
Invasive Mechanisms of One of the World’s Worst Alien Plant Species Mimosa pigra and Its Management

Abstract
:1. Introduction
1.1. Invasiveness of the Species
1.2. Impact of the Species
2. Invasive Mechanism
2.1. Reproduction and Growth
2.2. Adaptivity and Plasticity
2.3. Natural Enemy
2.4. Mutualism
2.5. Allelopathy
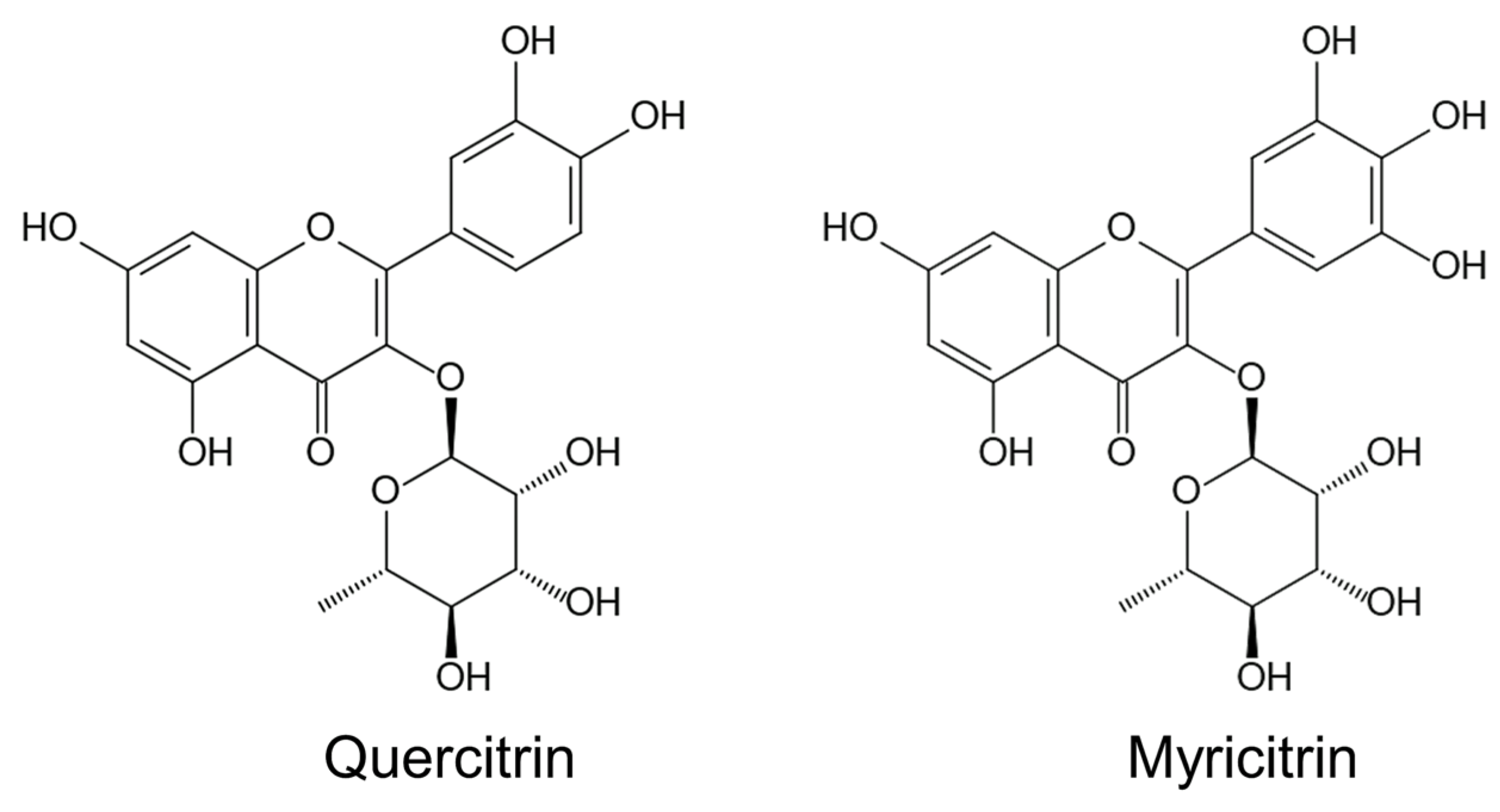
2.6. Secondary Metabolites

{kind=link}
{kind=link}
{kind=link}
| Characteristic | Reference |
|---|---|
| Rapid growth through the vegetative phase to flowering | [4,5,16,30,31] |
| Self-compatible | [4] |
| High seed output | [12,16,31,32,33,34] |
| High germination rate | [5,16,34] |
| Longevity of seeds | [31,36] |
| Regeneration from remaining plant parts | [34,41] |
| Vigorous mutualism with rhizobia and arbuscular mycorrhizal fungi | [62,63,64,70,71] |
| Colonizing with rhizobia with high nitrogen-fixing activity in the invasive ranges | [58,66,67,68,69] |
| Very few natural enemies in the invasive ranges | [53] |
| Allelopathy | [92,93,94,95,96,97] |
| Secondary metabolites | [104,105,106,107,108,109,110] |
3. Management
3.1. Mechanical Control and Fire
3.2. Chemical Control
3.3. Biological Control
| Species | Order | Group | Year of the First Release | Attacked Plant Part | Reference |
|---|---|---|---|---|---|
| Carmenta mimosa | Lepidoptera | Moth | 1989 | Stem | [50,126,128] |
| Neurostrota gunniella | Lepidoptera | Moth | 1989 | Stem, pinnules, seedling | [52,126,129,130,131,132] |
| Macaria pallidata | Lepidoptera | Moth | 2002 | Leaf | [133] |
| Coelocephalapion pigrae | Coleoptera | Weevil | 1994 | Flower, leaf | [126,134] |
| Chalcodermus serripes | Coleoptera | Weevil | 1996 | Seed, leaf | [126,137] |
| Acanthoscelides puniceus | Coleoptera | Bean weevil | 1983 | Seed | [126,129] |
| Chlamisus mimosae | Coleoptera | Leaf beetle | 1985 | Stem, leaf | [126] |
| Malacorhinus irregularis | Coleoptera | Leaf beetle | 2000 | Leaf, root, nodule | [126,135] |
| Diabole cubensis | Pucciniales | Fungus | 1996 | Leaf | [136] |
3.4. Integrated Control
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Global Invasive Species Database. Species Profile: Mimosa pigra. Available online: http://www.iucngisd.org/gisd/speciesname/Mimosa+pigra (accessed on 4 April 2023).
