
Identification and Expression Analysis of Phosphatidylinositol Transfer Proteins Genes in Rice
 and
and
Abstract
:1. Introduction
1.1. PITPs in Animals
1.2. PITPs in Plants
2. Results
2.1. Identification and Bioinformatics Analysis of PITP Genes in Rice
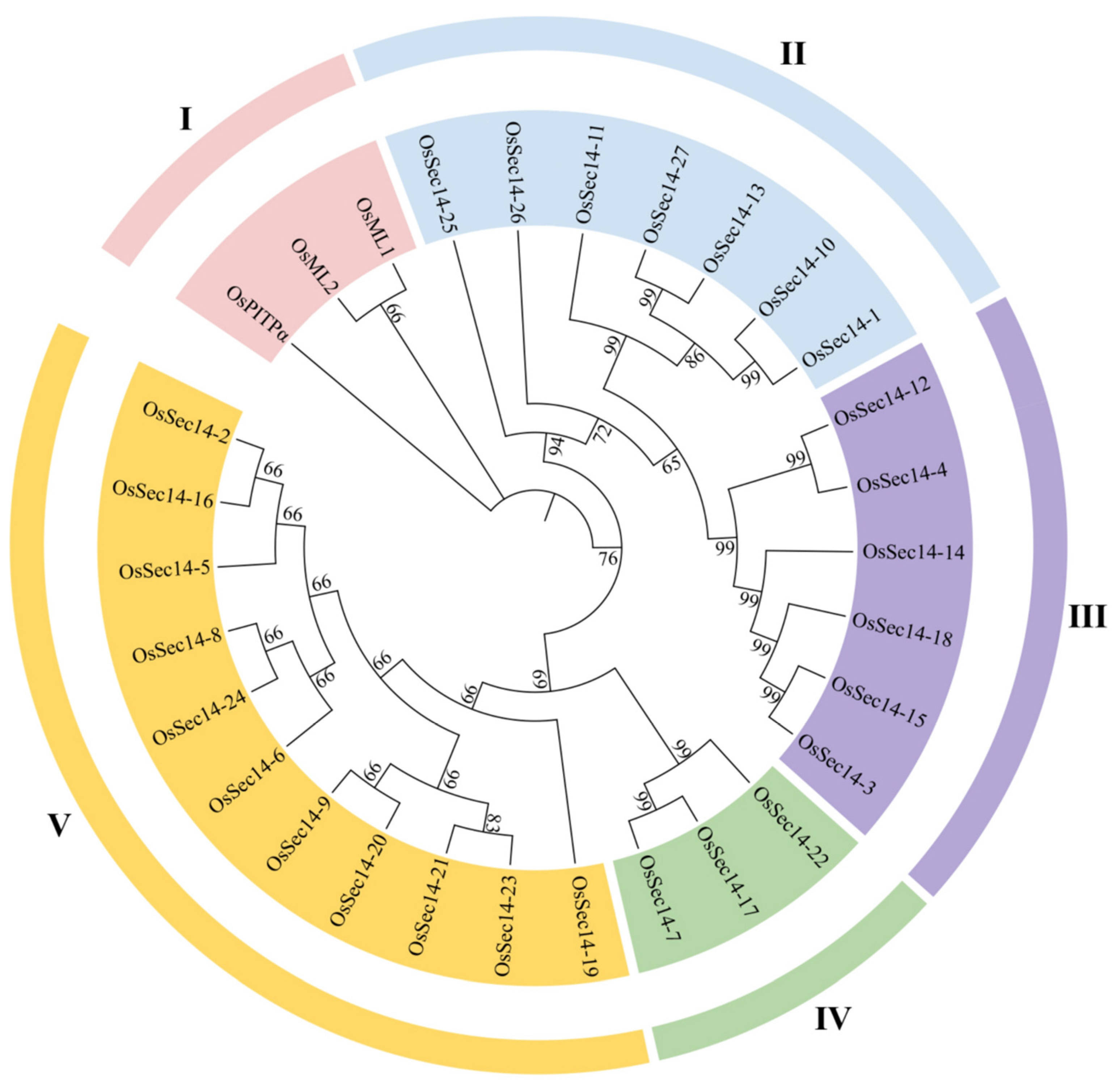
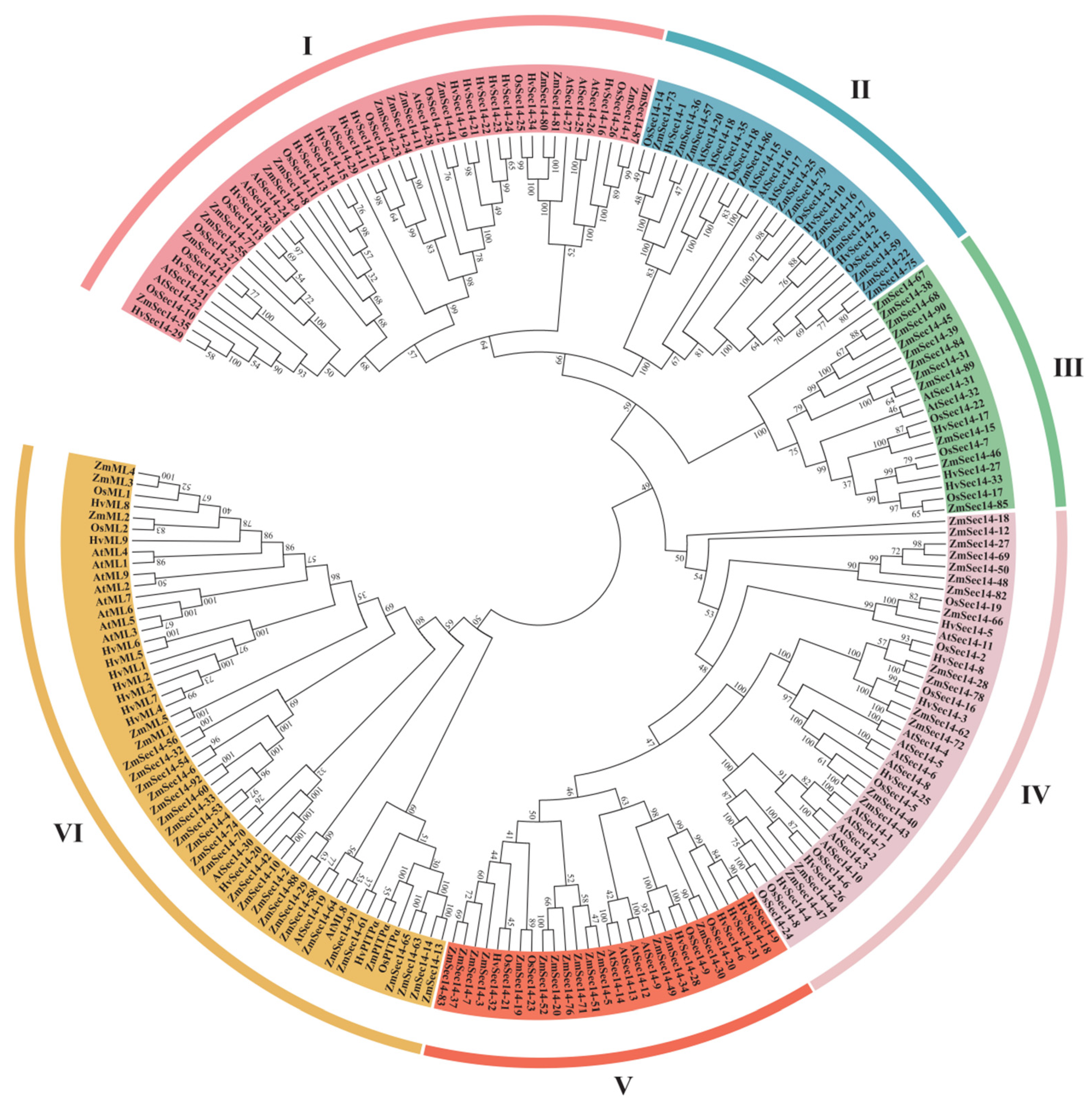
2.2. Evolution Analysis of PITPs in Plants
2.3. Analysis of Collinearity between PITPs
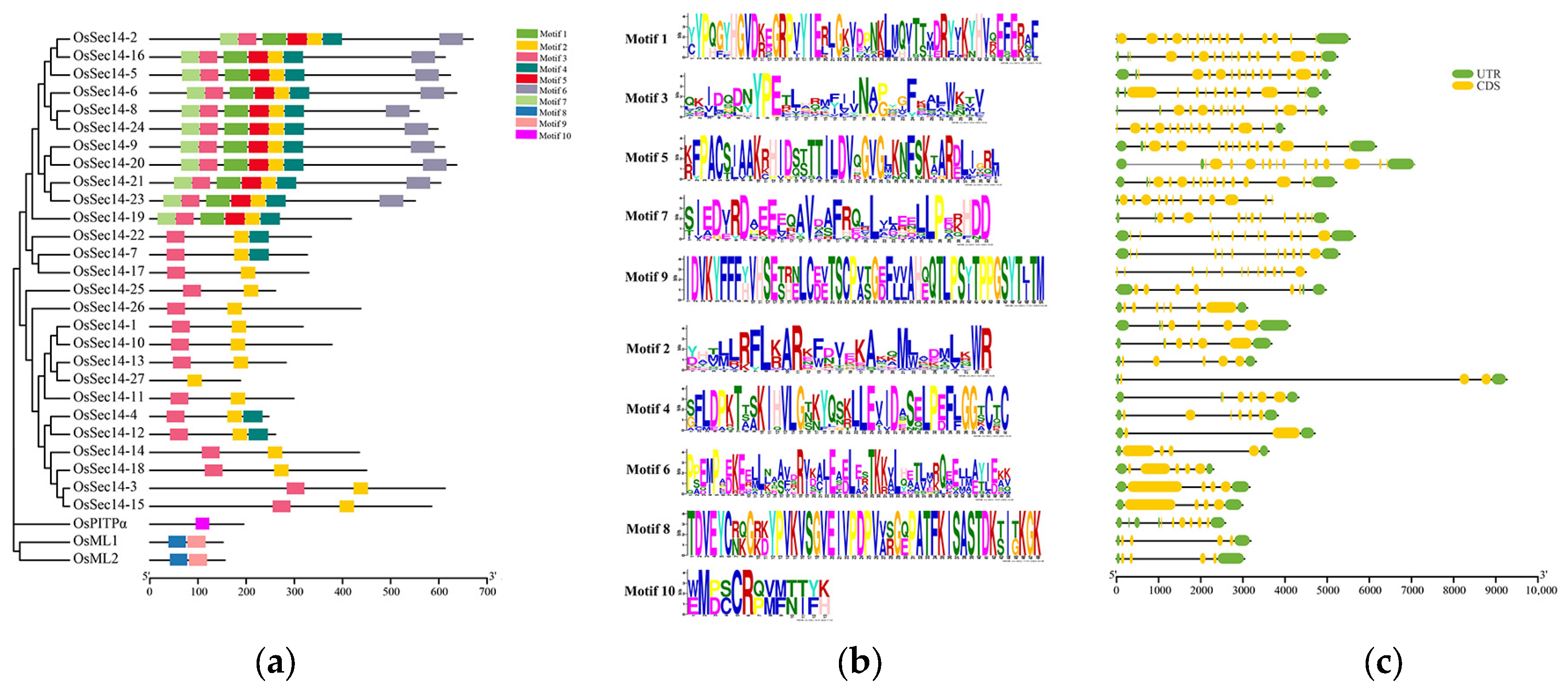
2.4. Examining the Conserved Motifs and Gene Structure of OsPITPs
2.5. The Functional Domains of PITPs Family Proteins
2.6. Predicting Cis-Acting Elements of PITPs Family Genes
2.7. Expression Levels of PITPs Genes during Infection of Rice Blast Fungus
2.8. Subcellular Localization of PITP Family Proteins
3. Materials and Methods
3.1. Identification of Rice PITP Gene Family Members
3.2. Physicochemical Properties and Subcellular Localization Analysis
3.3. System Evolutionary Analysis
3.4. Collinearity Analysis
3.5. Gene Structure and Protein Motif Analysis
3.6. Functional Domain Analysis
3.7. Subcellular Localization
3.8. Transcriptome Data
3.9. RNA Extraction and qRT-PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Allen-Baume, V.; Ségui, B.; Cockcroft, S. Current thoughts on the phosphatidylinositol transfer protein family. FEBS Lett. 2002, 531, 74–80. [Google Scholar] [CrossRef]
- Ashlin, T.G.; Blunsom, N.J.; Cockcroft, S. Courier service for phosphatidylinositol: PITPs deliver on demand. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 866, 158985. [Google Scholar] [CrossRef]
- Hamilton, B.A.; Smith, D.J.; Mueller, K.L.; Kerrebrock, A.W.; Bronson, R.T.; van Berkel, V.; Daly, M.J.; Kruglyak, L.; Reeve, M.P.; Nemhauser, J.L.; et al. The vibrator mutation causes neurodegeneration via reduced expression of PITP alpha: Positional complementation cloning and extragenic suppression. Neuron 1997, 18, 711–722. [Google Scholar] [CrossRef]
- Alb, J.G., Jr.; Cortese, J.D.; Phillips, S.E.; Albin, R.L.; Nagy, T.R.; Hamilton, B.A.; Bankaitis, V.A. Mice lacking phosphatidylinositol transfer protein-alpha exhibit spinocerebellar degeneration, intestinal and hepatic steatosis, and hypoglycemia. J. Biol. Chem. 2003, 278, 33501–33518. [Google Scholar] [CrossRef]
- Carvou, N.; Holic, R.; Li, M.; Futter, C.; Skippen, A.; Cockcroft, S. Phosphatidylinositol- and phosphatidylcholine-transfer activity of PITPbeta is essential for COPI-mediated retrograde transport from the Golgi to the endoplasmic reticulum. J. Cell Sci. 2010, 123 Pt 8, 1262–1273. [Google Scholar] [CrossRef]
- Trivedi, D.; Padinjat, R. RdgB proteins: Functions in lipid homeostasis and signal transduction. Biochim. Biophys. Acta 2007, 1771, 692–699. [Google Scholar] [CrossRef]
