

A PHLID Model for Tomato Bacterial Canker Predicting on Epidemics of the Pathogen


Abstract
:1. Introduction
2. Materials and Methods
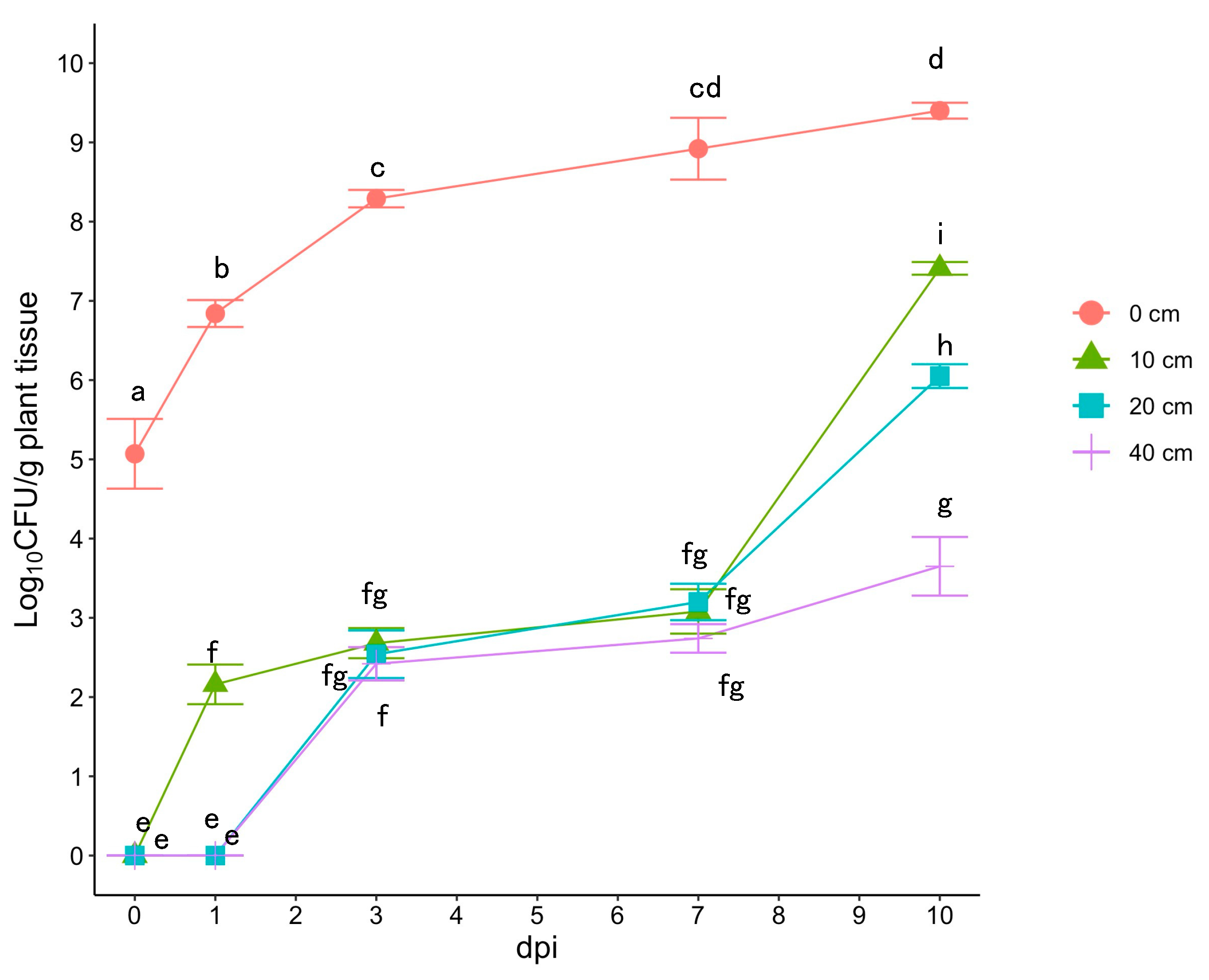
2.1. Incubation Period of Cmm in Latently Infected Tomato Plants
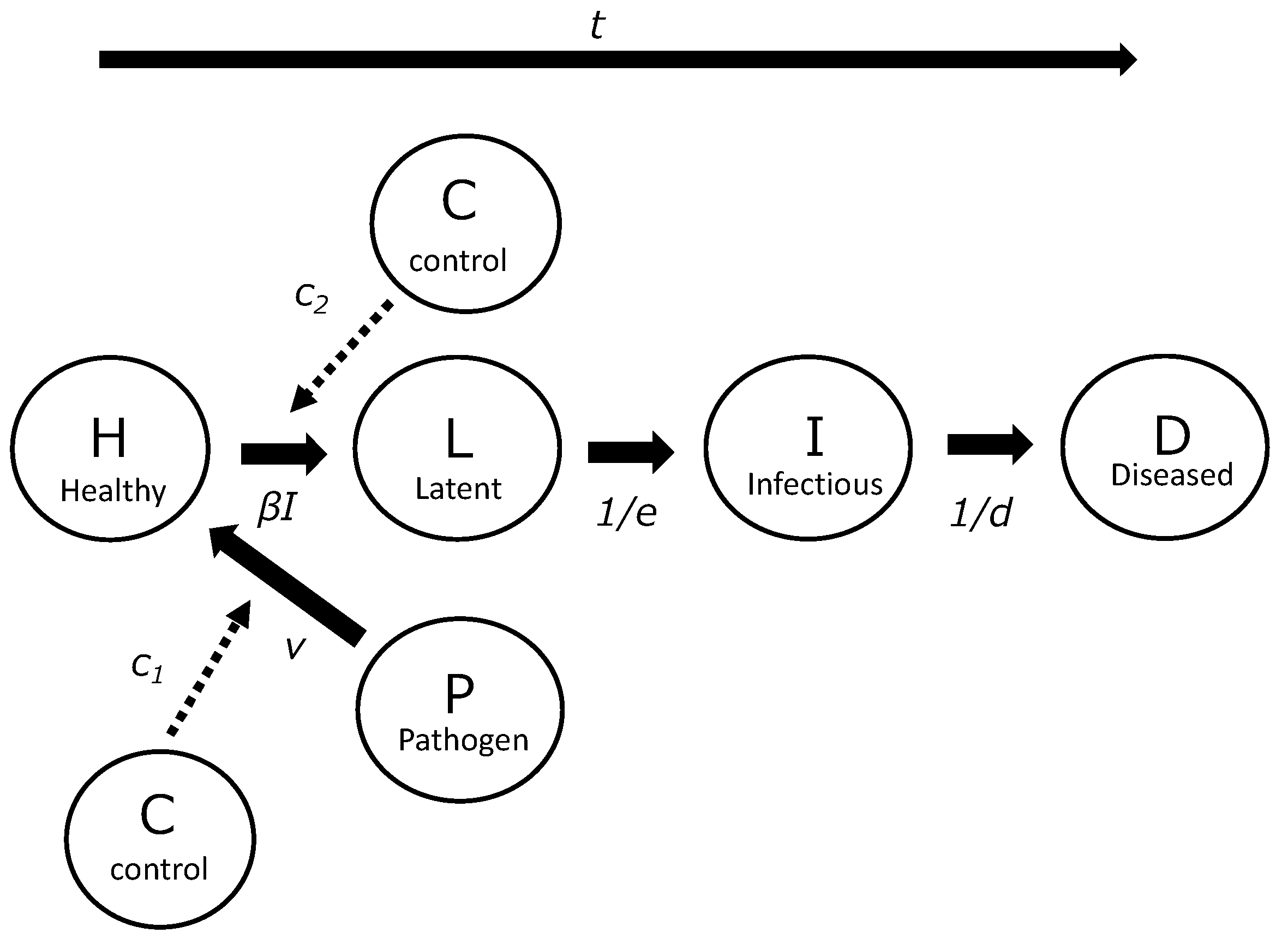
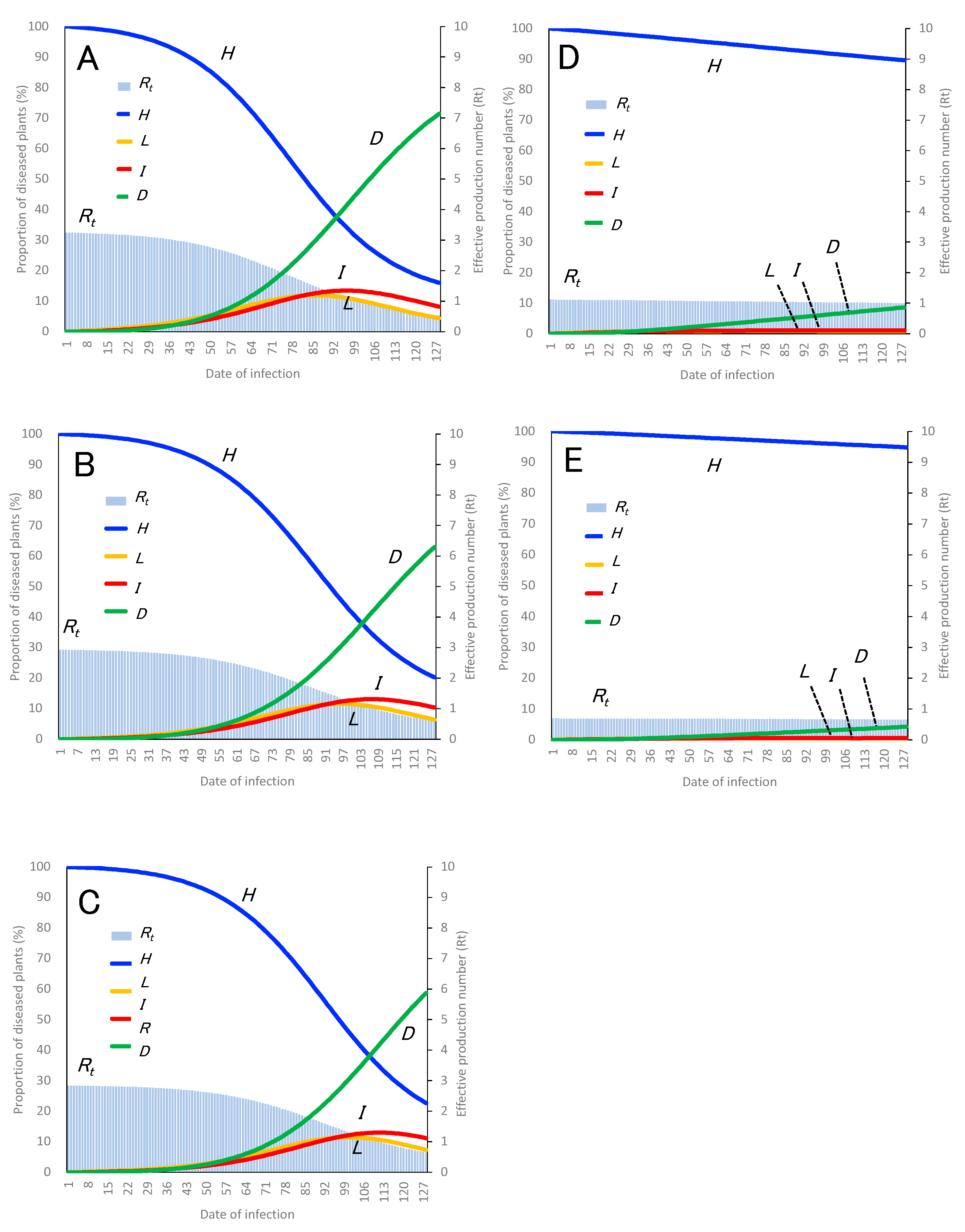
2.2. PHLID Model, Basic Reproduction Number (R0), and Effective Reproduction Number (Rt)
3. Results
3.1. Population Dynamics of Cmm in Latently Infected Tomato Plants and Incubation Period
3.2. Development of PHLID Model
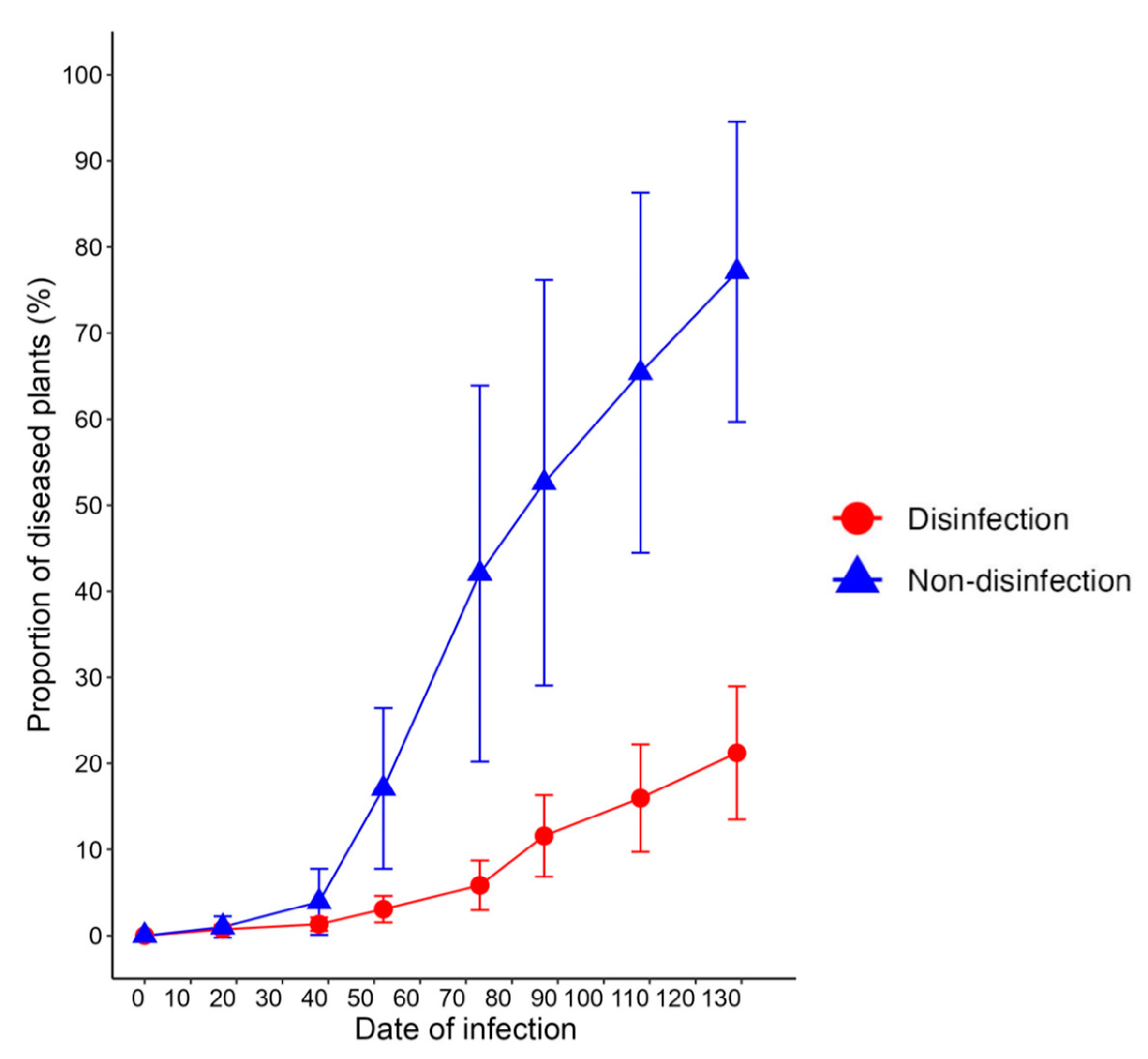
3.3. Estimating Control Effects Using PHLID Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de León, L.; Siverio, F.; López, M.M.; Rodríguez, A. Clavibacter michiganensis subsp. michiganensis, a seedborne tomato pathogen: Healthy seeds are still the goal. Plant Dis. 2011, 95, 1328–1338. [Google Scholar] [PubMed]
- Kawaguchi, A.; Tanina, K.; Inoue, K. Molecular typing and spread of Clavibacter michiganensis subsp. michiganensis in greenhouses in Japan. Plant Pathol. 2010, 59, 76–83. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Kitabayashi, S.; Inoue, K.; Tanina, K. An HLD Model for Tomato Bacterial Canker Focusing on Epidemics of the Pathogen Due to Cutting by Infected Scissors. Plants 2022, 11, 2253. [Google Scholar] [CrossRef] [PubMed]
