
Transcriptome Analysis of the Late-Acting Self-Incompatibility Associated with RNase T2 Family in Camellia oleifera

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Determination of the Occurrence of SI and Fruit Set Evaluation
2.2. Transcriptome Assembly and Functional Annotation
2.3. Comparative Analysis of Differentially Expressed Genes (DEGs)
2.4. GO and KEGG Pathway Analysis of DEGs
2.5. DEGs Related to Plant Hormone Signal Transduction
2.6. DEGs Involved in ABC Transporters
2.7. DEGs Related to the Phosphatidylinositol Signaling System
2.8. DEGs Related to ROS and the Ca2+ Signaling Pathway
2.9. A Putative S-RNase Gene Involved in SI of C. oleifera
2.10. Identification and Characterization of T2 RNases in Camellia Oil Tree
2.11. Prokaryotic Expression Analysis of RNases
2.12. Subcellular Localization of RNases
2.13. Quantitative Real-Time PCR (qRT–PCR) Validation
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Pollination Treatment and Sample Collection
4.2. Data Surveys and Cytological Observations
4.3. RNA-Seq, Transcriptome Assembly and Functional Annotations
4.4. Gene Cloning, Multiple Sequence Alignment and Phylogenetic Analysis
4.5. Recombinant Protein Expression
4.6. qRT–PCR Verification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, F.; Li, Z.; Zhou, J.; Gu, Y.; Tan, X. Comparative study on fruit development and oil synthesis in two cultivars of Camellia oleifera. BMC Plant Biol. 2021, 21, 348. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Wu, B.; Hong, W.; Li, X.; Li, Z.; Xue, L.; Huang, Y. Transcriptome analysis of the tea oil camellia (Camellia oleifera) reveals candidate drought stress genes. PLoS ONE 2017, 12, e0181835. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Choi, K.; Kim, G.-B.; Yu, H.-J.; Cho, A.; Jang, H.; Kim, C.; Kim, H.-J.; Chang, K.S.; Kim, J.-H.; et al. Draft genome sequence of wild Prunus yedoensis reveals massive inter-specific hybridization between sympatric flowering cherries. Genome Biol. 2018, 19, 127. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Wang, S.; Liu, Y.; Li, J.; Zhong, H.; Zou, F.; Yuan, D. Hydrogen cyanamide enhances flowering time in tea oil camellia (Camellia oleifera Abel.). Ind. Crop. Prod. 2021, 176, 114313. [Google Scholar] [CrossRef]
- He, Y.; Chen, R.; Yang, Y.; Liang, G.; Zhang, H.; Deng, X.; Xi, R. Sugar Metabolism and Transcriptome Analysis Reveal Key Sugar Transporters during Camellia oleifera Fruit Development. Int. J. Mol. Sci. 2022, 23, 822. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Song, Q.; Wu, Y.; Ye, S.; Chen, S.; Chen, H. TMT-Based Quantitative Proteomic Analysis Reveals the Crucial Biological Pathways Involved in Self-Incompatibility Responses in Camellia oleifera. Int. J. Mol. Sci. 2020, 21, 1987. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Wang, K.; Wang, Y.; Hu, Z.; Yan, C.; Huang, H.; Ma, X.; Cao, Y.; Long, W.; Liu, W.; et al. The genome of oil-Camellia and population genomics analysis provide insights into seed oil domestication. Genome Biol. 2022, 23, 14. [Google Scholar] [CrossRef]
- Zhou, J.; Lu, M.; Yu, S.; Liu, Y.; Yang, J.; Tan, X. In-Depth Understanding of Camellia oleifera Self-Incompatibility by Comparative Transcriptome, Proteome and Metabolome. Int. J. Mol. Sci. 2020, 21, 1600. [Google Scholar] [CrossRef]