- Vihtelic, T.S.; Goebl, M.; Milligan, S.; O’Tousa, J.E.; Hyde, D.R. Localization of Drosophila retinal degeneration B, a membrane-associated phosphatidylinositol transfer protein. J. Cell Biol. 1993, 122, 1013–1022. [Google Scholar] [CrossRef]
- Raghu, P.; Basak, B.; Krishnan, H. Emerging perspectives on multidomain phosphatidylinositol transfer proteins. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158984. [Google Scholar] [CrossRef]
- Garner, K.; Hunt, A.N.; Koster, G.; Somerharju, P.; Groves, E.; Li, M.; Raghu, P.; Holic, R.; Cockcroft, S. Phosphatidylinositol transfer protein, cytoplasmic 1 (PITPNC1) binds and transfers phosphatidic acid. J. Biol. Chem. 2012, 287, 32263–32276. [Google Scholar] [CrossRef]
- Cockcroft, S.; Garner, K. Potential role for phosphatidylinositol transfer protein (PITP) family in lipid transfer during phospholipase C signalling. Adv. Biol. Regul. 2013, 53, 280–291. [Google Scholar] [CrossRef]
- Saito, K.; Tautz, L.; Mustelin, T. The lipid-binding SEC14 domain. Biochim. Biophys. Acta 2007, 1771, 719–726. [Google Scholar] [CrossRef]
- Sha, B.; Phillips, S.E.; Bankaitis, V.A.; Luo, M. Crystal structure of the Saccharomyces cerevisiae phosphatidylinositol-transfer protein. Nature 1998, 391, 506–510. [Google Scholar] [CrossRef]
- Chinen, K.; Takahashi, E.; Nakamura, Y. Isolation and mapping of a human gene (SEC14L), partially homologous to yeast SEC14, that contains a variable number of tandem repeats (VNTR) site in its 3’ untranslated region. Cytogenet. Cell Genet. 1996, 73, 218–223. [Google Scholar] [CrossRef]
- Sonbul, S.N.; Aleskandarany, M.A.; Kurozumi, S.; Joseph, C.; Toss, M.S.; Diez-Rodriguez, M.; Nolan, C.C.; Mukherjee, A.; Martin, S.; Caldas, C.; et al. Saccharomyces cerevisiae-like 1 (SEC14L1) is a prognostic factor in breast cancer associated with lymphovascular invasion. Mod. Pathol. 2018, 31, 1675–1682. [Google Scholar] [CrossRef]
- Li, M.T.; Di, W.; Xu, H.; Yang, Y.K.; Chen, H.W.; Zhang, F.X.; Zhai, Z.H.; Chen, D.Y. Negative regulation of RIG-I-mediated innate antiviral signaling by SEC14L1. J. Virol. 2013, 87, 10037–10046. [Google Scholar] [CrossRef]
- Li, Z.; Lou, Y.; Tian, G.; Wu, J.; Lu, A.; Chen, J.; Xu, B.; Shi, J.; Yang, J. Discovering master regulators in hepatocellular carcinoma: One novel MR, SEC14L2 inhibits cancer cells. Aging 2019, 11, 12375–12411. [Google Scholar] [CrossRef]
- Zhu, Q.; Wan, N.B.; Deng, H.W.; Lu, L.L.; Zhang, Y.; He, X.; Liu, H.; He, Y. SEC14L3 plays a tumor-suppressive role in breast cancer through a Wnt/β-catenin-related way. Exp. Cell Res. 2022, 417, 113161. [Google Scholar] [CrossRef]
- Inohara, N.; Nuñez, G. ML—A conserved domain involved in innate immunity and lipid metabolism. Trends Biochem. Sci. 2002, 27, 219–221. [Google Scholar] [CrossRef]
