


Large Field Screening for Resistance to Broomrape (Orobanche crenata Forsk.) in a Global Lentil Diversity Panel (GLDP) (Lens culinaris Medik.)

Abstract
:1. Introduction
2. Results
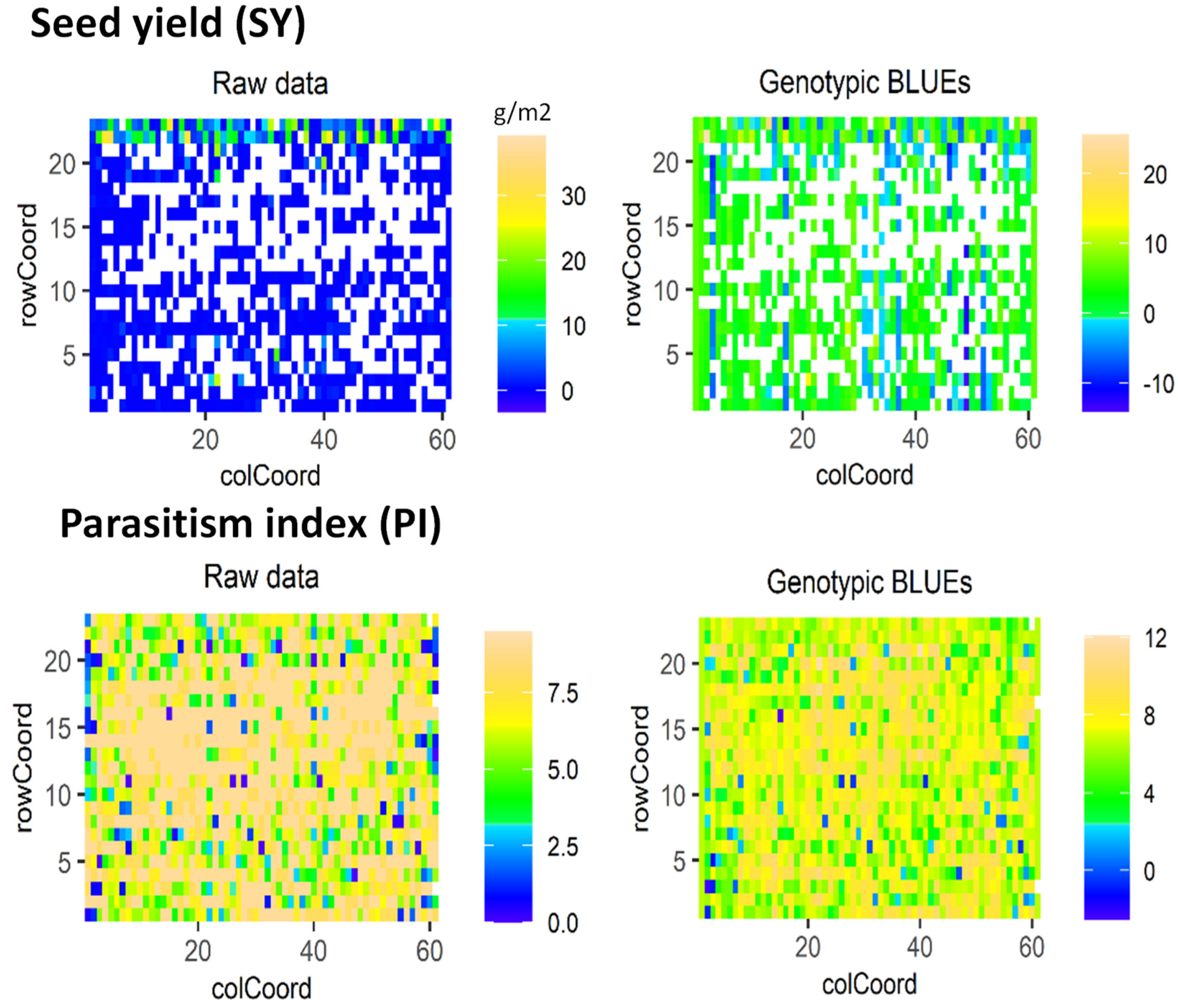
2.1. Variance, Descriptive Statistic and Spacial Analysis
