
Cereal–Pea Intercropping Reveals Variability in the Relationships among Yield, Quality Parameters, and Obligate Pathogens Infection in Wheat, Rye, Oat, and Triticale, in a Temperate Environment

Abstract
:1. Introduction
2. Results
2.1. Difference in Effects of Year and Cultivation Practices on Yield and TKW of Cereal Crops
2.2. Potentials of Cereal–Pea Intercropping Systems for Regulating Powdery Mildew and Leaf Rust Infections
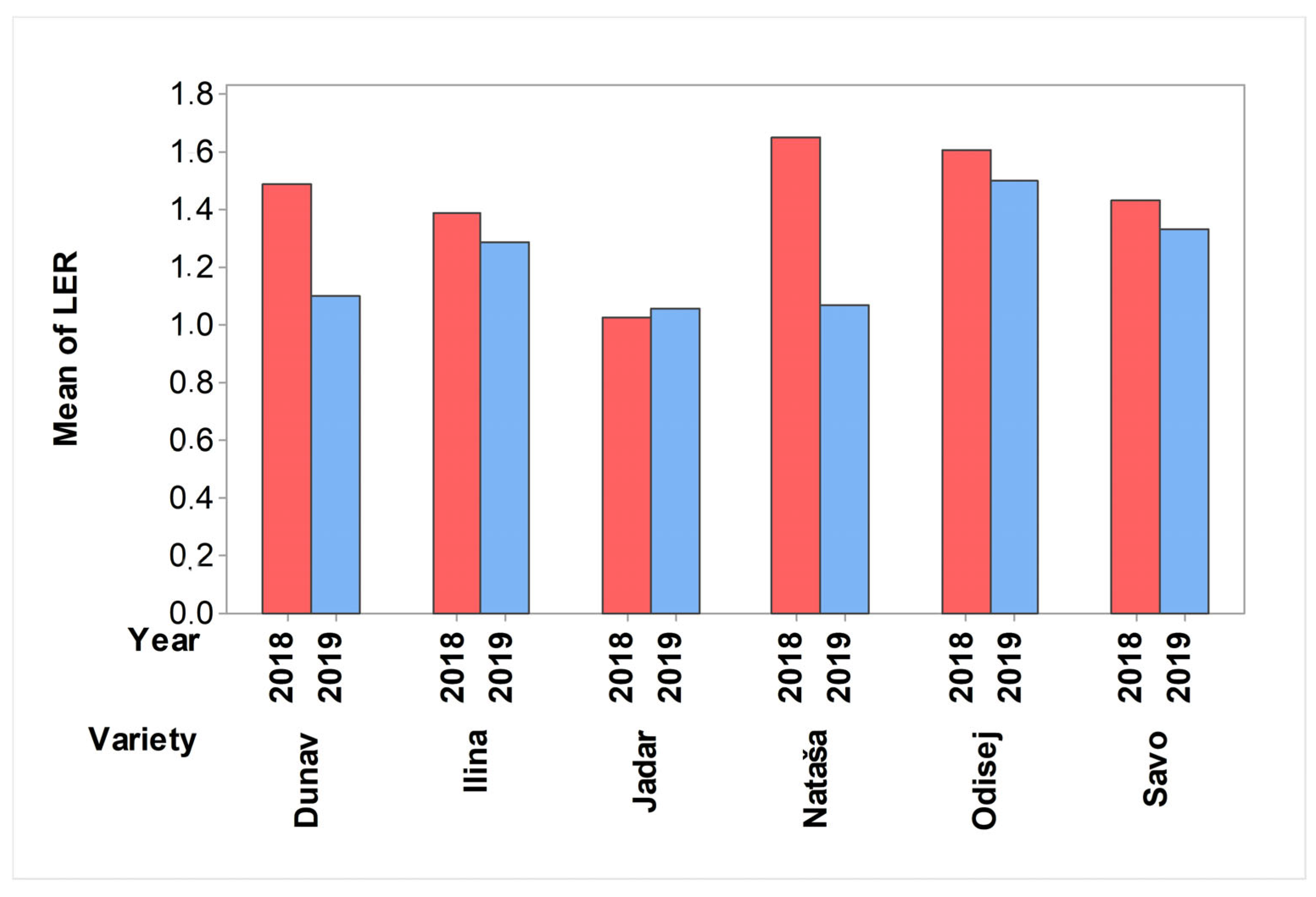
2.3. Effect of Intercropping Systems on Nitrogen Usage of Cereal Crops and Contribution of Cereal Crops to the Land Equivalent Ratio
3. Discussion
3.1. The Variable Effects of Year and Cultivation Practices on Yields and TKWs of Cereal Crops
3.2. Potential of Cereal–Pea Intercropping Systems for Regulating Powdery Mildew and Leaf Rust Infection
3.3. Effect of Intercropping Systems on Nitrogen Usage of Cereal Crops and Contribution of Cereal Crops to the Land Equivalent Ratio
4. Materials and Methods
4.1. Disease Assessments
4.2. Yield and Crude Protein
4.3. Land Equivalent Ratio (LER)
4.4. Climatic Conditions
4.5. Statistical Methods
5. Conclusions
- The effect of intercropping on yield and TKW was not straightforward. The changes in TKW in a single variety cultivated using different practices were not dependent on the year to the same extent as the yield.
- Intercropping could decrease the yield variation under extreme fluctuations of climatic factors during the flowering and grain-filling periods.