- Invasive Species Compendium, Mimosa pigra. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.34199 (accessed on 4 April 2023).
- IUCN Red List of Threatened Species, Mimosa pigra. Available online: https://www.iucnredlist.org/species/49486238/49486516 (accessed on 4 April 2023).
- Beilfuss, R. Adaptive Management of the Invasive Shrub Mimosa pigra at Gorongosa National Park; Department of Scientific Services: Gorongoza, Mozanbique, 2007; pp. 1–19.
- Lonsdale, W.M.; Miller, I.L.; Forno, I.W. The biology of Australian weeds. 20. Mimosa pigra. Plant Prod. Q. 1989, 4, 119–131. [Google Scholar]
- Miller, I.L.; Lonsdale, W.M. Early records of Mimosa pigra in the Northern Territory (woody weed). Plant Prod. Q. 1987, 2, 140–142. [Google Scholar]
- Witt, A.; Luke, Q. Guide to the Naturalized and Invasive Plants of Eastern Africa; CABI: Wallingford, UK, 2017; pp. 1–601. [Google Scholar]
- Plants of the World. Royal Botanical Gardens-Kew, Mimosa pigra L. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:313003-2 (accessed on 4 April 2023).
- Miller, I.L.; Wilson, C.G. Weed threats to Northern Territory wetlands. In Wetland Research in the Wet-Dry Tropics of Australia; Finlayson, C.M., Ed.; Supervising Scientist: Darwin, Australia, 1995; pp. 190–195. [Google Scholar]
- Crop Protection Compendium, Mimosa pigra L. Available online: http://www.mikepalmer.co.uk/woodyplantecology/docs/CPC-Mimosa_pigra.pdf (accessed on 4 April 2023).
- Marko, M. Controlling invasion of the exotic shrub (Mimosa pigra) in tropical Australian wetlands. Restor. Reclam. Rev. 1999, 4, 1–10. [Google Scholar]
- Lonsdale, W.M. Rates of spread of an invading species—Mimosa pigra in northern Australia. J. Ecol. 1993, 81, 513–521. [Google Scholar] [CrossRef]
- Mumba, M.; Thompson, J.R. Hydrological and ecological impacts of dams on the Kafue Flats floodplain system, southern Zambia. Phys. Chem. Earth. Part A/B/C 2005, 30, 442–447. [Google Scholar] [CrossRef]
- Shanungu, G.K. Management of the invasive Mimosa pigra L. in Lochinvar National Park, Zambia. Biodiversity 2009, 10, 56–60. [Google Scholar] [CrossRef]
- Witt, A.B.; Floyd, K.S.; Nunda, W.; Beale, T.; Shanungu, G.; Kriticos, D.J. Mimosa pigra in eastern and southern Africa: Distribution and socio-ecological impacts. Austral Ecol. 2020, 45, 788–799. [Google Scholar] [CrossRef]
- Marambe, B.; Amarasinghe, L.; Silva, K.; Gamage, G.; Dissanayake, S.; Seneviratne, A. Distribution, biology and management of Mimosa pigra in Sri Lanka. In Research and Management of Mimosa pigra; Julien, M., Flanagan, G., Heard, T., Hennecke, B., Paynter, Q., Wilson, C., Eds.; CSIRO Entomology: Canberra, Australia, 2004; pp. 85–90. [Google Scholar]
- Pitt, J.L.; Miller, I.L. A review of survey techniques for the detection of weeds with particular reference to Mimosa pigra L. Australia and Thailand. Plant Prod. Q. 1988, 3, 149–155. [Google Scholar]
- Rijal, S.; Cochard, R. Invasion of Mimosa pigra on the cultivated Mekong River floodplains near Kratie, Cambodia: Farmers’ coping strategies, perceptions, and outlooks. Reg. Environ. Chang. 2016, 16, 681–693. [Google Scholar] [CrossRef]
- Pramual, P.; Khumkratok, S.; Wongpakam, K. Population genetics of invasive weed Mimosa pigra L. (Mimosoideae) in Thailand. Pak. J. Bot. 2011, 43, 2721–2726. [Google Scholar]
- Son, N.H.; Lam, P.V.; Cam, N.V.; Thanh, D.V.T.; Dung, N.V.; Khanh, L.D.; Forno, I.W. Preliminary studies on control of Mimosa pigra in Vietnam. In Research and Management of Mimosa pigra; Julien, M., Flanagan, G., Heard, T., Hennecke, B., Paynter, Q., Wilson, C., Eds.; CSIRO Entomology: Canberra, Australia, 2004; pp. 110–116. [Google Scholar]
- Mansor, A.; Crawley, M.J. Current status of Mimosa pigra L. infestation in Peninsular Malaysia. Trop. Life Sci. Res. 2011, 22, 37–49. [Google Scholar] [PubMed]
- Braithwaite, R.W.; Lonsdale, W.M.; Estbergs, J.A. Alien vegetation and native biota in tropical Australia: The impact of Mimosa pigra. Biol. Conserv. 1989, 48, 189–210. [Google Scholar] [CrossRef]
