Front. Microbiol. 2015, 6, 1082. [Google Scholar] [CrossRef]
- Rojas-Solís, D.; Zetter-Salmón, E.; Contreras-Pérez, M.; Rocha-Granados, M.D.C.; MacíasRodríguez, L.; Santoyo, G. Pseudomonas stutzeri E25 and Stenotrophomonas maltophilia CR71 endophytes produce antifungal volatile organic compounds and exhibit additive plant growth-promoting effects. Biocatal. Agric. Biotechnol. 2018, 13, 46–52. [Google Scholar] [CrossRef]
- Suslow, T.V.; Schroth, M.N. Rhizobacteria of sugar beets: Effects of seed application and root colonization on yield. Phytopathology 1982, 72, 199–206. [Google Scholar] [CrossRef]
- Santoyo, G.; Orozco-Mosqueda, M.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: A review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Lorito, M.; Peterbauer, C.; Hayes, C.K.; Harman, G.E. Synergistic interaction between fungal cell-wall degrading enzymes and different antifungal compounds enhances inhibition of spore germination. Microbiology 1994, 140, 623–629. [Google Scholar] [CrossRef]
- Smith, J.A.; Hammerschmidt, R.; Fulbright, D.W. Rapid induction of systemic resistance in cucumber by Pseudomonas syringae pv syringae. Physiol. Mol. Plant Pathol. 1991, 38, 223–235. [Google Scholar] [CrossRef]
- van Peer, R.; Niemann, G.J.; Schippers, B. Induced resistance and phytoalexin accumulation in biological control of Fusarium wilt of carnation by Pseudomonas spp. strain WCS417r. Phytopathology 1991, 81, 728–734. [Google Scholar] [CrossRef]
- Benhamou, N.; Bélanger, R.R.; Paulitz, T.C. Induction of differential host responses by Pseudomonas fluorescens in Ri-T DNA transformed pea roots upon challenge with Fusarium oxysporum f.sp. pisi and Pythium ultimum. Phytopathology 1996, 86, 1174–1185. [Google Scholar] [CrossRef]
- M’Piga, P.; Bélanger, R.R.; Paulitz, T.C.; Benhamou, N. Increased resistance to Fusarium oxysporum f. sp. radicis-lycopersici in tomato plants treated with the endophytic bacterium Pseudomonas fluorescens strain 63-28. Physiol. Mol. Plant Pathol. 1997, 50, 301–320. [Google Scholar] [CrossRef]
- Audenaert, K.; Pattery, T.; Cornelis, P.; Höfte, M. Induction of systemic resistance to Botrytis cinerea in tomato by Pseudomonas aeruginosa 7NSK2: Role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant-Microbe Interact. 2002, 15, 1147–1156. [Google Scholar] [CrossRef]
- Iavicoli, A.; Boutet, E.; Buchala, A. Métraux, J.-P. Induced systemic resistance in Arabidopsis thaliana in response to root inoculation with Pseudomonas fluorescens CHA0. Mol. Plant Microbe Interact. 2003, 16, 851–858. [Google Scholar] [CrossRef]
- de Vleesschauwer, D.; Djavaheri, M.; Bakker, P.A.H.M.; Höfte, M. Pseudomonas fluorescens WCS374r-induced systemic resistance in rice against Magnaporthe oryzae is based on pseudobactin-mediated priming for a salicylic acid- repressible multifaceted defense response. Plant Physiol. 2008, 148, 1996–2012. [Google Scholar] [CrossRef]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; De Los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. AntiSMASH 4.0—Improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef]
- Nielsen, T.H.; Sørensen, D.; Tobiasen, C.; Andersen, J.B.; Christeophersen, C.; Givskov, M.; Sørensen, J. Antibiotic and biosurfactant properties of cyclic lipopeptides produced by fluorescent Pseudomonas spp. from the sugar beet rhizosphere. Appl. Environ. Microbiol. 2002, 68, 3416–3423. [Google Scholar] [CrossRef]