- The relationships between the level of pathogenic infection and yield results were not straightforward and were highly dependent on the yield potentials of the cultivars.
- The contributions of the cereal crops to the LER differed within and across the years.
- Year, variety, cultivation practices, and their interactions (year × variety × cultivation) were all determined as significantly influencing factors (p < 0.001) on the crude protein of cereal crops.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, W.; Piper, J. The Necessary Marriage Between Ecology and Agriculture. Ecology 1989, 70, 1591–1593. [Google Scholar] [CrossRef]
- Bedoussac, L.; Justes, E. A Comparison of Commonly Used Indices for Evaluating Species Interactions and Intercrop Efficiency: Application to Durum Wheat–Winter Pea Intercrops. Field Crops Res. 2011, 124, 25–36. [Google Scholar] [CrossRef]
- Jensen, E.S.; Carlsson, G.; Hauggaard-Nielsen, H. Intercropping of Grain Legumes and Cereals Improves the Use of Soil N Resources and Reduces the Requirement for Synthetic Fertilizer N: A Global-Scale Analysis. Agron. Sustain. Dev. 2020, 40, 5. [Google Scholar] [CrossRef]
- Timaeus, J.; Weedon, O.D.; Finckh, M.R. Harnessing the Potential of Wheat-Pea Species Mixtures: Evaluation of Multifunctional Performance and Wheat Diversity. Front. Plant Sci. 2022, 13, 683. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT 2018. Available online: http://www.fao.org/faostat/en/#home (accessed on 15 January 2023).