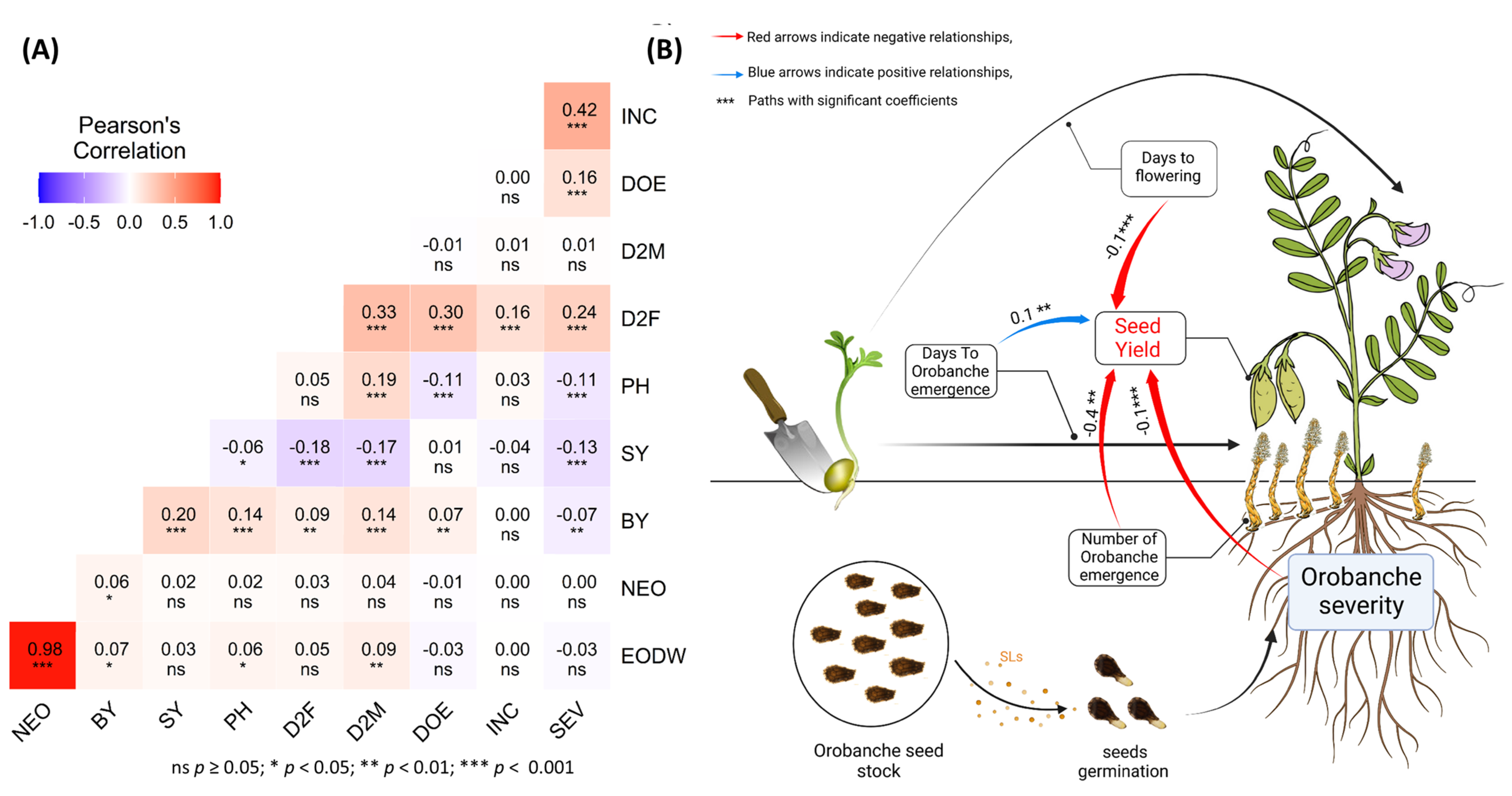
2.2. Relationship between Agronomic and Infestation Parameter
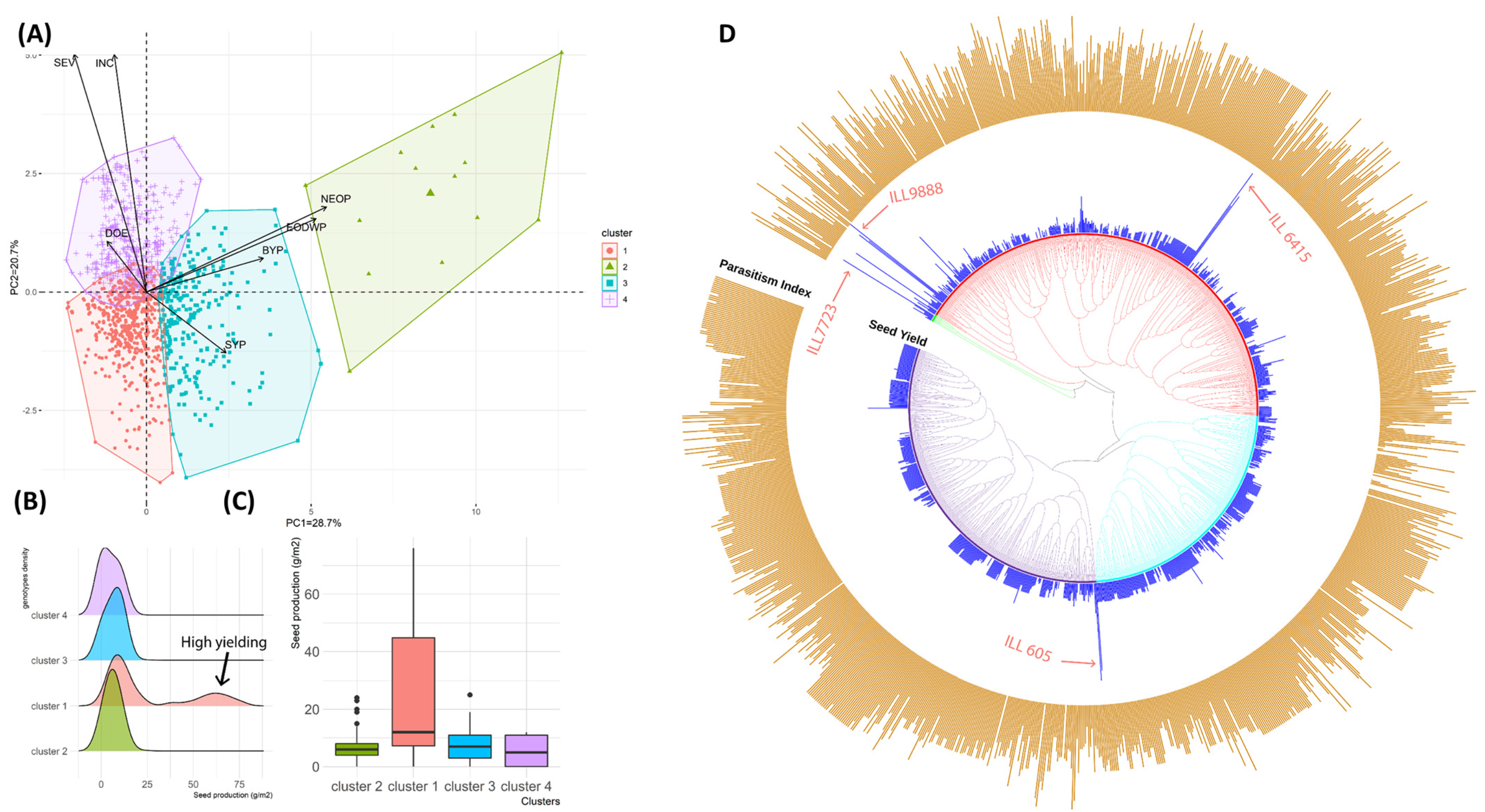
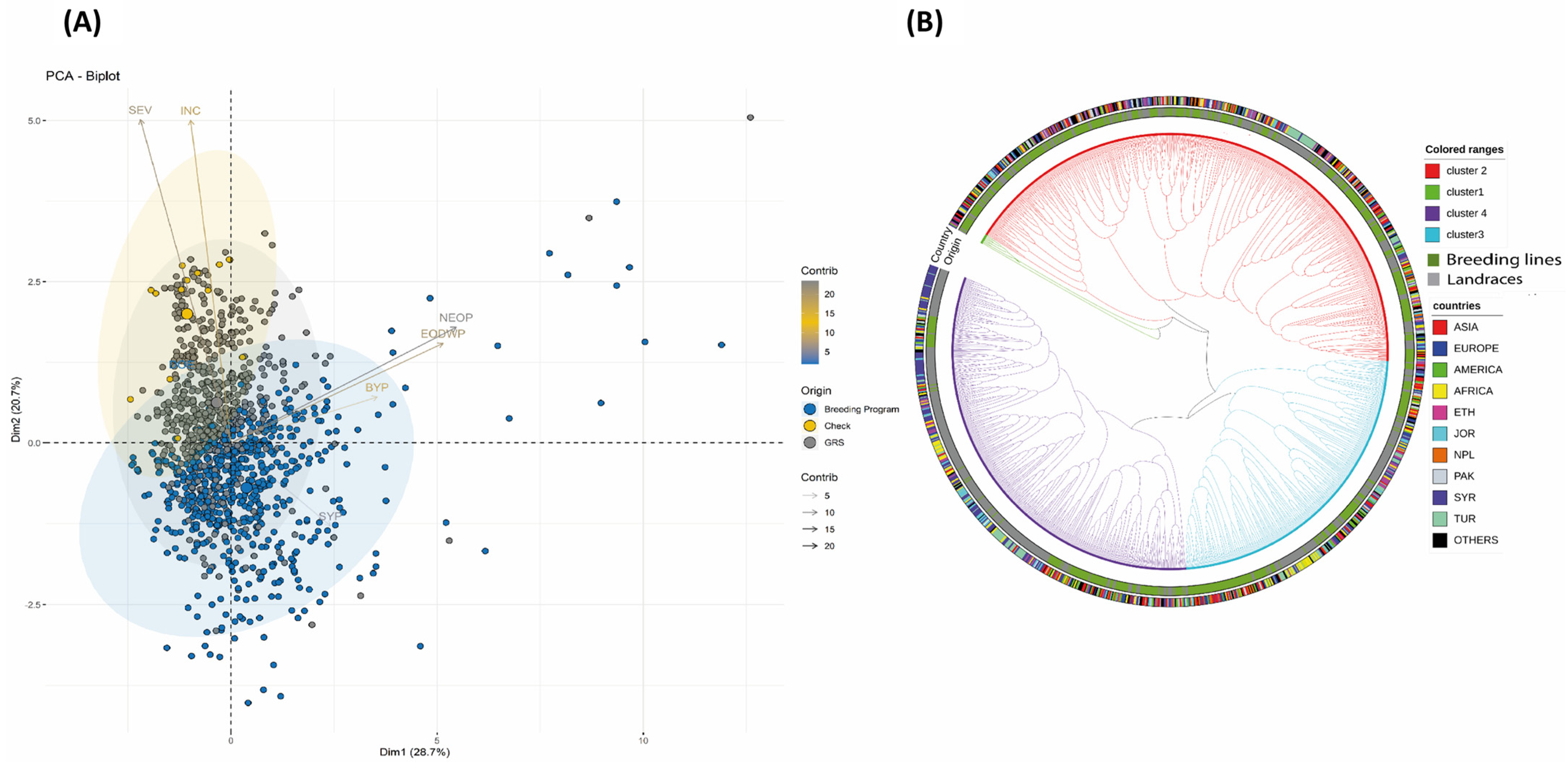
2.3. Multivariate Analysis
3. Discussion
4. Material and Methods
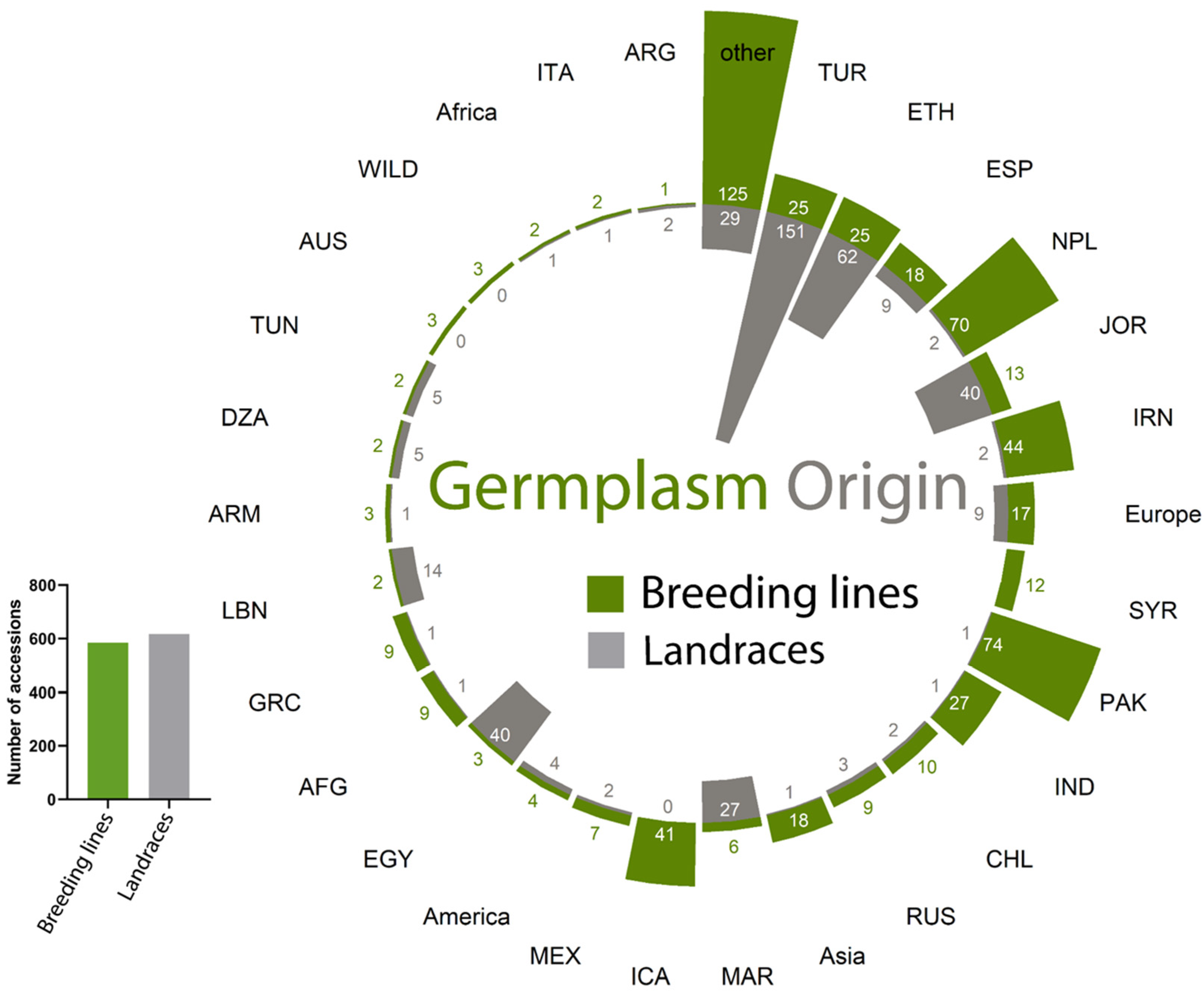
4.1. Plant Material and Field Trial
4.2. Exploratory Data Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sarker, A.; Erskine, W. Recent progress in the ancient lentil. J. Agric. Sci. 2006, 144, 19–29. [Google Scholar] [CrossRef]
- Muehlbauer, F.J.; Cho, S.; Sarker, A.; McPhee, K.E.; Coyne, C.J.; Rajesh, P.N.; Ford, R. Application of biotechnology in breeding lentil for resistance to biotic and abiotic stress. Euphytica 2006, 147, 149–165. [Google Scholar] [CrossRef]
- Amri, M.; Niane, A.A.; Agrawal, S.K.; Kemal, S.A.; Hamwieh, A.; Amri, A. Principales activités des programmes d’amélioration génétique de la lentille et du pois chiche Kabuli à ICARDA. Innov. Agron. 2019, 74, 15–24. Available online: https://hal.inrae.fr/hal-02620554%0Ahttps://hal.inrae.fr/hal-02620554/document (accessed on 4 February 2023).