- Gleason, M.L.; Gitaitis, R.D.; Ricker, M.D. Recent progress in understanding and controlling bacterial canker of tomato in Eastern North America. Plant Dis. 1993, 77, 1069–1076. [Google Scholar] [CrossRef]
- Jahr, H.; Bahro, R.; Bruger, A.; Ahlemeyer, J.; Eichenlaub, R. Interactions between Clavibacter michiganensis and its host plants. Environ. Microbiol. 1999, 1, 113–118. [Google Scholar] [CrossRef]
- Peritore-Galve, F.C.; Tancos, M.A.; Smart, C.D. Bacterial canker of tomato: Revisiting a global and economically damaging seedborne pathogen. Plant Dis. 2021, 105, 1581–1595. [Google Scholar] [CrossRef]
- Fatmi, M.; Schaad, N.W. Survival of Clavibacter michiganensis ssp. michiganensis in infected tomato stems under natural field conditions in California, Ohio and Morocco. Plant Pathol. 2002, 51, 149–154. [Google Scholar]
- Grogan, R.G.; Kimble, K.A. The role of seed contamination in the transmission of Pseudomonas phaseolicola in Phaseolus vulgaris. Phytopathology 1967, 57, 28–31. [Google Scholar]
- Kawaguchi, A.; Tanina, K. Genetic groups of Clavibacter michiganensis subsp. michiganensis identified by DNA fingerprinting and the effects of inoculation methods on disease development. Euro. J. Plant Pathol. 2014, 140, 399–406. [Google Scholar] [CrossRef]