- Pelzer, E.; Bazot, M.; Makowski, D.; Corre-Hellou, G.; Naudin, C.; Al Rifaï, M.; Baranger, E.; Bedoussac, L.; Biarnès, V.; Boucheny, P.; et al. Pea–Wheat Intercrops in Low-Input Conditions Combine High Economic Performances and Low Environmental Impacts. Eur. J. Agron. 2012, 40, 39–53. [Google Scholar] [CrossRef]
- Brooker, R.W.; Bennett, A.E.; Cong, W.-F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.M.; Jones, H.G.; Karley, A.J.; et al. Improving Intercropping: A Synthesis of Research in Agronomy, Plant Physiology and Ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef]
- Østergård, H.; Finckh, M.R.; Fontaine, L.; Goldringer, I.; Hoad, S.P.; Kristensen, K.; Lammerts van Bueren, E.T.; Mascher, F.; Munk, L.; Wolfe, M.S. Time for a Shift in Crop Production: Embracing Complexity through Diversity at All Levels. J. Sci. Food Agric. 2009, 89, 1439–1445. [Google Scholar] [CrossRef]
- Finckh, M.; Junge, S.; Schmidt, J.; Šišić, A.; Weedon, O. Lntra- and Interspecific Diversity—The Cornerstones of Agroecological Crop Health Management. In Intercropping for Sustainability: Research Developments and Their Application; Association of Applied Biologists: Warwick, UK, 2021; pp. 193–206. [Google Scholar]
- El Titi, A.; Boller, E.F.; Gendrier, J.P. Integrated Production. Principles and Technical Guidelines. IOBC/WPRS Bull. 1993, 16, 13–38. [Google Scholar]
- Altieri, M.A. The Ecological Role of Biodiversity in Agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef]
- Malézieux, E.; Crozat, Y.; Dupraz, C.; Laurans, M.; Makowski, D.; Ozier-Lafontaine, H.; Rapidel, B.; de Tourdonnet, S.; Valantin-Morison, M. Mixing Plant Species in Cropping Systems: Concepts, Tools and Models. A Review. Agron. Sustain. Dev. 2009, 29, 43–62. [Google Scholar] [CrossRef]
- Yin, W.; Chai, Q.; Zhao, C.; Yu, A.; Fan, Z.; Hu, F.; Fan, H.; Guo, Y.; Coulter, J.A. Water Utilization in Intercropping: A Review. Agric. Water Manag. 2020, 241, 106335. [Google Scholar] [CrossRef]
- Stefan, L.; Hartmann, M.; Engbersen, N.; Six, J.; Schöb, C. Positive Effects of Crop Diversity on Productivity Driven by Changes in Soil Microbial Composition. Front. Microbiol. 2021, 12, 660749. [Google Scholar] [CrossRef]
- Watiki, J.M.; Fukai, S.; Banda, J.A.; Keating, B.A. Radiation Interception and Growth of Maize/Cowpea Intercrop as Affected by Maize Plant Density and Cowpea Cultivar. Field Crops Res. 1993, 35, 123–133. [Google Scholar] [CrossRef]
- Jahansooz, M.R.; Yunusa, I.A.M.; Coventry, D.R.; Palmer, A.R.; Eamus, D. Radiation- and Water-Use Associated with Growth and Yields of Wheat and Chickpea in Sole and Mixed Crops. Eur. J. Agron. 2007, 26, 275–282. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Gooding, M.; Ambus, P.; Corre-Hellou, G.; Crozat, Y.; Dahlmann, C.; Dibet, A.; von Fragstein, P.; Pristeri, A.; Monti, M.; et al. Pea–Barley Intercropping for Efficient Symbiotic N2-Fixation, Soil N Acquisition and Use of Other Nutrients in European Organic Cropping Systems. Field Crops Res. 2009, 113, 64–71. [Google Scholar] [CrossRef]