- Welgama, A.; Florentine, S.; Roberts, J.A. Global review of the woody invasive alien species Mimosa pigra (giant sensitive plant): Its biology and management implications. Plants 2022, 11, 2366. [Google Scholar] [CrossRef] [PubMed]
- Heard, T.A.; Paynter, Q. Mimosa pigra (Leguminosae). In Biological Control of Tropical Weeds Using Arthropods; Muniappan, R., Reddy, G.V.P., Raman, A., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 256–273. [Google Scholar]
- Walden, D.; van Dam, R.; Finlayson, C.; Storrs, M.; Lowry, J.; Kriticos, D. A risk assessment of the tropical wetland weed Mimosa pigra in northern Australia. In Research and Management of Mimosa pigra; Julien, M., Flanagan, G., Heard, T., Hennecke, B., Paynter, Q., Wilson, C., Eds.; CSIRO Entomology: Canberra, Australia, 2004; pp. 11–21. [Google Scholar]
- Haq, R.U.; Eiam-Ampai, K.; Ngoprasert, D.; Sasaki, N.; Shrestha, R.P. Changing landscapes and declining populations of resident waterbirds: A 12-year study in Bung Boraphet Wetland, Thailand. Tropic. Conserv. Sci. 2018, 11, 1–17. [Google Scholar] [CrossRef]
- Thamasara, S. Mimosa pigra management in irrigation systems in Thailand. In Proceedings of the International Symposium on Mimosa pigra Management, Chiang Mai, Thailand, 22–26 February 1982; International Plant Protection Center: Chiang Mai, Thailand, 1983; pp. 109–112. [Google Scholar]
- Chamroeun, K.; Seang, P.T.; Sophal, H.; Hout, S.S.; Vuthy, H. An investigation of the impacts of Mimosa pigra on rice and fishery productivity in Kandal Province, Cambodia. In Economy and Environment: Case Studies in Cambodia, Singapore; McKenney, B., Ed.; Economy and Environment Program for Southeast Asia: Singapore, 2002; pp. 24–31. [Google Scholar]
- Mohamed, M.S.; Seman, I.A. Occurrence of common weeds in immature plantings of oil palm plantations in Malaysia. Planter 2012, 88, 537–547. [Google Scholar]
- Vitelli, J.S.; Madigan, B.A.; Worsley, K.J. Mimosa pigra in Queensland. In Proceedings of the 15th Australian Weeds Conference, Adelaide, Australia, 24–28 September 2006; Weed Management Society of South Australia: Adelaide, Australia, 2006; pp. 251–254. [Google Scholar]
- Lonsdale, W.M.; Miller, I.L.; Forno, I.W. Mimosa pigra L. In The Biology of Australian Weeds; Groves, R.H., Shepherd, R.C.H., Richardson, R.G., Eds.; RG and FJ Richardson Publisher: Melbourne, Australia, 1995; pp. 169–188. [Google Scholar]
- Lonsdale, W.M. The biology of Mimosa pigra. In A Guide to the Management of Mimosa pigra; Harley, K.L.S., Ed.; CSIRO: Canberra, Australia, 1992; pp. 8–32. [Google Scholar]
- Lonsdale, W.; Braithwaite, R.W. The shrub that conquered the bush. New Sci. 1988, 15, 52–55. [Google Scholar]
- Creager, R.A. Seed germination, physical and chemical control of catclaw mimosa (Mimosa pigra var. pigra). Weed Technol. 1992, 6, 884–891. [Google Scholar] [CrossRef]
- van Klinken, R.D.; Goulier, J.B. Habitat-specific seed dormancy-release mechanisms in four legume species. Seed Sci. Res. 2013, 23, 181–188. [Google Scholar] [CrossRef]
- Lukitsch, B.; Elliott, L. Mimosa pigra seed bank remains significant 10 years after stand removal: Further investigation on a floodplain in northern Australia. In Proceedings of the 18th Australasian Weeds Conference, Melbourne, Australia, 8–11 October 2012; Weed Society of Victoria Inc.: Melbourne, Australia, 2012; pp. 364–366. [Google Scholar]
- Marambe, B.; Amarasinghe, L.; Dissanayake, S. Growth and development of Mimosa pigra: An alien invasive plant in Sri Lanka. In Plant Invasions: Species Ecology and Ecosystem Manageme; Brundu, G., Brock, J., Camarda, I., Child, L., Wade, M., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2001; pp. 115–122. [Google Scholar]
- Facts Sheet: Restricted Invasive Plant: Mimosa pigra: Queensland Department of Agriculture and Fisheries. Available online: https://www.daf.qld.gov.au/__data/assets/pdf_file/0007/65149/mimosa-pigra.pdf (accessed on 4 April 2023).