- de Souza, J.T.; de Boer, M.; de Waard, P.; van Beek, T.A.; Raaijmakers, J.M. Biochemical, genetic, and zoosporicidal properties of cyclic lipopeptide surfactants produced by Pseudomonas fluorescens. Appl. Environ. Microbiol. 2003, 69, 7161–7172. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, J.M.; de Bruijn, I.; de Kock, M.J.D. Cyclic lipopeptide production by plant-associated Pseudomonas spp.: Diversity, activity, biosynthesis, and regulation. Mol. Plant-Microbe Interact. 2006, 19, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Zachow, C.; Jahanshah, G.; de Bruijn, I.; Song, C.; Ianni, F.; Pataj, Z.; Gerhardt, H.; Pianet, I.; Lämmerhofer, M.; Berg, G. The novel lipopeptide poaeamide of the endophyte Pseudomonas poae RE*1–1–14 is involved in pathogen suppression and root colonization. Mol. Plant-Microbe Interact. 2015, 28, 800–810. [Google Scholar] [CrossRef]
- Geudens, N.; Martins, J.C. Cyclic lipodepsipeptides from Pseudomonas spp.–biological swiss-army knives. Front. Microbiol. 2018, 9, 1867. [Google Scholar] [CrossRef] [PubMed]
- Bassarello, C.; Lazzaroni, S.; Bifulco, G.; Lo Cantore, P.; Iacobellis, N.S.; Riccio, R.; Gomez-Paloma, L.; Evidente, A. Tolaasins A-E, five new lipodepsipeptides produced by Pseudomonas tolaasii. J. Nat. Prod. 2004, 67, 811–816. [Google Scholar] [CrossRef]
- Wunderlich, N.; Ash, G.J.; Steel, C.C.; Raman, H.; Savocchia, S. Association of Botryosphaeriaceae grapevine trunk disease fungi with the reproductive structures of Vitis vinifera. Vitis 2011, 50, 89–96. [Google Scholar]
- Qiu, Y.; Steel, C.C.; Ash, G.J.; Savocchia, S. Survey of Botryosphaeriaceae associated with grapevine decline in the Hunter Valley and Mudgee grape growing regions of New South Wales. Australas. Plant Pathol. 2011, 40, 1–11. [Google Scholar] [CrossRef]
- Billones-Baaijens, R.; Úrbez-Torres, J.R.; Liu, M.; Ayres, M.; Sosnowski, M.; Savocchia, S. Molecular methods to detect and quantify Botryosphaeriaceae inocula associated with grapevine dieback in Australia. Plant Dis. 2018, 102, 1489–1499. [Google Scholar] [CrossRef]
- Pouzoulet, J.; Mailhac, N.; Couderc, C.; Besson, X.; Daydé, J.; Lummerzheim, M.; Jacques, A. A method to detect and quantify Phaeomoniella chlamydospora and Phaeoacremonium aleophilum DNA in grapevine-wood samples. Appl. Microbiol. Biotechnol. 2013, 97, 10163–10175. [Google Scholar] [CrossRef]
- Adorada, D.L.; Stodart, B.J.; Pangga, I.B.; Ash, G.J. Implications of bacterial contaminated seed lots and endophytic colonization by Pseudomonas fuscovaginae on rice establishment. Plant Pathol. 2015, 64, 43–50. [Google Scholar] [CrossRef]
- Talboys, P.W. A culture-medium aiding the identification of Verticillium albo-atrum and V. dahliae. Plant Pathol. 1960, 9, 57–58. [Google Scholar] [CrossRef]
- Niem, J.; Billones-Baaijens, R.; Savocchia, S.; Stodart, B. Draft genome sequences of endophytic Pseudomonas spp. isolated from grapevine tissue and antagonistic to grapevine trunk disease pathogens. Microbiol. Resour. Announc. 2019, 8, e00345-19. [Google Scholar] [CrossRef] [PubMed]
- Sajitha, K.L.; Dev, S.A.; Florence, E.J.M. Identification and characterization of lipopeptides from Bacillus subtilis B1 against sapstain fungus of rubberwood through MALDI-TOF-MS and RT-PCR. Curr. Microbiol. 2016, 73, 46–53. [Google Scholar] [CrossRef]
- Behrendt, U.; Ulrich, A.; Schumann, P. Fluorescent pseudomonads associated with the phyllosphere of grasses; Pseudomonas trivialis sp. nov., Pseudomonas poae sp. nov. and Pseudomonas congelans sp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1461–1469. [Google Scholar] [CrossRef] [PubMed]