- Yu, Y.; Stomph, T.-J.; Makowski, D.; Zhang, L.; van der Werf, W. A Meta-Analysis of Relative Crop Yields in Cereal/Legume Mixtures Suggests Options for Management. Field Crops Res. 2016, 198, 269–279. [Google Scholar] [CrossRef]
- Trenbath, B.R. Intercropping for the Management of Pests and Diseases. Intercrop. Bases Product. 1993, 34, 381–405. [Google Scholar] [CrossRef]
- Anil; Park; Phipps; Miller Temperate Intercropping of Cereals for Forage: A Review of the Potential for Growth and Utilization with Particular Reference to the UK. Grass Forage Sci. 1998, 53, 301–317. [CrossRef]
- Finckh, M.R.; Gacek, E.S.; Goyeau, H.; Lannou, C.; Merz, U.; Mundt, C.C.; Munk, L.; Nadziak, J.; Newton, A.C.; de Vallavieille-Pope, C.; et al. Cereal Variety and Species Mixtures in Practice, with Emphasis on Disease Resistance. Agronomie 2000, 20, 813–837. [Google Scholar] [CrossRef]
- Banik, P.; Midya, A.; Sarkar, B.K.; Ghose, S.S. Wheat and Chickpea Intercropping Systems in an Additive Series Experiment: Advantages and Weed Smothering. Eur. J. Agron. 2006, 24, 325–332. [Google Scholar] [CrossRef]
- Lithourgidis, A.S.; Vasilakoglou, I.B.; Dhima, K.V.; Dordas, C.A.; Yiakoulaki, M.D. Forage Yield and Quality of Common Vetch Mixtures with Oat and Triticale in Two Seeding Ratios. Field Crops Res. 2006, 99, 106–113. [Google Scholar] [CrossRef]
- Kontturi, M.; Laine, A.; Niskanen, M.; Hurme, T.; Hyövelä, M.; Peltonen-Sainio, P. Pea–Oat Intercrops to Sustain Lodging Resistance and Yield Formation in Northern European Conditions. Acta Agric. Scand. Sect. B Soil Plant Sci. 2011, 61, 612–621. [Google Scholar] [CrossRef]
- Podgórska-Lesiak, M.; Sobkowicz, P. Prevention of Pea Lodging by Intercropping Barley with Peas at Different Nitrogen Fertilization Levels. Field Crops Res. 2013, 149, 95–104. [Google Scholar] [CrossRef]
- Boudreau, M.A. Diseases in Intercropping Systems. Annu. Rev. Phytopathol. 2013, 51, 499–519. [Google Scholar] [CrossRef] [PubMed]
- Bedoussac, L.; Journet, E.-P.; Hauggaard-Nielsen, H.; Naudin, C.; Corre-Hellou, G.; Jensen, E.S.; Prieur, L.; Justes, E. Ecological Principles Underlying the Increase of Productivity Achieved by Cereal-Grain Legume Intercrops in Organic Farming. A Review. Agron. Sustain. Dev. 2015, 35, 911–935. [Google Scholar] [CrossRef]
- Connor, D.; Hall, A.; Sadras, V. Effect of Nitrogen Content on the Photosynthetic Characteristics of Sunflower Leaves. Funct. Plant Biol. 1993, 20, 251–263. [Google Scholar] [CrossRef]
- Jevtić, R.; Župunski, V.; Lalošević, M.; Jocković, B.; Orbović, B.; Ilin, S. Diversity in Susceptibility Reactions of Winter Wheat Genotypes to Obligate Pathogens under Fluctuating Climatic Conditions. Sci. Rep. 2020, 10, 19608. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Collins, R.P.; De Ron, A.M.; Firmat, C.; Litrico, I.; Hauggaard-Nielsen, H. Chapter Three—Do We Need Specific Breeding for Legume-Based Mixtures? In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 157, pp. 141–215. ISBN 0065-2113. [Google Scholar]