- Shimazu, R.; Akashi, S.; Ogata, H.; Nagai, Y.; Fukudome, K.; Miyake, K.; Kimoto, M. MD-2, a molecule that confers lipopolysaccharide responsiveness on Toll-like receptor 4. J. Exp. Med. 1999, 189, 1777–1782. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, Z.; Lei, Z.; Lei, P. CD14: Biology and role in the pathogenesis of disease. Cytokine Growth Factor. Rev. 2019, 8, 24–31. [Google Scholar] [CrossRef]
- Miyake, K. Innate immune sensing of pathogens and danger signals by cell surface Toll-like receptors. Semin. Immunol. 2007, 19, 3–10. [Google Scholar] [CrossRef]
- Gioannini, T.L.; Weiss, J.P. Regulation of interactions of Gram-negative bacterial endotoxins with mammalian cells. Immunol. Res. 2007, 39, 249–260. [Google Scholar] [CrossRef]
- Nagai, Y.; Akashi, S.; Nagafuku, M.; Ogata, M.; Iwakura, Y.; Akira, S.; Kitamura, T.; Kosugi, A.; Kimoto, M.; Miyake, K. Essential role of MD-2 in LPS responsiveness and TLR4 distribution. Nat. Immunol. 2002, 3, 667–672. [Google Scholar] [CrossRef]
- Ao, J.Q.; Ling, E.; Rao, X.J.; Yu, X.Q. A novel ML protein from Manduca sexta may function as a key accessory protein for lipopolysaccharide signaling. Mol. Immunol. 2008, 45, 2772–2781. [Google Scholar] [CrossRef]
- Liao, J.X.; Yin, Z.X.; Huang, X.D.; Weng, S.P.; Yu, X.Q.; He, J.G. Cloning and characterization of a shrimp ML superfamily protein. Fish Shellfish Immunol. 2011, 30, 713–719. [Google Scholar] [CrossRef]
- Dong, Y.; Aguilar, R.; Xi, Z.; Warr, E.; Mongin, E.; Dimopoulos, G. Anopheles gambiae immune responses to human and rodent Plasmodium parasite species. PLoS Pathog. 2006, 2, e52. [Google Scholar] [CrossRef]
- Zhang, R.N.; Ren, F.F.; Zhou, C.B.; Xu, J.F.; Yi, H.Y.; Ye, M.Q.; Deng, X.J.; Cao, Y.; Yu, X.Q.; Yang, W.Y. An ML protein from the silkworm Bombyx mori may function as a key accessory protein for lipopolysaccharide signaling. Dev. Comp. Immunol. 2018, 88, 94–103. [Google Scholar] [CrossRef]
- Zhang, R.; Li, X.; Zhang, J.; Li, Y.; Wang, Y.; Song, Y.; Ren, F.; Yi, H.; Deng, X.; Zhong, Y.; et al. Toll9 from Bombyx mori functions as a pattern recognition receptor that shares features with Toll-like receptor 4 from mammals. Proc. Natl. Acad. Sci. USA 2021, 118, e2103021118. [Google Scholar] [CrossRef]
- Horáčková, J.; Rudenko, N.; Golovchenko, M.; Havlíková, S.; Grubhoffer, L. IrML—A gene encoding a new member of the ML protein family from the hard tick, Ixodes ricinus. J. Vector Ecol. 2010, 35, 410–418. [Google Scholar] [CrossRef]
- Huang, J.; Ghosh, R.; Bankaitis, V.A. Sec14-like phosphatidylinositol transfer proteins and the biological landscape of phosphoinositide signaling in plants. Biochim. Biophys. Acta 2016, 1861, 1352–1364. [Google Scholar] [CrossRef]