- Gitaitis, R.D. Bacterial canker. In Compendium of Tomato Diseases; Jones, J.B., Jones, J.P., Stall, R.E., Zitter, T.A., Eds.; APS Press: St. Paul, MN, USA, 1991; pp. 25–26. [Google Scholar]
- Kawaguchi, A.; Tanina, K.; Inoue, K. Spatiotemporal distribution of tomato plants naturally infected with bacterial canker in greenhouses. J. Gen. Plant Pathol. 2013, 79, 46–50. [Google Scholar] [CrossRef]
- Sharabani, G.; Manulis-Sasson, S.; Borenstein, M.; Shulhani, R.; Lofthouse, M.; Chalupowicz, L.; Shtienberg, D. The significance of guttation in the secondary spread of Clavibacter michiganensis subsp. michiganensis in tomato greenhouses. Plant Pathol. 2013, 62, 578–586. [Google Scholar] [CrossRef]
- Sen, Y.; van der Wolf, J.; Visser, R.G.F.; van Heusden, S. Bacterial canker of tomato: Current knowledge of detection, management, resistance, and interactions. Plant Dis. 2015, 99, 4–13. [Google Scholar] [CrossRef]
- Kermack, W.O.; McKendrick, A.G. A contribution to the mathematical theory of epidemics. Proc. R. Soc. London Ser. A 1927, 115, 700–721. [Google Scholar] [CrossRef]
- Madden, L.V. Botanical epidemiology: Some key advances and its continuing role in disease management. Eur. J. Plant Pathol. 2006, 115, 3–23. [Google Scholar] [CrossRef]
- Madden, L.V.; Van den Bosch, F. A population-dynamics approach to assess the threat of plant pathogens as biological weapons against annual crops. BioScience 2002, 52, 65–74. [Google Scholar] [CrossRef]