- Malagoli, P.; Naudin, C.; Vrignon-Brenas, S.; Sester, M.; Jeuffroy, M.-H.; Corre-Hellou, G. Modelling Nitrogen and Light Sharing in Pea-Wheat Intercrops to Design Decision Rules for N Fertilisation According to Farmers’ Expectations. Field Crops Res. 2020, 255, 107865. [Google Scholar] [CrossRef]
- Junk, J.; Kouadio, L.; Delfosse, P.; El Jarroudi, M. Effects of Regional Climate Change on Brown Rust Disease in Winter Wheat. Clim. Chang. 2016, 135, 439–451. [Google Scholar] [CrossRef]
- Jevtić, R.; Maširević, S.; Vajgand, D. The Impact of Climate Change on Diseases and Pests of Small Grains and Sunflowers in the Vojvodina Region (Serbia). In Essays on Fundamental and Applied Environmental Topics; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2012; pp. 277–305. [Google Scholar]
- Hauggaard-Nielsen, H.; Andersen, M.K.; Jørnsgaard, B.; Jensen, E.S. Density and Relative Frequency Effects on Competitive Interactions and Resource Use in Pea–Barley Intercrops. Field Crops Res. 2006, 95, 256–267. [Google Scholar] [CrossRef]
- Li, C.; He, X.; Zhu, S.; Zhou, H.; Wang, Y.; Li, Y.; Yang, J.; Fan, J.; Yang, J.; Wang, G.; et al. Crop Diversity for Yield Increase. PLoS ONE 2009, 4, e8049. [Google Scholar] [CrossRef] [PubMed]
- Jevtić, R.; Župunski, V.; Lalošević, M.; Mladenov, N.; Orbović, B. The Combined Effects of Multiple Diseases and Climatic Conditions on Thousand Kernel Weight Losses in Winter Wheat. Eur. J. Plant Pathol. 2018, 152, 469–477. [Google Scholar] [CrossRef]
- Herrera-Foessel, S.A.; Singh, R.P.; Huerta-Espino, J.; Crossa, J.; Yuen, J.; Djurle, A. Effect of Leaf Rust on Grain Yield and Yield Traits of Durum Wheats with Race-Specific and Slow-Rusting Resistance to Leaf Rust. Plant Dis. 2006, 90, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Sugár, E.; Berzsenyi, Z.; Árendás, T.; Bónis, P. Effect of Nitrogen Fertilization and Genotype on the Yield and Yield Components of Winter Wheat. Bodenkult. J. Land Manag. Food Environ. 2016, 67, 25–34. [Google Scholar] [CrossRef]
- Harasim, E.; Wesołowski, M.; Kwiatkowski, C.; Harasim, P.; Staniak, M.; Feledyn-Szewczyk, B. The Contribution of Yield Components in Determining the Productivity of Winter Wheat (Triticum aestivum L.). Acta Agrobot. 2016, 69, 1675. [Google Scholar] [CrossRef]
- Neugschwandtner, R.; Kaul, H.-P. Sowing Ratio and N Fertilization Affect Yield and Yield Components of Oat and Pea in Intercrops. Field Crops Res. 2014, 155, 159–163. [Google Scholar] [CrossRef]
- Protić, R.; Jovin, P.; Protić, N.; Jovanovic, Ž. Mass of 1,000 Grains in Several Winter Wheat Genotypes, at Different Dates of Sowing and Rates of Nitrogen Fertilizer. Rom. Agric. Res. 2007, 24, 39–42. [Google Scholar]
- Xu, K.; Chai, Q.; Hu, F.; Fan, Z.; Yin, W. N-Fertilizer Postponing Application Improves Dry Matter Translocation and Increases System Productivity of Wheat/Maize Intercropping. Sci. Rep. 2021, 11, 22825. [Google Scholar] [CrossRef]