- De Campos, M.K.; Campos, M.; Schaaf, G. The regulation of cell polarity by lipid transfer proteins of the SEC14 family. Curr. Opin. Plant Biol. 2017, 40, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Kiełbowicz-Matuk, A.; Banachowicz, E.; Turska-Tarska, A.; Rey, P.; Rorat, T. Expression and characterization of a barley phosphatidylinositol transfer protein structurally homologous to the yeast Sec14p protein. Plant Sci. 2016, 246, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Hua, M.; Wen, W.; Wei, S.; Ya, S.; Feng, L.; Cong, L.; Ling, W.; Xu, Z.; Li, X.; You, Q. Genome-wide identification, phylogeny, and expression analysis of Sec14-like PITP gene family in sugarcane. Plant. Cell Rep. 2019, 38, 637–655. [Google Scholar]
- Wang, X.; Shan, X.; Xue, C.; Wu, Y.; Su, S.; Li, S.; Liu, H.; Jiang, Y.; Zhang, Y.; Yuan, Y. Isolation and functional characterization of a cold responsive phosphatidylinositol transfer-associated protein, ZmSEC14p, from maize (Zea may L.). Plant Cell Rep. 2016, 35, 1671–1686. [Google Scholar] [CrossRef] [PubMed]
- Kiba, A.; Nakano, M.; Vincent-Pope, P.; Takahashi, H.; Sawasaki, T.; Endo, Y.; Ohnishi, K.; Yoshioka, H.; Hikichi, Y. A novel Sec14 phospholipid transfer protein from Nicotiana benthamiana is up-regulated in response to Ralstonia solanacearum infection, pathogen associated molecular patterns and effector molecules and involved in plant immunity. J. Plant Physiol. 2012, 169, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Kiba, A.; Galis, I.; Hojo, Y.; Ohnishi, K.; Yoshioka, H.; Hikichi, Y. SEC14 phospholipid transfer protein is involved in lipid signaling-mediated plant immune responses in Nicotiana benthamiana. PLoS ONE 2014, 9, e98150. [Google Scholar] [CrossRef]
- Ghosh, R.; de Campos, M.K.; Huang, J.; Huh, S.K.; Orlowski, A.; Yang, Y.; Tripathi, A.; Nile, A.; Lee, H.C.; Dynowski, M.; et al. Sec14-nodulin proteins and the patterning of phosphoinositide landmarks for developmental control of membrane morphogenesis. Mol. Biol. Cell 2015, 26, 1764–1781. [Google Scholar] [CrossRef]
- Zhou, H.; Duan, H.; Liu, Y.; Sun, X.; Zhao, J.; Lin, H. Patellin protein family functions in plant development and stress response. J. Plant Physiol. 2019, 234–235, 94–97. [Google Scholar] [CrossRef]
- Vincent, P.; Chua, M.; Nogue, F.; Fairbrother, A.; Mekeel, H.; Xu, Y.; Allen, N.; Bibikova, T.N.; Gilroy, S.; Bankaitis, V.A. A Sec14p-nodulin domain phosphatidylinositol transfer protein polarizes membrane growth of A. thaliana root hairs. J. Cell Biol. 2005, 168, 801–812. [Google Scholar] [CrossRef]
- Böhme, K.; Li, Y.; Charlot, F.; Grierson, C.; Marrocco, K.; Okada, K.; Laloue, M.; Nogué, F. The Arabidopsis COW1 gene encodes a phosphatidylinositol transfer protein essential for root hair tip growth. Plant J. 2004, 40, 686–698. [Google Scholar] [CrossRef]