- Gilligan, C.A.; Gubbins, S. Analysis and fitting of an SIR model with host response to infection load for a plant disease. Philos. Trans. R. Soc. B Biol. Sci. 1997, 352, 353–364. [Google Scholar] [CrossRef]
- Ozcaglar, C.; Shabbeer, A.; Vandenberg, S.L.; Yener, B.; Bennett, K.P. Epidemiological models of mycobacterium tuberculosis complex infections. Math. Biosci. 2012, 236, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Hethcote, H.W. The mathematics of infectious diseases. Soc. Ind. Appl. Math. Siam Rev. 2000, 42, 599–653. [Google Scholar] [CrossRef]
- Shinta, R.; Arsal, S.R.; Aldila, D.; Handari, B.D. Short review of mathematical model of measles. AIP Conf. Proc. 2020, 2264, 020003. [Google Scholar] [CrossRef]
- Shirakawa, T.; Sasaki, T. A selective medium for isolation of Corynebacterium michiganense pv. michiganense, the pathogen of tomato bacterial canker disease. Ann. Phytopathol. Soc. Jpn. 1988, 54, 540–543. [Google Scholar] [CrossRef]
- Tanina, K.; Kawaguchi, A. Simplified method to isolate Clavibacter michiganensis subsp. michiganensis using ImmunoStripTMCmm. Jpn. J. Phytopathol. 2011, 77, 287–290. [Google Scholar] [CrossRef]