- Ennami, M.; Briache, F.Z.; Gaboun, F.; Abdelwahd, R.; Ghaouti, L.; Belqadi, L.; Westwood, J.; Mentag, R. Host differentiation and variability of Orobanche crenata populations from legume species in Morocco as revealed by cross-infestation and molecular analysis. Pest Manag. Sci. 2017, 73, 1753–1763. [Google Scholar] [CrossRef]
- Idrissi, O.; Houasli, C.; Amamou, A.; Nsarellah, N. Lentil genetic improvement in Morocco: State of art of the program, major achievements and perspective. Moroc. J. Agric. Sci. 2020, 1, 9–13. [Google Scholar]
- Kumar, S.; Rajendran, K.; Kumar, J.; Hamwieh, A.; Baum, M. Current knowledge in lentil genomics and its application for crop improvement. Front. Plant Sci. 2015, 6, 78. [Google Scholar] [CrossRef]
- En-Nahli, Y.; El Arroussi, H.; Kumar, S.; Bouhlal, O.; Mentag, R.; Hejjaoui, K.; Douaik, A.; Abbes, Z.; Es-Safi, N.E.; Amri, M. Resistance to Orobanche crenata Forsk. in lentil (Lens culinaris Medik.): Exploring some potential altered physiological and biochemical defense mechanisms. J. Plant Interact. 2021, 16, 321–331. [Google Scholar] [CrossRef]
- Cartry, D.; Steinberg, C.; Gibot-Leclerc, S. Main drivers of broomrape regulation. A review. Agron. Sustain. Dev. 2021, 41, 17. [Google Scholar] [CrossRef]
- Triki, E.; Trabelsi, I.; Amri, M.; Nefzi, F.; Kharrat, M.; Abbes, Z. Effect of benzothiadiazole and salicylic acid resistance inducers on Orobanche foetida infestation in Vicia faba. Tunis. J. Plant Prot. 2018, 13, 113–125. [Google Scholar]
- Briache, F.Z.; Ennami, M.; Mbasani-Mansi, J.; Lozzi, A.; Abousalim, A.; El Rodeny, W.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Effects of Salicylic Acid and Indole Acetic Acid Exogenous Applications on Induction of Faba Bean Resistance against Orobanche crenata. Plant Pathol. J. 2020, 36, 476–490. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G., Jr. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Amri, M.; Abbes, Z.; Trabelsi, I.; Ghanem, M.E.; Mentag, R.; Kharrat, M. Chlorophyll content and fluorescence as physiological parameters for monitoring Orobanche foetida Poir. infection in faba bean. PLoS ONE 2021, 16, e0241527. [Google Scholar] [CrossRef] [PubMed]
- Abbes, Z.; Bouallegue, A.; Trabelsi, I.; Trabelsi, N.; Taamalli, A.; Amri, M.; Mhadhbi, H.; Kharrat, M. Investigation of some biochemical mechanisms involved in the resistance of faba bean (Vicia faba L.) varieties to Orobanche spp. Plant Prot. Sci. 2020, 56, 317–328. [Google Scholar] [CrossRef]
- Abbes, Z.; Trabelsi, I.; Kharrat, M.; Amri, M. Intercropping with fenugreek (Trigonella foenum-graecum) enhanced seed yield and reduced Orobanche foetida infestation in faba bean (Vicia faba). Biol. Agric. Hortic. 2019, 35, 238–247. [Google Scholar] [CrossRef]
- Abbes, Z.; Mkadmi, M.; Trabelsi, I.; Amri, M.; Kharrat, M. Orobanche foetida control in faba bean by foliar application of benzothiadiazole (BTH) and salicylic acid. Bulg. J. Agric. Sci. 2014, 20, 1439–1443. [Google Scholar]
- Attia, S.M.; El-Hady, M.M.; Saber, H.A.; Omer, M.; Khalil, S.A.; Mahmoud, S.A.; Ashrei, A.A.M.; Rehab, A.M.; Abd-Elrahman, M.A.M.; Ibrahim, Z.; et al. Misr 3, a New Orobanche Tolerant Faba Bean Variety. Egypt. J. Plant Breed. 2013, 17, 143–152. [Google Scholar] [CrossRef]
- Amri, M.; Trabelsi, I.; Abbes, Z.; Kharrat, M. Release of a new faba bean variety ‘Chourouk’ resistant to the parasitic plants Orobanche foetida and O. crenata in Tunisia. Int. J. Agric. Biol. 2019, 21, 499–505. [Google Scholar] [CrossRef]
- Kharrat, M.; Abbes, Z.; Amri, M. A New Faba Bean Small Seeded Variety Najeh Tolerant to Orobanche Registered in the Tunisian Catalogue. Tunis. J. Plant Prot. 2010, 5, 125–130. [Google Scholar]