- Suzuki, T.; Matsushima, C.; Nishimura, S.; Higashiyama, T.; Sasabe, M.; Machida, Y. Identification of Phosphoinositide-Binding Protein PATELLIN2 as a Substrate of Arabidopsis MPK4 MAP Kinase during Septum Formation in Cytokinesis. Plant. Cell Physiol. 2016, 57, 1744–1755. [Google Scholar] [CrossRef] [PubMed]
- Dalal, J.; Lewis, D.R.; Tietz, O.; Brown, E.M.; Brown, C.S.; Palme, K.; Muday, G.K.; Sederoff, H.W. ROSY1, a novel regulator of gravitropic response is a stigmasterol binding protein. J. Plant Physiol. 2016, 196–197, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Hakenjos, J.P.; Bejai, S.; Ranftl, Q.; Behringer, C.; Vlot, A.C.; Absmanner, B.; Hammes, U.; Heinzlmeir, S.; Kuster, B.; Schwechheimer, C. ML3 is a NEDD8- and ubiquitin-modified protein. Plant Physiol. 2013, 163, 135–149. [Google Scholar] [CrossRef]
- Yeats, T.H.; Howe, K.J.; Matas, A.J.; Buda, G.J.; Thannhauser, T.W.; Rose, J.K. Mining the surface proteome of tomato (Solanum lycopersicum) fruit for proteins associated with cuticle biogenesis. J. Exp. Bot. 2010, 61, 3759–3771. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chen, H.; Fan, J.; Wang, Y.; Li, Y.; Chen, J.; Fan, J.; Yang, S.; Hu, L.; Leung, H.; et al. Genetic diversity and disease control in rice. Nature 2000, 406, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Mallik, S.; Tawfik, D.S.; Levy, E.D. How gene duplication diversifies the landscape of protein oligomeric state and function. Curr. Opin. Genet. Dev. 2022, 76, 101966. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef]
- Fernandez, J.; Orth, K. Rise of a Cereal Killer: The Biology of Magnaporthe oryzae Biotrophic Growth. Trends Microbiol. 2018, 26, 582–597. [Google Scholar] [CrossRef]
- Pennisi, E. Armed and dangerous. Science 2010, 327, 804–805. [Google Scholar]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef] [PubMed]
- KuoChen, C.; HongBin, S. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–428. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.S.; Baek, K.H. Jasmonic Acid Signaling Pathway in Response to Abiotic Stresses in Plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel, M.; Brini, F.; Sharma, A.; Landi, M. Role of jasmonic acid in plants: The molecular point of view. Plant Cell Rep. 2021, 40, 1471–1494. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mostafa, S.; Zeng, W.; Jin, B. Function and Mechanism of Jasmonic Acid in Plant Responses to Abiotic and Biotic Stresses. Int. J. Mol. Sci. 2021, 22, 8568. [Google Scholar] [CrossRef]
- Loake, G.; Grant, M. Salicylic acid in plant defence—the players and protagonists. Curr. Opin. Plant Biol. 2007, 10, 466–472. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic Acid: Biosynthesis and Signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef]
- An, C.; Mou, Z. Salicylic acid and its function in plant immunity. J. Integr. Plant Biol. 2011, 53, 412–428. [Google Scholar] [CrossRef]