- Presnell, K. The potential use of mimosa as fuel for power generation. In Proceedings of the 3rd International Symposium on the Management of of Mimosa pigra, Darwin, Australia, 22–25 September 2002; CSIRO Entomology: Canberra, Australia, 2004; pp. 68–72. [Google Scholar]
- Paynter, Q.; Flanagan, G.J. Integrating herbicide and mechanical control treatments with fire and biological control to manage an invasive wetland shrub, Mimosa pigra. J. Appl. Ecol. 2004, 41, 615–629. [Google Scholar] [CrossRef]
- Lonsdale, W.M.; Miller, I.L. Fire as a management tool for a tropical woody weed: Mimosa pigra in northern Australia. J. Environ. Manag. 1993, 30, 77–87. [Google Scholar] [CrossRef]
- Thompson, J.D.; McNeilly, T.; Gray, A.J. Population variation in Spartina anglica C.E. Hubbard. I. Evidence from a common garden experiment. New Phytol. 1991, 117, 115–128. [Google Scholar] [CrossRef]
- Mack, R.M. Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biol. Conserv. 1996, 78, 107–121. [Google Scholar] [CrossRef]
- Chengxu, W.; Mingxing, Z.; Xuhui, C.; Bo, Q. Review on allelopathy of exotic invasive plants. Procedia. Engin. 2011, 18, 240–246. [Google Scholar] [CrossRef]
- Warren, R.J.; Matt Candeias, M.; Labatore, A.; Olejniczak, M.; Yang, L. Multiple mechanisms in woodland plant species invasion. J. Plant Ecol. 2019, 12, 201–209. [Google Scholar] [CrossRef]
- NurZhafarina, A.; Asyraf, M. Effects of biotic and abiotic environmental stimuli on the morphology and biomass allocation of Mimosa pigra L. Sains Malays 2017, 46, 1241–1248. [Google Scholar]
- Harley, K.L.S.; Gillett, J.D.; Winder, J.; Forno, I.W.; Segura, R.; Miranda, H.; Kassulke, R.C. Natural enemies of Mimosa pigra and M. berlandieri (Mimosaceae) and prospects for biological control of M. pigra. Environ. Entomol. 1995, 24, 1664–1678. [Google Scholar] [CrossRef]
- Heard, T.A.; Pettit, W. Review and analysis of the surveys for natural enemies of Mimosa pigra: What does it tell us about surveys for broadly distributed hosts? Biol. Control. 2005, 34, 247–254. [Google Scholar] [CrossRef]
- Heard, T.A.; Segura, R. Agents for biological control of Mimosa pigra in Australia: Review and future prospects. In Research and Management of Mimosa pigra; Julien, M., Flanagan, G., Heard, T., Hennecke, B., Paynter, Q., Wilson, C., Eds.; CSIRO Entomology: Canberra, Australia, 2004; pp. 126–140. [Google Scholar]
- Paynter, Q. Evaluating Mimosa pigra biological control in Australia. In Research and Management of Mimosa pigra; Julien, M., Flanagan, G., Heard, T., Hennecke, B., Paynter, Q., Wilson, C., Eds.; CSIRO Entomology: Canberra, Australia, 2004; pp. 141–148. [Google Scholar]
- Evans, H.C.; Carrión, G.; Guzman, G. A new species of Sphaerulina and its Phloeospora anamorph, with potential for biological control of Mimosa pigra. Mycol. Res. 1993, 97, 59–67. [Google Scholar] [CrossRef]
- Evans, H.C.; Carrión, G.; Ruiz-Belin, F. Mycobiota of the giant sensitive plant, Mimosa pigra sensu lato in the Neotropics. Mycol. Res. 1995, 99, 420–428. [Google Scholar] [CrossRef]
- Flanagan, G.J.; Wilson, C.G.; Gillett, J.D. The abundance of native insects on the introduced weed Mimosa pigra in Northern Australia. J. Trop. Ecol. 1990, 6, 219–230. [Google Scholar] [CrossRef]
- Moulin, L.; Munive, A.; Dreyfus, B.; Boivin-Masson, C. Nodulation of legumes by members of the b-subclass of proteobacteria. Nature 2001, 411, 948–950. [Google Scholar] [CrossRef]
- Vandamme, P.; Goris, J.; Chen, W.M.; de Vos, P.; Willems, A. Burkholderia tuberum sp. nov. and Burkholderia phymatum sp. nov. nodulate the roots of tropical legumes. Syst. Appl. Microbiol. 2002, 25, 507–512. [Google Scholar] [CrossRef]
- Barrett, C.F.; Parker, M.A. Coexistence of Burkholderia, Cupriavidus and Rhizobium sp. nodule bacteria on two Mimosa species in Costa Rica. Appl. Environ. Microbiol. 2006, 72, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; James, E.K.; Coenye, T.; Chou, J.H.; Barrios, E.; de Faria, S.M.; Ellio, N.; Sheu, S.Y.; Sprent, J.I.; Vandamme, P. Burkholderia mimosarum sp. nov., isolated from root nodules of Mimosa spp. from Taiwan and South America. Int. J. Syst. Evol. Microbiol. 2006, 56, 1847–1851. [Google Scholar] [CrossRef]