- Serrago, R.A.; Carretero, R.; Bancal, M.O.; Miralles, D.J. Foliar Diseases Affect the Eco-Physiological Attributes Linked with Yield and Biomass in Wheat (Triticum aestivum L.). Eur. J. Agron. 2009, 31, 195–203. [Google Scholar] [CrossRef]
- Carretero, R.; Bancal, M.O.; Miralles, D.J. Effect of Leaf Rust (Puccinia triticina) on Photosynthesis and Related Processes of Leaves in Wheat Crops Grown at Two Contrasting Sites and with Different Nitrogen Levels. Eur. J. Agron. 2011, 35, 237–246. [Google Scholar] [CrossRef]
- Simón, M.R.; Fleitas, M.C.; Castro, A.C.; Schierenbeck, M. How Foliar Fungal Diseases Affect Nitrogen Dynamics, Milling, and End-Use Quality of Wheat. Front. Plant Sci. 2020, 11, 569401. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R. Nitrogen and Photosynthesis in the Flag Leaf of Wheat (Triticum aestivum L.). Plant Physiol. 1983, 72, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.; Munk, L. Nitrogen-Induced Changes in Colony Density and Spore Production of Erysiphe Graminis f.Sp. Hordei on Seedlings of Six Spring Barley Cultivars. Plant Pathol. 1997, 46, 191–202. [Google Scholar] [CrossRef]
- Hoffland, E.; Jeger, M.J.; van Beusichem, M.L. Effect of Nitrogen Supply Rate on Disease Resistance in Tomato Depends on the Pathogen. Plant Soil 2000, 218, 239–247. [Google Scholar] [CrossRef]
- Fleitas, M.C.; Schierenbeck, M.; Gerard, G.S.; Dietz, J.I.; Golik, S.I.; Campos, P.E.; Simón, M.R. How Leaf Rust Disease and Its Control with Fungicides Affect Dough Properties, Gluten Quality and Loaf Volume under Different N Rates in Wheat. J. Cereal Sci. 2018, 80, 119–127. [Google Scholar] [CrossRef]
- Neumann, S.; Paveley, N.D.; Beed, F.D.; Sylvester-Bradley, R. Nitrogen per Unit Leaf Area Affects the Upper Asymptote of Puccinia striiformis f.Sp. Tritici Epidemics in Winter Wheat. Plant Pathol. 2004, 53, 725–732. [Google Scholar] [CrossRef]
- Devadas, R.; Simpfendorfer, S.; Backhouse, D.; Lamb, D.W. Effect of Stripe Rust on the Yield Response of Wheat to Nitrogen. Crop J. 2014, 2, 201–206. [Google Scholar] [CrossRef]
- Gao, X.; Wu, M.; Xu, R.; Wang, X.; Pan, R.; Kim, H.-J.; Liao, H. Root Interactions in a Maize/Soybean Intercropping System Control Soybean Soil-Borne Disease, Red Crown Rot. PLoS ONE 2014, 9, e95031. [Google Scholar] [CrossRef]
- Solomon, P.S.; Tan, K.-C.; Oliver, R.P. The Nutrient Supply of Pathogenic Fungi; a Fertile Field for Study. Mol. Plant Pathol. 2003, 4, 203–210. [Google Scholar] [CrossRef]
- Tavernier, V.; Cadiou, S.; Pageau, K.; Laugé, R.; Reisdorf-Cren, M.; Langin, T.; Masclaux-Daubresse, C. The Plant Nitrogen Mobilization Promoted by Colletotrichum lindemuthianum in Phaseolus Leaves Depends on Fungus Pathogenicity. J. Exp. Bot. 2007, 58, 3351–3360. [Google Scholar] [CrossRef]
- Lithourgidis, A.; Dordas, C.; Damalas, C.; Vlachostergios, D. Annual Intercrops: An Alternative Pathway for Sustainable Agriculture. Aust. J. Crop Sci. 2011, 5, 396–410. [Google Scholar]