- Ding, P.; Ding, Y. Stories of Salicylic Acid: A Plant Defense Hormone. Trends Plant Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Busch, W.; Benfey, P.N. Information processing without brains—The power of intercellular regulators in plants. Development 2010, 137, 1215–1226. [Google Scholar] [CrossRef]
- Van Norman, J.M.; Breakfield, N.W.; Benfey, P.N. Intercellular communication during plant development. Plant Cell 2011, 23, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Oldroyd, G.E. Plant signalling in symbiosis and immunity. Nature 2017, 543, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhou, J.M. Receptor-Like Cytoplasmic Kinases: Central Players in Plant Receptor Kinase-Mediated Signaling. Annu. Rev. Plant Biol. 2018, 69, 267–299. [Google Scholar] [CrossRef]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nürnberger, T.; Jones, J.D.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef]
- Heese, A.; Hann, D.R.; Gimenez-Ibanez, S.; Jones, A.M.; He, K.; Li, J.; Schroeder, J.I.; Peck, S.C.; Rathjen, J.P. The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Proc. Natl. Acad. Sci. USA 2007, 104, 12217–12222. [Google Scholar] [CrossRef]
- Kanda, Y.; Yokotani, N.; Maeda, S.; Nishizawa, Y.; Kamakura, T.; Mori, M. The receptor-like cytoplasmic kinase BSR1 mediates chitin-induced defense signaling in rice cells. Biosci. Biotechnol. Biochem. 2017, 81, 1497–1502. [Google Scholar] [CrossRef]
- Kanda, Y.; Nakagawa, H.; Nishizawa, Y.; Kamakura, T.; Mori, M. Broad-Spectrum Disease Resistance Conferred by the Overexpression of Rice RLCK BSR1 Results from an Enhanced Immune Response to Multiple MAMPs. Int. J. Mol. Sci. 2019, 20, 5523. [Google Scholar] [CrossRef]
- Kiba, A.; Nakano, M.; Ohnishi, K.; Hikichi, Y. The SEC14 phospholipid transfer protein regulates pathogen-associated molecular pattern-triggered immunity in Nicotiana benthamiana. Plant. Physiol. Biochem. 2018, 125, 212–218. [Google Scholar] [CrossRef]
- Peiro, A.; Izquierdo-Garcia, A.C.; Sanchez-Navarro, J.A.; Pallas, V.; Mulet, J.M.; Aparicio, F. Patellins 3 and 6, two members of the Plant Patellin family, interact with the movement protein of Alfalfa mosaic virus and interfere with viral movement. Mol. Plant Pathol. 2014, 15, 881–891. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, J.R. Effectors and effector delivery in Magnaporthe oryzae. PLoS Pathog. 2014, 10, e1003826. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Liu, J.; Ning, Y.; Ding, B.; Wang, X.; Wang, Z.; Wang, G.L. Recent progress in understanding PAMP- and effector-triggered immunity against the rice blast fungus Magnaporthe oryzae. Mol. Plant 2013, 6, 605–620. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Gene ID | Amino Acid Number | Molecular Weight | Theoretical pI | Instability Index | Aliphatic Index | GRAVY |