- Hadasa, R.; Kritzmanb, G.; Klietmanb, F.; Gefenc, T.; Manulis, S. Comparison of extraction procedures and determination of the detection threshold for Clavibacter michiganensis ssp. michiganensis in tomato seeds. Plant Pathol. 2005, 54, 643–649. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Tanina, K.; Inoue, K. Effect of soil iisinfestation and prevention of contagious infection against tomato bacterial canker in greenhouses. Kinki Chugoku Shikoku Agric. Res. 2011, 18, 13–17. [Google Scholar]
- Adam, D. A guide to R—The pandemic’s misunderstood metric. Nature 2020, 583, 346–348. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Nanaumi, T. Model-based forecasting of twig cankers incidence of bacterial spot of peach in Fukushima Prefecture. J. Gen. Plant Pathol. 2022, 88, 41–47. [Google Scholar] [CrossRef]
- Kawaguchi, A. Risk factors for bacterial spot on peach in Okayama Prefecture, Japan. J. Gen. Plant Pathol. 2014, 80, 435–442. [Google Scholar] [CrossRef]
- Kawaguchi, A. Risk factors and assessments for bacterial black node on barley. J. Gen. Plant Pathol. 2020, 86, 193–198. [Google Scholar] [CrossRef]
- Kawaguchi, A. Model-based forecasting of bacterial black node of barley by hierarchical Bayesian model. J. Gen. Plant Pathol. 2022, 88, 48–54. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, S.; Li, Z.; Yang, W. Advances in the Characterization of the Mechanism Underlying Bacterial Canker Development and Tomato Plant Resistance. Horticulturae 2022, 8, 209. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Variable | Reference |