- Jensen, E.S. Barley Uptake of N Deposited in the Rhizosphere of Associated Field Pea. Soil Biol. Biochem. 1996, 28, 159–168. [Google Scholar] [CrossRef]
- Xiao, Y.; Li, L.; Zhang, F. Effect of Root Contact on Interspecific Competition and N Transfer between Wheat and Fababean Using Direct and Indirect 15N Techniques. Plant Soil 2004, 262, 45–54. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, X.; Tong, C.; Wu, Y. Effect of Root Interaction on Nodulation and Nitrogen Fixation Ability of Alfalfa in the Simulated Alfalfa/Triticale Intercropping in Pots. Sci. Rep. 2020, 10, 4269. [Google Scholar] [CrossRef]
- Jensen, E.S. Grain Yield, Symbiotic N2 Fixation and Interspecific Competition for Inorganic N in Pea-Barley Intercrops. Plant Soil 1996, 182, 25–38. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Breathnach, J.A.; Baenziger, P.S. Effect of Growth Stage on the Relationship between Tan Spot and Spot Blotch Severity and Yield in Winter Wheat. Crop Prot. 2009, 28, 696–702. [Google Scholar] [CrossRef]
- Seem, R.C. Disease Incidence and Severity Relationships. Annu. Rev. Phytopathol. 1984, 22, 133–150. [Google Scholar] [CrossRef]
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A diagrammatic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 1948, 26c, 496–500. [Google Scholar] [CrossRef]
- Willey, R.W.; Osiru, D.S.O. Studies on Mixtures of Maize and Beans (Phaseolus vulgaris) with Particular Reference to Plant Population. J. Agric. Sci. 1972, 79, 517–529. [Google Scholar] [CrossRef]
- Bursac, Z.; Gauss, C.H.; Williams, D.K.; Hosmer, D.W. Purposeful Selection of Variables in Logistic Regression. Source Code Biol. Med. 2008, 3, 17. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield t/ha | TKW g | Crude Protein% | Powdery Mildew% | Leaf Rust % | |
|---|---|---|---|---|---|
| Powdery mildew | 0.063 | - | - | - | - |
| Leaf rust | 0.147 | - | - | - | - |
| Year | 0.003 | 0.006 | <0.001 | 0.202 | 0.009 |
| Variety | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Cultivation | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Seeding time | - | - | - | - | - |
| Year × Variety | <0.001 | <0.001 | <0.001 | <0.001 | 0.004 |
| Year × Cultivation | 0.192 | - | <0.001 | 0.669 | 0.503 |
| Variety × Cultivation | 0.428 | <0.001 | <0.001 | 0.029 | <0.001 |
| Year × Variety × Cultivation | 0.033 | - | <0.001 | 0.021 | 0.001 |
| Variety 1 | Cultivation Practice | Yield t/ha | SE of Mean | Yield t/ha | SE of Mean | TKW g | SE of Mean | TKW g | SE of Mean | Crude Protein% | SE of Mean | Crude Protein % | SE of Mean | PM 2 % | SE of Mean | PM 2 % | SE of Mean | LR 3 % | SE of Mean | LR 3 % | SE of Mean |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | ||||||||||||
| Dunav | Intercropping | 3.9 | 0.37 | 6.5 | 0.16 | 34.2 | 0.46 | 34.5 | 1.08 | 10.4 | 0.36 | 6.1 | 0.14 | 7.5 | 1.44 | 8.7 | 1.25 | 0.0 | 0 | 0.0 | 0 |
| Standalone | 5.9 | 0.28 | 9.5 | 0.04 | 29.9 | 0.49 | 30.5 | 0.29 | 8.9 | 0.21 | 6.7 | 0.17 | 5.0 | 2.04 | 12.5 | 2.50 | 0.0 | 0 | 0.0 | 0 | |