|---|---|---|---|---|---|---|---|
| OsPitpα | LOC_Os03g17800.1 | 195 | 22,111.60 | 8.40 | 39.63 | 87.54 | −0.293 |
| OsSec14-1 | LOC_Os01g16000.1 | 318 | 36,127.60 | 5.896 | 41.42 | 68.71 | −0.643 |
| OsSec14-2 | LOC_Os01g50616.1 | 671 | 75,911.99 | 9.02 | 53.38 | 84.43 | −0.556 |
| OsSec14-3 | LOC_Os01g65380.1 | 613 | 66,457.94 | 4.65 | 55.45 | 74.81 | −0.453 |
| OsSec14-4 | LOC_Os01g70210.1 | 247 | 28,078.39 | 8.24 | 42.17 | 86.11 | −0.366 |
| OsSec14-5 | LOC_Os02g04020.1 | 624 | 71,010.26 | 8.47 | 50.20 | 82.34 | −0.527 |
| OsSec14-6 | LOC_Os02g04030.1 | 637 | 71,116.11 | 7.19 | 54.24 | 70.50 | −0.532 |
| OsSec14-7 | LOC_Os02g10650.1 | 327 | 37,028.52 | 6.87 | 39.43 | 89.94 | −0.341 |
| OsSec14-8 | LOC_Os02g24430.1 | 559 | 63,004.79 | 7.88 | 49.62 | 82.72 | −0.404 |
| OsSec14-9 | LOC_Os02g48990.1 | 612 | 69,750.04 | 6.49 | 43.21 | 79.31 | −0.622 |
| OsSec14-10 | LOC_Os02g51610.1 | 378 | 42,355.55 | 5.16 | 50.09 | 70.77 | −0.636 |
| OsSec14-11 | LOC_Os03g51430.1 | 299 | 34,667.65 | 8.55 | 51.96 | 78.90 | −0.528 |
| OsSec14-12 | LOC_Os03g63370.1 | 261 | 30,159.22 | 8.20 | 46.17 | 77.39 | −0.621 |
| OsSec14-13 | LOC_Os05g18470.1 | 283 | 32,973.69 | 8.88 | 48.97 | 71.38 | −0.616 |
| OsSec14-14 | LOC_Os05g27820.1 | 435 | 47,828.78 | 8.27 | 44.09 | 83.29 | −0.183 |
| OsSec14-15 | LOC_Os05g35460.1 | 585 | 63,948.04 | 4.59 | 48.61 | 77.85 | −0.457 |
| OsSec14-16 | LOC_Os05g46720.1 | 613 | 70,003.75 | 8.34 | 51.93 | 73.03 | −0.621 |
| OsSec14-17 | LOC_Os06g40510.1 | 330 | 37,299.38 | 5.67 | 37.85 | 92.70 | −0.377 |
| OsSec14-18 | LOC_Os06g45990.2 | 450 | 49,793.09 | 4.83 | 52.01 | 76.80 | −0.517 |
| OsSec14-19 | LOC_Os07g27310.1 | 418 | 47,494.42 | 8.55 | 45.03 | 88.90 | −0.427 |
| OsSec14-20 | LOC_Os08g25310.1 | 637 | 73,245.23 | 8.91 | 62.16 | 72.70 | −0.669 |
| OsSec14-21 | LOC_Os08g38850.1 | 604 | 69,256.46 | 6.86 | 55.89 | 77.95 | −0.595 |
| OsSec14-22 | LOC_Os09g08390.1 | 335 | 37,958.67 | 7.65 | 36.43 | 85.55 | −0.331 |
| OsSec14-23 | LOC_Os09g30330.1 | 551 | 63,232.14 | 7.29 | 63.43 | 75.34 | 0.507 |
| OsSec14-24 | LOC_Os10g03400.2 | 598 | 67,600.15 | 8.49 | 54.00 | 77.01 | −0.484 |
| OsSec14-25 | LOC_Os02g21630.1 | 261 | 29,758.36 | 8.86 | 44.11 | 86.55 | −0.248 |
| OsSec14-26 | LOC_Os03g11950.1 | 438 | 48,675.95 | 9.95 | 50.69 | 78.45 | −0.219 |
| OsSec14-27 | LOC_Os05g18294.1 | 188 | 21,704.77 | 6.39 | 53.41 | 89.20 | −0.384 |
| OsMl-1 | LOC_Os07g06590.1 | 152 | 16,196.64 | 5.32 | 36.31 | 89.87 | 0.215 |
| OsMl-2 | LOC_Os03g57420.1 | 156 | 16,946.55 | 5.66 | 40.16 | 96.86 | 0.238 |
| Protein Name | CELLO | Plant-mPLoc | Protein Name | CELLO | Plant-mPLoc |
|---|---|---|---|---|---|
| OsPitpα | Nucleus | Nucleus | OsSec14-15 | Nucleus | Cytoplasm |