- Zachow, C.; Fatehi, J.; Cardinale, M.; Tilcher, R.; Berg, G. Strain-specific colonization pattern of Rhizoctonia antagonists in the root system of sugar beet. FEMS Microbiol. Ecol. 2010, 74, 124–135. [Google Scholar] [CrossRef]
- Müller, H.; Zachow, C.; Alavi, M.; Tilcher, R.; Krempl, P.M.; Thallinger, G.G.; Berg, G. Complete genome sequence of the sugar beet endophyte Pseudomonas poae RE*1-1-14, a disease-suppressive bacterium. Genome Announc. 2013, 1, e00020-13. [Google Scholar] [CrossRef]
- Wicaksono, W.A.; Jones, E.E.; Monk, J.; Ridgway, H.J. The bacterial signature of Leptospermum scoparium (mānuka) reveals core and accessory communities with bioactive properties. PLoS ONE 2016, 11, e0163717. [Google Scholar] [CrossRef]
- Wicaksono, W.A.; Jones, E.E.; Monk, J.; Ridgway, H.J. Using bacterial endophytes from a New Zealand native medicinal plant for control of grapevine trunk diseases. Biol. Control 2017, 114, 65–72. [Google Scholar] [CrossRef]
- Tarbah, F.; Goodman, R.N. Systemic spread of Agrobacterium tumefaciens biovar 3 in the vascular system of grapes. Phytopathology 1987, 77, 915–920. [Google Scholar] [CrossRef]
- Thorne, E.T.; Young, B.M.; Young, G.M.; Stevenson, J.F.; Labavitch, J.M.; Matthews, M.A.; Rost, T.L. The structure of xylem vessels in grapevines (Vitaceae) and a possible passive mechanism for the systemic spread of bacterial disease. Am. J. Bot. 2006, 93, 497–504. [Google Scholar] [CrossRef]
- Billones-Baaijens, R.; Ridgway, H.J.; Jones, E.E.; Cruickshank, R.H.; Jaspers, M.V. Prevalence and distribution of Botryosphaeriaceae species in New Zealand grapevine nurseries. Eur. J. Plant Pathol. 2012, 135, 175–185. [Google Scholar] [CrossRef]
- Leal, C.; Gramaje, D.; Fontaine, F.; Richet, N.; Trotel-Aziz, P.; Armengol, J. Evaluation of Bacillus subtilis PTA-271 and Trichoderma atroviride SC1 to control Botryosphaeria dieback and black-foot pathogens in grapevine propagation material. Pest Manag. Sci. 2023, 79, 1674–1683. [Google Scholar] [CrossRef] [PubMed]
- Howell, C.R.; Stipanovic, R.D. Suppression of Pythium ultimum-induced damping-off of cotton seedlings by Pseudomonas fluorescens and its antibiotic, pyoluteorin. Phytopathology 1980, 70, 712–715. [Google Scholar] [CrossRef]
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W.I.I.; Lim, C.K.; Shaffer, B.T.; Elbourne, L.D.H.; Stockwell, V.O.; Hartney, S.L.; Breakwell, K.; et al. Comparative genomics of plant-associated Pseudomonas spp.: Insights into diversity and inheritance of traits involved in multitrophic interactions. PLoS Genet. 2012, 8, e1002784. [Google Scholar] [CrossRef]
- Arseneault, T.; Goyer, C.; Filion, M. Phenazine production by Pseudomonas sp. LBUM223 contributes to the biological control of potato common scab. Phytopathology 2013, 103, 995–1000. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybro., O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef]
- Calderón, C.E.; Ramos, C.; de Vicente, A.; Cazorla, F.M. Comparative genomic analysis of Pseudomonas chlororaphis PCL1606 reveals new insight into antifungal compounds involved in biocontrol. Mol. Plant-Microbe Interact. 2018, 28, 249–260. [Google Scholar] [CrossRef]
- Gross, H.; Loper, J.E. Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep. 2009, 26, 1408–1446. [Google Scholar] [CrossRef]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Wieland Brown, L.C.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [PubMed]