|---|---|---|---|
| B | Infection rate per contact (inoculated 106 cells/mL of Cmm) | 0.706 | [3] |
| k | Number of contacts per day | 0.286 | [3] |
| d | Duration of infection (days) | 12 | [3] |
| 1/e | Incubation period (days) | 10 | Figure 2 |
| vsoil | Infection rate by infected seeds as primary inoculum | 0.067 | [9] |
| vseed | Infection rate by infected seeds as primary inoculum | 0.04 | [23] |
| c1 | Integrated relative risk of soil disinfection from five independent field experiments | 0.48 | [24] |
| c2 | Integrated relative risk of scissors disinfection from four independent field experiments | 0.12 | [3] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawaguchi, A.; Kitabayashi, S.; Inoue, K.; Tanina, K. A PHLID Model for Tomato Bacterial Canker Predicting on Epidemics of the Pathogen. Plants 2023, 12, 2099. https://doi.org/10.3390/plants12112099
Kawaguchi A, Kitabayashi S, Inoue K, Tanina K. A PHLID Model for Tomato Bacterial Canker Predicting on Epidemics of the Pathogen. Plants. 2023; 12(11):2099. https://doi.org/10.3390/plants12112099
Chicago/Turabian StyleKawaguchi, Akira, Shoya Kitabayashi, Koji Inoue, and Koji Tanina. 2023. "A PHLID Model for Tomato Bacterial Canker Predicting on Epidemics of the Pathogen" Plants 12, no. 11: 2099. https://doi.org/10.3390/plants12112099