- Elliott, G.N.; Chou, J.H.; Chen, W.M.; Bloemberg, G.V.; Bontemps, C.; Martínez-Romero, E.; Velázquez, E.; Young, J.P.W.; Sprent, J.I.; James, E.K. Burkholderia spp. are the most competitive symbionts of Mimosa, particularly under N-limited conditions. Environ. Microbiol. 2009, 11, 762–778. [Google Scholar]
- Chen, W.M.; James, E.K.; Chou, J.H.; Sheu, S.Y.; Yang, S.Z.; Sprent, J.I. β-Rhizobia from Mimosa pigra, a newly discovered invasive plant in Taiwan. New Pytol. 2005, 168, 661–675. [Google Scholar] [CrossRef] [PubMed]
- Tsyganova, A.V.; Brewin, N.J.; Tsyganov, V.E. Structure and development of the legume-rhizobial symbiotic interface in infection threads. Cells 2021, 10, 1050. [Google Scholar] [CrossRef] [PubMed]
- Mathesius, U. Are legumes different? Origins and consequences of evolving nitrogen fixing symbioses. J. Plant Physiol. 2022, 276, 153765. [Google Scholar] [CrossRef]
- de Faria, S.M.; de Lima, H.C. Additional studies of the nodulation status of legume species in Brazil. Plant Soil 1998, 200, 185–192. [Google Scholar] [CrossRef]
- Saur, E.; Carcelle, S.; Guezennec, S.; Rousteau, A. Nodulation of legume species in wetlands of Guadeloupe (Lesser Antilles). Wetlands 2000, 20, 730–734. [Google Scholar] [CrossRef]
- James, E.K.; de Fatima Loureiro, M.; Pott, A.; Pott, V.J.; Martins, C.M.; Franco, A.A.; Sprent, J.I. Flooding-tolerant legume symbioses from the Brazilian Pantanal. New Phytol. 2001, 150, 723–738. [Google Scholar] [CrossRef]
- Chen, W.M.; de Faria, S.M.; Straliotto, R.; Pitard, R.M.; Simões-Araùjo, J.L.; Chou, J.H.; Chou, Y.J.; Barrios, E.; Prescott, A.R.; Elliott, G.N.; et al. Proof that Burkholderia forms effective symbioses with legumes: A study of novel Mimosa-nodulating strains from South America. Appl. Environ. Microbiol. 2005, 71, 7461–7471. [Google Scholar] [CrossRef]
- Willems, A.; Tian, R.; Bräu, L.; Goodwin, L.; Han, J.; Liolios, K.; Huntemann, M.; Pati, A.; Woyke, T.; Mavrommatis, K.; et al. Genome sequence of Burkholderia mimosarum strain LMG 23256T, a Mimosa pigra microsymbiont from Anso, Taiwan. Stand. Genom. Sci. 2014, 9, 484–494. [Google Scholar] [CrossRef]
- Howieson, J.G.; Ewing, M.A.; D’antuono, M.F. Selection for acid tolerance in Rhizobium meliloti. Plant Soil. 1988, 105, 179–188. [Google Scholar] [CrossRef]
- Terpolilli, J.J. Why Are the Symbioses between Some Genotypes of Sinorhizobium and Medicago suboptimal for N2 Fixation? Murdoch University: Perth, Australia, 2009; pp. 1–223. Available online: https://researchrepository.murdoch.edu.au/id/eprint/683/ (accessed on 4 April 2023).
- Parker, M.A.; Wurtz, A.K.; Paynter, Q. Nodule symbiosis of invasive Mimosa pigra in Australia and in ancestral habitats: A comparative analysis. Biol. Invasions 2007, 9, 127–138. [Google Scholar] [CrossRef]
- Rodríguez, Y.; Dalpé, Y.; Séguin, S.; Fernández, K.; Fernández, F.; Rivera, R.A. Glomus cubense sp. nov., an arbuscular mycorrhizal fungus from Cuba. Mycotaxon 2011, 118, 337–347. [Google Scholar] [CrossRef]
- Santillán-Manjarrez, J.; Solís-Hernández, A.P.; Castilla-Hernández, P.; Maldonado-Mendoza, I.E.; Vela-Correa, G.; Chimal-Hernández, A.; Hernández-Díaz, C.; Signoret-Poillon, M.; Diederik van Tuinen, D.; Rivera-Becerril, F. Exploring plant root-fungal interactions in a neotropical freshwater wetland. Bot. Sci. 2019, 97, 661–674. [Google Scholar] [CrossRef]
- Davison, J.; Öpik, M.; Daniell, T.J.; Moora, M.; Zobel, M. Arbuscular mycorrhizal fungal communities in plant roots are not random assemblages. FEMS Microbiol. Ecol. 2011, 78, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; pp. 1–815. [Google Scholar]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Tang, H.; Hassan, M.U.; Feng, L.; Nawaz, M.; Shah, A.N.; Qari, S.H.; Liu, Y.; Miao, J. The critical role of arbuscular mycorrhizal fungi to improve drought tolerance and nitrogen use efficiency in crops. Front. Plant Sci. 2022, 13, 919166. [Google Scholar] [CrossRef]
- Ramírez-Viga, T.K.; Aguilar, R.; Castillo-Argüero, S.; Chiappa-Carrara, X.L.; Guadarrama, P.; Ramos-Zapata, J. Wetland plant species improve performance when inoculated with arbuscular mycorrhizal fungi: A meta-analysis of experimental pot studies. Mycorrhiza 2018, 28, 477–493. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 419–426. [Google Scholar] [CrossRef]