- De Bruijn, I.; de Kock, M.J.D.; Yang, M.; de Waard, P.; van Beek, T.A.; Raaijmakers, J.M. Genome-based discovery, structure prediction and functional analysis of cyclic lipopeptide antibiotics in Pseudomonas species. Mol. Microbiol. 2007, 63, 417–428. [Google Scholar] [CrossRef]
- Alsohim, A.S.; Taylor, T.B.; Barrett, G.A.; Gallie, J.; Zhang, X.; Altamirano-Junqueira, A.E.; Johnson, L.J.; Rainey, P.B.; Jackson, R.W. The biosurfactant viscosin produced by Pseudomonas fluorescens SBW25 aids spreading motility and plant growth promotion. Environ. Microbiol. 2014, 16, 2267–2281. [Google Scholar] [CrossRef]
- Yang, M.; Wen, S.; Mavrodi, M.V.; Mavrodi, O.V.; von Wettstein, D.; Thomashow, L.S.; Guo, J. Biological control of wheat root diseases by the CLP-producing strain Pseudomonas fluorescens HC1-07. Phytopathology 2014, 104, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Kakembo, D.; Lee, Y.H. Analysis of traits for biocontrol performance of Pseudomonas parafulva JBCS1880 against bacterial pustule in soybean plants. Biol. Control 2019, 134, 72–81. [Google Scholar] [CrossRef]
- Olorunleke, F.E.; Hua, G.K.H.; Kieu, N.P.; Ma, Z.; Höfte, M. Interplay between orfamides, sessilins and phenazines in the control of Rhizoctonia diseases by Pseudomonas sp. CMR12a. Environ. Microbiol. Rep. 2015, 7, 774–781. [Google Scholar] [CrossRef]
- Olorunleke, F.E.; Kieu, N.P.; De Waele, E.; Timmerman, M.; Ongena, M.; Höfte, M. Coregulation of the cyclic lipopeptides orfamide and sessilin in the biocontrol strain Pseudomonas sp. CMR12a. Microbiol. Open 2017, 6, e499. [Google Scholar] [CrossRef]
- Omoboye, O.O.; Geudens, N.; Duban, M.; Chevalier, M.; Flahaut, C.; Martins, J.C.; Leclère, V.; Oni, F.E.; Höfte, M. Pseudomonas sp. COW3 produces new bananamide-type cyclic lipopeptides with antimicrobial activity against Pythium myriotylum and Pyricularia oryzae. Molecules 2019, 24, 4170. [Google Scholar] [CrossRef]
- Rokni-Zadeh, H.; Li, W.; Sanchez-Rodriguez, A.; Sinnaeve, D.; Rozenski, J.; Martins, J.C.; De Mot, R. Genetic and functional characterization of cyclic lipopeptide white-line-inducing principle (WLIP) production by rice rhizosphere isolate Pseudomonas putida RW10S2. Appl. Environ. Microbiol. 2012, 78, 4826–4834. [Google Scholar] [CrossRef]
- Götze, S.; Herbst-Irmer, R.; Klapper, M.; Görls, H.; Schneider, K.R.A.; Barnett, R.; Burks, T.; Neu, U.; Stallforth, P. Structure, biosynthesis, and biological activity of the cyclic lipopeptide anikasin. ACS Chem. Biol. 2017, 12, 2498–2502. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niem, J.M.; Billones-Baaijens, R.; Stodart, B.J.; Reveglia, P.; Savocchia, S. Biocontrol Potential of an Endophytic Pseudomonas poae Strain against the Grapevine Trunk Disease Pathogen Neofusicoccum luteum and Its Mechanism of Action. Plants 2023, 12, 2132. https://doi.org/10.3390/plants12112132
Niem JM, Billones-Baaijens R, Stodart BJ, Reveglia P, Savocchia S. Biocontrol Potential of an Endophytic Pseudomonas poae Strain against the Grapevine Trunk Disease Pathogen Neofusicoccum luteum and Its Mechanism of Action. Plants. 2023; 12(11):2132. https://doi.org/10.3390/plants12112132
Chicago/Turabian StyleNiem, Jennifer Millera, Regina Billones-Baaijens, Benjamin J. Stodart, Pierluigi Reveglia, and Sandra Savocchia. 2023. "Biocontrol Potential of an Endophytic Pseudomonas poae Strain against the Grapevine Trunk Disease Pathogen Neofusicoccum luteum and Its Mechanism of Action" Plants 12, no. 11: 2132. https://doi.org/10.3390/plants12112132