| OsSec14-1 | Nucleus | Cytoplasm | OsSec14-16 | Chloroplast | Cytoplasm |
| OsSec14-2 | Nucleus | Cytoplasm | OsSec14-17 | Nucleus | Cytoplasm |
| OsSec14-3 | Nucleus | Nucleus | OsSec14-18 | Nucleus | Cytoplasm |
| OsSec14-4 | Cytoplasm | Cytoplasm | OsSec14-19 | Mitochondrio | Cytoplasm |
| OsSec14-5 | Chloroplast | Cytoplasm | OsSec14-20 | Nucleus | Cytoplasm |
| OsSec14-6 | Chloroplast | Cytoplasm | OsSec14-21 | Chloroplast | Cytoplasm |
| OsSec14-7 | Cytoplasm | Cytoplasm | OsSec14-22 | Cytoplasm | Cytoplasm |
| OsSec14-8 | Cytoplasm | Cytoplasm | OsSec14-23 | Mitochondrio | Cytoplasm |
| OsSec14-9 | Chloroplast | Cytoplasm | OsSec14-24 | Chloroplast | Cytoplasm |
| OsSec14-10 | Cytoskeleton | Cytoplasm | OsSec14-25 | Cytoplasm | Cytoplasm |
| OsSec14-11 | Nucleus | Cytoplasm | OsSec14-26 | Cytoplasm | Cytoplasm |
| OsSec14-12 | Cytoplasm | Cytoplasm | OsSec14-27 | Cytoplasm | Cytoplasm |
| OsSec14-13 | Cytoplasm | Cytoplasm | OsMl-1 | Cytoplasm | Nucleus |
| OsSec14-14 | Chloroplast | Cytoplasm | OsMl-2 | Extracellular | Nucleus |
| Primer Name | Primer Sequence (5′-3′) | Exon-Exon Junction | qRT-PCR Products (bp) |
|---|---|---|---|
| QRT-OsACTIN-F | GGACTCTGGTGATGGTGTCAGCCA | Yes | 243 |
| QRT-OsACTIN-R | GAGCTGGTCTTGGCAGTCTCCA | ||
| QRT-OsML-1-F | GTGGTGACTTCTTGGTAG | Yes | 76 |
| QRT-OsML-1-R | TTCATGGTGATGGTGTAAG | ||
| QRT-OsML-2-F | CACGAACAGACTTTACCA | Yes | 75 |
| QRT-OsML-2-R | GTTGCCGTCATCTAGTAG | ||
| QRT-OsSEC14-3-F | ACCGTTGAGATTCCTGTC | Yes | 89 |
| QRT-OsSEC14-3-R | GTGAACTCTGCTCCGTAG | ||
| QRT-OsSEC14-4-F | TGGAAGATGATCTACCCTTTC | Yes | 103 |
| QRT-OsSEC14-4-R | TATCGTCGATCTCCTGGT | ||
| QRT-OsSEC14-5-F | TTCAAGATAAGCCTTCAG | Yes | 125 |
| QRT-OsSEC14-5-R | GATAAGAGCCTCATGTAG | ||
| QRT-OsSEC14-15-F | AACTGTTGAGATTCCTGCTA | Yes | 90 |
| QRT-OsSEC14-15-R | GTGAACTCTGCACCGTAG | ||
| QRT-OsSEC14-16-F | GAGATGCCATTTGAAAAG | Yes | 183 |
| QRT-OsSEC14-16-R | CTAGAAGCAGAATTTCTTT | ||
| QRT-OsSEC14-19-F | TACTACAGAAGCAGAATG | Yes | 107 |
| QRT-OsSEC14-19-R | TCACTTCAAGATTAGGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, M.; Xie, X.; Peng, B.; Chen, X.; Chen, Y.; Li, Y.; Wang, Z.; Lu, G. Identification and Expression Analysis of Phosphatidylinositol Transfer Proteins Genes in Rice. Plants 2023, 12, 2122. https://doi.org/10.3390/plants12112122
Pei M, Xie X, Peng B, Chen X, Chen Y, Li Y, Wang Z, Lu G. Identification and Expression Analysis of Phosphatidylinositol Transfer Proteins Genes in Rice. Plants. 2023; 12(11):2122. https://doi.org/10.3390/plants12112122
Chicago/Turabian StylePei, Mengtian, Xuze Xie, Baoyi Peng, Xinchi Chen, Yixuan Chen, Ya Li, Zonghua Wang, and Guodong Lu. 2023. "Identification and Expression Analysis of Phosphatidylinositol Transfer Proteins Genes in Rice" Plants 12, no. 11: 2122. https://doi.org/10.3390/plants12112122