- Cappuccino, N.; Arnason, J.T. Novel chemistry of invasive exotic plants. Biol. Lett. 2006, 2, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Meiners, S.J.; Kong, C.H.; Ladwig, L.M.; Pisula, N.L.; Lang, K.A. Developing an ecological context for allelopathy. Plant. Ecol. 2012, 213, 1861–1867. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; pp. 1–422. [Google Scholar]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest. Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Involvement of allelopathy in the invasive potential of Tithonia diversifolia. Plants 2020, 9, 766. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy of Lantana camara as an invasive plant. Plants 2021, 10, 1028. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy of knotweeds as invasive plants. Plants 2022, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy and allelochemicals of Leucaena leucocephala as an invasive plant species. Plants 2022, 11, 1672. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy and allelochemicals of Imperata cylindrica as an invasive plant species. Plants 2022, 11, 2551. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Kato, M. Allelopathy and allelochemicals of Solidago canadensis L. and S. altissima L. for their naturalization. Plants 2022, 11, 3235. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kato, M. Evolution of the secondary metabolites in invasive plant species Chromolaena odorata for the defense and allelopathic functions. Plants 2023, 12, 521. [Google Scholar] [CrossRef] [PubMed]
- Koodkaew, I.; Rottasa, R. Allelopathic effects of giant sensitive plant (Mimosa pigra) leaf powder on germination and growth of popping pod and purslane. Int. J. Agric. Biol. 2017, 19, 1113–1118. [Google Scholar] [CrossRef]
- Oso, O.A.; Ajayi, B.; Okanume, O.E. Descriptive anatomy of invasive weed, Ruellia tuberosa Linn. Asian J. Res. Bot. 2022, 7, 31–36. [Google Scholar]
- Prasangani, W.D.; Sinhalage, I.D.; Karunathilake, A.A.; Madusinghe, M. Phytotoxicity Effect of three invasive plant species: Mimosa pigra, Parthenium hysierophorus, Ulex europaeus. In Proceedings of the Research Symposium of Uva Wellassa University, Badulla, Sri Lanka, 22–23 November 2012. [Google Scholar]
- Araújo, M.S.; Santos, S.P.; Barros-Filho, B.A.; Lima, M.M.O.; Leite, A.S. Toxicogenetic potential of Mimosa pigra (Fabaceae) infusion in Allium cepa meristematic cells. Genet. Mol. Res. 2020, 19, gmr18588. [Google Scholar] [CrossRef]
- Koodkaew, I.; Wannathong, R. Effects of Mimosa pigra L. leaf extract on growth behavior of Ruellia tuberosa L. and Echinochloa crus-galli (L.) P. Beauv. Asia-Pac. J. Sci. Technol. 2018, 23, APST-23-07. [Google Scholar]
- Koodkaew, I.; Senaphan, C.; Sengseang, N.; Suwanwong, S. Characterization of phytochemical profile and phytotoxic activity of Mimosa pigra L. Agric. Nat. Resour. 2018, 52, 162–168. [Google Scholar] [CrossRef]
- Lonsdale, W.M. Litterfall in an Australian population of Mimosa pigra, an invasive tropical shrub. J. Trop. Ecol. 1988, 4, 381–392. [Google Scholar] [CrossRef]
- Renz, J. Mimosine. Physiol. Chem. 1936, 244, 153–158. [Google Scholar] [CrossRef]
- Brewbaker, J.L.; Hylin, J.W. Variations in mimosine content among Leucaena species and related mimosaceae. Crop Sci. 1965, 5, 348–349. [Google Scholar] [CrossRef]
- Smith, I.K.; Fowden, L.A. Study of mimosine toxicity in plants. J. Exp. Bot. 1996, 17, 750–761. [Google Scholar] [CrossRef]
- Soedarjo, M.; Borthakur, D. Mimosine produced by the tree-legume Leucaena provides growth advantages to some Rhizobium strains that utilize it as a source of carbon and nitrogen. Plant Soil 1996, 186, 87–92. [Google Scholar] [CrossRef]
- Nguyen, B.C.; Tawata, S. The chemistry and biological activities of mimosine: A review. Phytother. Res. 2016, 30, 1230–1242. [Google Scholar] [CrossRef]
- Rakotomalala, G.; Agard, C.; Tonnerre, P.; Tesse, A.; Derbré, S.; Michalet, S.; Hamzaoui, J.; Rio, M.; Cario-Toumaniantz, C.; Richomme, P.; et al. Extract from Mimosa pigra attenuates chronic experimental pulmonary hypertension. J. Ethnopharm. 2013, 148, 106–116. [Google Scholar] [CrossRef]