- Briache, F.Z.; Ennami, M.; Mbasani-Mansi, J.; Gaboun, F.; Abdelwahd, R.; Fatemi, Z.E.A.; El-Rodeny, W.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Field and controlled conditions screenings of some faba bean (Vicia faba L.) genotypes for resistance to the parasitic plant Orobanche crenata Forsk. and investigation of involved resistance mechanisms. J. Plant Dis. Prot. 2019, 126, 211–224. [Google Scholar] [CrossRef]
- Mbasani-Mansi, J.; Ennami, M.; Briache, F.Z.; Gaboun, F.; Benbrahim, N.; Triqui, Z.E.A.; Mentag, R. Characterization of genetic diversity and population structure of Moroccan lentil cultivars and landraces using molecular markers. Physiol. Mol. Biol. Plants 2019, 25, 965–974. [Google Scholar] [CrossRef]
- Ennami, M.; Mbasani-Mansi, J.; Briache, F.Z.; Oussible, N.; Gaboun, F.; Ghaouti, L.; Belqadi, L.; Ghanem, M.E.; Aberkani, K.; Westwood, J.; et al. Growth-defense tradeoffs and source-sink relationship during both faba bean and lentil interactions with Orobanche crenata Forsk. Crop. Prot. 2020, 127, 104924. [Google Scholar] [CrossRef]
- Grenz, J.; Manschadi, A.; Uygur, F.; Sauerborn, J. Effects of environment and sowing date on the competition between faba bean (Vicia faba) and the parasitic weed Orobanche crenata. Field Crop. Res. 2005, 93, 300–313. [Google Scholar] [CrossRef]
- Abbes, Z.; Sellami, F.; Amri, M.; Kharrat, M. Variation in the resistance of some faba bean genotypes to Orobanche crenata. Pak. J. Bot. 2011, 43, 2017–2021. [Google Scholar]
- Trabelsi, I.; Abbes, Z.; Amri, M.; Kharrat, M. Study of some resistance mechanisms to Orobanche spp. Infestation in faba bean (Vicia faba L.) breeding lines in Tunisia. Plant Prod. Sci. 2016, 19, 562–573. [Google Scholar] [CrossRef]
- Nefzi, F.; Trabelsi, I.; Amri, M.; Triki, E.; Kharrat, M.; Abbes, Z. Response of some chickpea (Cicer arietinum L.) genotypes to Orobanche foetida Poir. parasitism. Chil. J. Agric. Res. 2016, 76, 170–178. [Google Scholar] [CrossRef]
- De Ron, A.M. Grain Legumes; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Abdallah, F.; Kumar, S.; Amri, A.; Mentag, R.; Kehel, Z.; Mejri, R.K.; Triqui, Z.E.A.; Hejjaoui, K.; Baum, M.; Amri, M. Wild Lathyrus species as a great source of resistance for introgression into cultivated grass pea (Lathyrus sativus L.) against broomrape weeds (Orobanche crenata Forsk. and Orobanche foetida Poir.). Crop. Sci. 2021, 61, 263–276. [Google Scholar] [CrossRef]
- Mesa-Garcia, J.; Garcia-Torres, L. A competition index for Orobanche crenata Forsk effects on broad bean (Vicia faba L.). Weed Res. 1984, 24, 379–382. [Google Scholar] [CrossRef]
- Linke, K.-H.; El-Moneim, A.A.; Saxena, M. Variation in resistance of some forage legumes species to Orobanche crenata Forsk. Field Crop. Res. 1993, 32, 277–285. [Google Scholar] [CrossRef]
- Trabelsi, I.; Yoneyama, K.; Abbes, Z.; Amri, M.; Xie, X.; Kisugi, T.; Kim, H.; Kharrat, M. Characterization of strigolactones produced by Orobanche foetida and Orobanche crenata resistant faba bean (Vicia faba L.) genotypes and effects of phosphorous, nitrogen, and potassium deficiencies on strigolactone production. South Afr. J. Bot. 2017, 108, 15–22. [Google Scholar] [CrossRef]
- Radwan, M.S.; Abdalla, M.M.F.; Fischbeck, G.; Metwally, A.A.; Darwish, D.S. Variation in Reaction of Faba Bean Lines to Different Accessions of Orobanche crenata Forsk. Plant Breed. 1988, 101, 208–216. [Google Scholar] [CrossRef]
- Manschadi, A.M.; Sauerborn, J.; Kroschel, J.; Saxena, M.C. Effect of plant density on grain yield, root–length density and Orobanche crenata infestation in two faba bean genotypes. Weed Res. 1997, 37, 39–49. [Google Scholar] [CrossRef]