| Ilina | Intercropping | 4.7 | 0.20 | 4.2 | 0.19 | 41.8 | 0.72 | 35.4 | 0.42 | 8.9 | 0.12 | 9.1 | 0.12 | 10.0 | 0 | 12.5 | 2.50 | 0.0 | 0 | 0.0 | 0 |
| Standalone | 7.6 | 0.08 | 7.4 | 0.37 | 39.4 | 1.55 | 35.5 | 0.45 | 7.4 | 0.09 | 6.9 | 0.09 | 35.0 | 2.89 | 30.0 | 4.08 | 10.0 | 4.08 | 2.5 | 2.50 | |
| Jadar | Intercropping | 3.6 | 0.18 | 4.3 | 0.36 | 28.7 | 0.96 | 28.8 | 0.73 | 8.7 | 0.16 | 7.6 | 0.19 | 3.7 | 2.39 | 5.0 | 2.04 | 0.0 | 0 | 0.0 | 0 |
| Standalone | 6.3 | 0.52 | 7.9 | 0.30 | 28.7 | 1.10 | 28.5 | 0.55 | 7.9 | 0.12 | 6.2 | 0.07 | 15.0 | 2.89 | 6.2 | 2.39 | 0.0 | 0 | 0.0 | 0 | |
| Nataša | Intercropping | 4.2 | 0.19 | 4.4 | 0.29 | 38.4 | 0.51 | 39.2 | 0.34 | 10.4 | 0.04 | 8.2 | 0.07 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 5.0 | 2.89 |
| Standalone | 6.4 | 0.23 | 7.5 | 0.10 | 40.3 | 1.43 | 38.9 | 0.69 | 9.1 | 0.04 | 7.3 | 0.06 | 0.0 | 0 | 3.7 | 2.39 | 0.0 | 0 | 12.5 | 4.79 | |
| Odisej | Intercropping | 4.6 | 0.09 | 3.2 | 0.21 | 48.5 | 0.89 | 49.6 | 0.47 | 12.1 | 0.06 | 9.6 | 0.13 | 0.0 | 0 | 6.2 | 1.25 | 8.7 | 1.25 | 6.2 | 4.73 |
| Standalone | 7.8 | 0.15 | 6.4 | 0.46 | 45.8 | 0.55 | 46.6 | 0.39 | 9.5 | 0.04 | 7.4 | 0.09 | 10.0 | 4.08 | 15 | 5.00 | 15.0 | 2.89 | 5 | 5.00 | |
| Savo | Intercropping | 5.4 | 0.21 | 5.2 | 0.21 | 32.1 | 0.97 | 29.5 | 0.76 | 9.6 | 0.07 | 6.8 | 0.14 | 0.0 | 0 | 0.0 | 0 | 8.7 | 1.25 | 11.2 | 3.15 |
| Standalone | 8.3 | 0.39 | 6.9 | 0.38 | 28.4 | 0.83 | 28.1 | 0.62 | 7.2 | 0.07 | 5.6 | 0.09 | 0.0 | 0 | 16.2 | 2.39 | 12.5 | 2.50 | 27.5 | 2.50 | |
| Mean Intercropping | 4.4 | 4.6 | 37.3 | 36.2 | 10.0 | 7.9 | 3.5 | 5.4 | 2.9 | 3.7 | |||||||||||
| Mean Standalone | 7.1 | 7.6 | 35.4 | 34.7 | 8.3 | 6.7 | 10.8 | 13.9 | 6.2 | 7.9 | |||||||||||
| The Score | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| Level of Infection | no infection | 1–10% | 11–20% | 21–30% | 31–40% | 41–50% | 51–60% | 61–70% | 71–80% | ≥81% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jevtić, R.; Župunski, V.; Grčak, M.; Živančev, D.; Knežević, D. Cereal–Pea Intercropping Reveals Variability in the Relationships among Yield, Quality Parameters, and Obligate Pathogens Infection in Wheat, Rye, Oat, and Triticale, in a Temperate Environment. Plants 2023, 12, 2067. https://doi.org/10.3390/plants12112067
Jevtić R, Župunski V, Grčak M, Živančev D, Knežević D. Cereal–Pea Intercropping Reveals Variability in the Relationships among Yield, Quality Parameters, and Obligate Pathogens Infection in Wheat, Rye, Oat, and Triticale, in a Temperate Environment. Plants. 2023; 12(11):2067. https://doi.org/10.3390/plants12112067
Chicago/Turabian StyleJevtić, Radivoje, Vesna Župunski, Milosav Grčak, Dragan Živančev, and Desimir Knežević. 2023. "Cereal–Pea Intercropping Reveals Variability in the Relationships among Yield, Quality Parameters, and Obligate Pathogens Infection in Wheat, Rye, Oat, and Triticale, in a Temperate Environment" Plants 12, no. 11: 2067. https://doi.org/10.3390/plants12112067