- De Morais, C.B.; Scopel, M.; Pedrazza, G.P.R.; da Silva, F.K.; Dalla Lana, D.F.; Tonello, M.L.; Miotto, S.T.S.; Machado, M.M.; De Oliveira, L.F.S.; Fuentefria, A.M.; et al. Anti-dermatophyte activity of Leguminosae plants from Southern Brazil with emphasis on Mimosa pigra (Leguminosae). J. Mycol. Med. 2017, 27, 530–538. [Google Scholar] [CrossRef]
- Rizwan, K.; Majeed, I.; Bilal, M.; Rasheed, T.; Shakeel, A.; Iqbal, S. Phytochemistry and diverse pharmacology of Genus Mimosa: A review. Biomolecules 2022, 12, 83. [Google Scholar] [CrossRef]
- Okonkwo, C.J.; Njoku, O.U.; Okonkwo, T.J.; Afieroho, O.E.; Proksch, P. Two new acylated flavonol glycosides from Mimosa pigra L. leaves sub-family Mimosoideae. Future J. Pharm. Sci. 2016, 2, 71–75. [Google Scholar] [CrossRef]
- Hawwal, M.F.; Ali, Z.; Fantoukh, O.I.; Chittiboyina, A.G.; Khan, I.A. Phytochemical investigation of Mimosa pigra leaves, a sensitive species. Biochem. Syst. Ecol. 2021, 99, 104354. [Google Scholar] [CrossRef]
- Englert, J.; Weniger, B.; Lobstein, A.; Anton, R.; Krempp, E.; Guillaume, D.; Leroy, Y. Triterpenoid saponins from Mimosa pigra. J. Nat. Prod. 1995, 58, 1265–1269. [Google Scholar] [CrossRef]
- Nguyen, L.N.P.; Huu, D.M.N.; Dang, H.P.; Huynh, N.V.; Dang, P.H. A new furanochromone from the leaves of Mimosa pigra. Nat. Prod. Res. 2021, 35, 3963–3969. [Google Scholar] [CrossRef] [PubMed]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147. [Google Scholar] [CrossRef]
- Weston, L.A.; Mathesius, U. Flavonoids: Their structure, biosynthesis and role in the rhizosphere, including allelopathy. J. Chem. Ecol. 2013, 39, 283–297. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Santonja, M.; Le Rouzic, B.; Thiébaut, G. Seasonal dependence and functional implications of macrophyte-phytoplankton allelopathic interactions. Freshw. Biol. 2018, 63, 1161–1172. [Google Scholar] [CrossRef]
- Thouvenot, L.; Haury, J.; Thiébaut, G. A success story: Water primroses, aquatic plant pests. Aquatic Conservation Mar. Freshw. Ecosyst. 2013, 23, 790–803. [Google Scholar] [CrossRef]
- Okada, S.; Iwasaki, A.; Kataoka, I.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of a phytotoxic substance in kiwifruit leaves. Acta Hortic. 2018, 1218, 207–212. [Google Scholar] [CrossRef]
- Parvez, M.M.K.; Yokotani, T.; Fujii, Y.; Konishi, T.; Iwashina, T. Effects of quercetin and its seven derivatives on the growth of Arabidopsis thaliana and Neurospora crassa. Biochem. Syst. Ecol. 2004, 32, 631–635. [Google Scholar] [CrossRef]
- Moody, M.E.; Mack, R.N. Controlling the spread of plant invasions: The importance of nascent foci. J. Appl. Ecol. 1988, 25, 1009–1021. [Google Scholar] [CrossRef]
- Lake, E.C.; Minteer, C.R. A review of the integration of classical biological control with other techniques to manage invasive weeds in natural areas and rangelands. BioControl 2018, 63, 71–86. [Google Scholar] [CrossRef]
- Siriworakul, M.; Schultz, G.C. Physical and Mechanical Control of Mimosa pigra. In A Guide to the Management of Mimosa pigra; Harley, K.L.S., Ed.; CSIRO: Canberra, Australia, 1992; pp. 102–103. [Google Scholar]
- DiTomaso, J.M.; Kyser, G.B.; Miller, J.R.; Garcia, S.; Smith, R.F.; Nader, G.; Connor, J.M.; Orloff, S.B. Integrating prescribed burning and clopyralid for the management of yellow starthistle (Centaurea solstitialis). Weed Sci. 2006, 54, 757–767. [Google Scholar] [CrossRef]
- Drus, G.M.; Dudley, T.L.; Brooks, M.L.; Matchett, J. The effect of leaf beetle herbivory on the fire behaviour of tamarisk (Tamarix ramosissima Lebed.). Int. J. Wildland Fire 2013, 22, 446–458. [Google Scholar] [CrossRef]
- Miller, I.L.; Siriworakul, M. Herbicide research and recommendations for control of Mimosa pigra. In A Guide to the Management of Mimosa pigra; Harley, K.L.S., Ed.; CSIRO: Canberra, Australia, 1992; pp. 86–91. [Google Scholar]
- Department of Land Resource Management. Weed Management Plan for Mimosa (Mimosa pigra). 2013. Available online: https://denr.nt.gov.au/__data/assets/pdf_file/0016/400372/Final-Weed-Management-Plan-for-Mimosa-Dec-2013.pdf (accessed on 14 April 2022).