- Trabelsi, I.; Abbes, Z.; Amri, M.; Kharrat, M. Performance of faba bean genotypes with Orobanche foetida Poir. and Orobanche crenata Forsk. infestation in Tunisia. Chil. J. Agric. Res. 2015, 75, 27–34. [Google Scholar] [CrossRef]
- Darwish, D.S.; Abdalla, M.M.F.; El-Metwally, E.A.; El-Sherbeeny, M.H.; Attia, S.M. Investigations on faba beans, Vicia faba L. 13-Performance of some faba bean genotypes and their hybrids under Orobanche-infestation. Egypt J. Plant Breed. 1999, 3, 231–246. [Google Scholar]
- Abbes, Z.; Kharrat, M.; Delavault, P.; Simier, P.; Chaïbi, W. Field evaluation of the resistance of some faba bean (Vicia faba L.) genotypes to the parasitic weed Orobanche foetida Poiret. Crop. Prot. 2007, 26, 1777–1784. [Google Scholar] [CrossRef]
- Abbes, Z.; Sellami, F.; Amri, M.; Kharrat, M. Effect of sowing date onOrobanche foetidainfection and seed yield of resistant and susceptible faba bean cultivars. Acta Phytopathol. Et Èntomol. Hung. 2010, 45, 267–275. [Google Scholar] [CrossRef]
- Revelle, W.; Revelle, M. Procedures for psychological, psychometric, and personality. R Package Bpsych 2017, 1. Available online: https://www.scholars.northwestern.edu/en/publications/psych-procedures-for-personality-and-psychological-research (accessed on 4 February 2023).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). Statistician 2017, 56, e24. [Google Scholar]
- Qasem, J.R. Broomrapes (Orobanche spp.) the Challenge and Management: A review. Jordan J. Agric. Sci. 2021, 17, 117–150. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | BY | D2F | D2M | DOE | EODW | INC | NEO | PH | PI | SEV | SY |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Block | 24,993.2 ** | 557.4 ** | 32.7 ** | 324.9 ** | 18.9 ** | 978.7 ns | 30.0 ** | 125.6 ** | 20.1 ** | 15.8 ** | 2518.1 ** |
| Genotype | 13,144.8 ** | 103.4 ** | 21.1 ** | 21.5 ns | 10.6 ** | 399.4 ns | 21.8 ** | 24.82 ** | 4.4 ns | 1.9 ns | 74.6 ns |
| Check | 29,319.6 ** | 7.9 ns | 4.9 ns | 185 ** | 5.7 ** | 6667.0 ** | 3.2 ** | 11.8 ns | 54.5 ** | 18.2 ** | 1300.1 ** |
| Gen vs. Check | 13,107.8 ** | 103.6 ** | 21.2 ** | 21.2 ns | 10.6 ** | 385.0 ns | 21.8 ** | 24.8 ** | 4.3 ns | 1.9 ns | 71.8 ns |
| Residuals | 6583.9 | 5.2 | 4.8 | 19.7 | 0.4 | 596.4 | 0.3 | 9.5 | 6.2 | 2.9 | 167.9 |
| Trait | Count | Mean | SE | Min | Max |
|---|---|---|---|---|---|
| D2F | 1315 | 85 | 0.3 | 60 | 111 |
| D2M | 1315 | 127 | 0.1 | 115 | 145 |
| DOE | 1315 | 88 | 0.2 | 70 | 109 |
| PH | 1315 | 28.8 | 0.2 | 14 | 46.3 |
| NEO | 1315 | 1.2 | 0.1 | 0 | 168 |
| EODW | 1315 | 1.13 | 0.1 | 0 | 114.3 |
| INC | 1315 | 91.4 | 0.7 | 0 | 138.1 |
| SEV | 1315 | 8 | 0.1 | 2 | 9 |
| PI | 1315 | 7 | 0.1 | 0 | 9 |
| BY | 1315 | 211.5 | 3.3 | 0 | 1797.9 |
| SY | 1315 | 7.1 | 0.2 | 0 | 108.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
En-nahli, Y.; Hejjaoui, K.; Mentag, R.; Es-safi, N.E.; Amri, M. Large Field Screening for Resistance to Broomrape (Orobanche crenata Forsk.) in a Global Lentil Diversity Panel (GLDP) (Lens culinaris Medik.). Plants 2023, 12, 2064. https://doi.org/10.3390/plants12102064
En-nahli Y, Hejjaoui K, Mentag R, Es-safi NE, Amri M. Large Field Screening for Resistance to Broomrape (Orobanche crenata Forsk.) in a Global Lentil Diversity Panel (GLDP) (Lens culinaris Medik.). Plants. 2023; 12(10):2064. https://doi.org/10.3390/plants12102064
Chicago/Turabian StyleEn-nahli, Youness, Kamal Hejjaoui, Rachid Mentag, Nour Eddine Es-safi, and Moez Amri. 2023. "Large Field Screening for Resistance to Broomrape (Orobanche crenata Forsk.) in a Global Lentil Diversity Panel (GLDP) (Lens culinaris Medik.)" Plants 12, no. 10: 2064. https://doi.org/10.3390/plants12102064