- Lane, A.M.; Williams, R.J.; Muller, W.J.; Lonsdale, W.M. The effects of the herbicide tebuthiuron on seedlings of Mimosa pigra and native floodplain. Austral. J. Ecol. 1997, 22, 439–447. [Google Scholar] [CrossRef]
- Ostermeyer, N.; Grace, B.S. Establishment, distribution and abundance of Mimosa pigra biological control agents in northern Australia: Implications for biological control. BioControl 2007, 52, 703–720. [Google Scholar] [CrossRef]
- Burrows, N.; Lukitsch, B. Biological control agents are observed on Mimosa pigra six and 12 years after their release in the Northern Territory, Australia. In Proceedings of the 18th Australasian Weeds Conference, Melbourne, Australia, 8–11 October 2012; Weed Society of Victoria Inc.: Melbourne, Australia, 2012; pp. 347–348. [Google Scholar]
- Paynter, Q. Evaluating the impact of a biological control agent Carmenta mimosa on the woody wetland weed Mimosa pigra in Australia. J. Appl. Ecol. 2005, 42, 1054–1062. [Google Scholar] [CrossRef]
- Wilson, C.G.; Flanagan, G.J. Establishment and spread of Neurostrota gunniella on Mimosa pigra in the Northern Territory. In Proceedings of the 9th Australasian Weeds Conference, Adelaide, Australia, 6–10 August 1990; Crop Science Society of South Australia Inc.: Adelaide, Australia, 1992; pp. 505–507. [Google Scholar]
- Davis, D.R.; Kassulke, R.C.; Harley, K.L.S.; Gillett, J.D. Systematic, morphology, biology, and host specificity of Neurostrota gunniella (Busk) (Lepidoptera: Gracillariidae), an agent for the biological control of Mimosa pigra L. Proc. Entomol. Soc. Wash. 1991, 93, 16–44. [Google Scholar]
- Paynter, Q.; Hennecke, B. Competition between two biological control agents, Neurostrota gunniella and Phloeospora mimosae-pigrae, and their impact on the invasive tropical shrub Mimosa pigra. Biocontrol Sci. Technol. 2001, 11, 575–582. [Google Scholar] [CrossRef]
- Paynter, Q. Evaluating the impact of biological control against Mimosa pigra in Australia: Comparing litterfall before and after the introduction of biological control agents. Biol. Control. 2006, 38, 166–173. [Google Scholar] [CrossRef]
- Heard, T.A.; Elliott, L.P.; Anderson, B.; White, L.; Burrows, N.; Mira, A.; Zonneveld, R.; Fichera, G.; Chan, R.; Segura, R. Biology, host specificity, release and establishment of Macaria pallidata and Leuciris fimbriaria (Lepidoptera: Geometridae), biological control agents of the weed Mimosa pigra. Biol. Control. 2010, 55, 248–255. [Google Scholar] [CrossRef]
- Heard, T.A.; Forno, W.I. Host selection and host range of the flower-feeding weevil, Coelocephalapion pigrae, a potential biological control agent of Mimosa pigra. Biol. Control. 1996, 6, 83–95. [Google Scholar] [CrossRef]
- Heard, T.A.; Paynter, Q.; Chan, R.; Mira, A. Malacorhinus irregularis for biological control of Mimosa pigra: Host specificity, life cycle and establishment in Australia. Biol. Control. 2005, 32, 252–262. [Google Scholar] [CrossRef]
- Burrows, N.J.; Lukitsch, B.V.; Liberato, J.R. Rediscovery of the rust Diabole cubensis, released as a classical biological control agent against the invasive weed Mimosa pigra in Australia. Australas. Plant Dis. Notes 2012, 7, 171–175. [Google Scholar] [CrossRef]
- Suasa-ard, W.; Sommartya, P.; Jaitui, S. Evaluation of seed-feeding bruchids, Acanthoscelides species, as biological control agents for Mimosa pigra in Thailand. In Research and Management of Mimosa pigra; Julien, M., Flanagan, G., Heard, T., Hennecke, B., Paynter, Q., Wilson, C., Eds.; CSIRO Entomology: Canberra, Australia, 2004; pp. 122–125. [Google Scholar]
- Buckley, Y.; Rees, M.; Paynter, Q.; Lonsdale, M. Modelling integrated weed management of an invasive shrub in tropical Australia, Mimosa pigra. J. Appl. Ecol. 2004, 41, 547–560. [Google Scholar] [CrossRef]
- van Driesche, R. The role of biological control in wildlands. BioControl 2012, 57, 131–137. [Google Scholar] [CrossRef]
- Mosley, J.C.; Frost, R.A.; Roeder, B.L.; Mosley, T.K.; Marks, G. Combined herbivory by targeted sheep grazing and biological control insects to suppress spotted knapweed (Centaurea stoebe). Invasive Plant Sci. Manag. 2016, 9, 22–32. [Google Scholar] [CrossRef]
- Hatcher, P.; Melander, B. Combining physical, cultural and biological methods: Prospects for integrated non-chemical weed management strategies. Weed Res. 2003, 43, 303–322. [Google Scholar] [CrossRef]
- Lym, R.G. Integration of biological control agents with other weed management technologies: Successes from the leafy spurge (Euphorbia esula) IPM program. Biol. Control 2005, 35, 366–375. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato-Noguchi, H. Invasive Mechanisms of One of the World’s Worst Alien Plant Species Mimosa pigra and Its Management. Plants 2023, 12, 1960. https://doi.org/10.3390/plants12101960
Kato-Noguchi H. Invasive Mechanisms of One of the World’s Worst Alien Plant Species Mimosa pigra and Its Management. Plants. 2023; 12(10):1960. https://doi.org/10.3390/plants12101960
Chicago/Turabian StyleKato-Noguchi, Hisashi. 2023. "Invasive Mechanisms of One of the World’s Worst Alien Plant Species Mimosa pigra and Its Management" Plants 12, no. 10: 1960. https://doi.org/10.3390/plants12101960