- He, Y.; Song, Q.; Chen, S.; Wu, Y.; Zheng, G.; Feng, J.; Yang, Z.; Lin, W.; Li, Y.; Chen, H. Transcriptome analysis of self- and cross-pollinated pistils revealing candidate unigenes of self-incompatibility in Camellia oleifera. J. Hortic. Sci. Biotechnol. 2019, 95, 19–31. [Google Scholar] [CrossRef]
- Zhang, C.-C.; Wang, L.-Y.; Wei, K.; Wu, L.-Y.; Li, H.-L.; Zhang, F.; Cheng, H.; Ni, D.-J. Transcriptome analysis reveals self-incompatibility in the tea plant (Camellia sinensis) might be under gametophytic control. BMC Genom. 2016, 17, 359. [Google Scholar] [CrossRef]
- Tan, L.-Q.; Wang, L.-Y.; Wei, K.; Zhang, C.-C.; Wu, L.-Y.; Qi, G.-N.; Cheng, H.; Zhang, Q.; Cui, Q.-M.; Liang, J.-B. Floral Transcriptome Sequencing for SSR Marker Development and Linkage Map Construction in the Tea Plant (Camellia sinensis). PLoS ONE 2013, 8, e81611. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, Q.; Gu, Z.; Wu, C.; Meng, D.; Yu, J.; Chen, Q.; Li, Y.; Yuan, H.; Wang, D.; et al. Molecular and genetic characterization of a self-compatible apple cultivar, ‘CAU-1’. Plant Sci. 2016, 252, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Herrera, S.; Rodrigo, J.; Hormaza, J.I.; Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int. J. Mol. Sci. 2018, 19, 3612. [Google Scholar] [CrossRef]
- Gibbs, P.E. Late-acting self-incompatibility–the pariah breeding system in flowering plants. New Phytol. 2014, 203, 717–734. [Google Scholar] [CrossRef]
- LaDoux, T.; Friar, E.A. Late-Acting Self-Incompatibility in Ipomopsis tenuifolia (Gray) V. Grant (Polemoniaceae). Int. J. Plant Sci. 2006, 167, 463–471. [Google Scholar] [CrossRef]
- Bagheri, M.; Ershadi, A. Self-incompatibility alleles in Iranian pear cultivars. Biocatal. Agric. Biotechnol. 2020, 27, 101672. [Google Scholar] [CrossRef]
- Ma, J.-Z.; Wang, H.-J.; Wang, D.-S.; Li, X.-Y.; Liu, J.-Z.; Xiao, X.; Zhang, L.-B.; Wu, J.-K. Self-compatibility of Crataegus pinnatifida Bge. ‘Zizhenzhu’ is associated with the mutation of a pistil-part non-S factor. Sci. Hortic. 2020, 268, 109362. [Google Scholar] [CrossRef]
- Li, Y.; Wu, J.; Wu, C.; Yu, J.; Liu, C.; Fan, W.; Li, T.; Li, W. A mutation near the active site of S-RNase causes self-compatibility in S-RNase-based self-incompatible plants. Plant Mol. Biol. 2020, 103, 129–139. [Google Scholar] [CrossRef]
- Yang, Q.; Meng, D.; Gu, Z.; Li, W.; Chen, Q.; Li, Y.; Yuan, H.; Yu, J.; Liu, C.; Li, T. Apple S -RNase interacts with an actin-binding protein, MdMVG, to reduce pollen tube growth by inhibiting its actin-severing activity at the early stage of self-pollination induction. Plant J. 2018, 95, 41–56. [Google Scholar] [CrossRef]
- Li, T.; Long, S.; Li, M.; Bai, S.; Zhang, W. Determination S-Genotypes and Identification of Five Novel S-RNase Alleles in Wild Malus Species. Plant Mol. Biol. Rep. 2011, 30, 453–461. [Google Scholar] [CrossRef]
- Wu, J.; Li, M.; Li, T. Genetic Features of the Spontaneous Self-Compatible Mutant, ‘Jin Zhui’ (Pyrus bretschneideri Rehd.). PLoS ONE 2013, 8, e76509. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Cao, Z.; Zhu, A.; Liu, Y.; Tao, M.; Yang, H.; Xu, Q.; Wang, S.; Liu, J.; Li, Y.; et al. Evolution of self-compatibility by a mutant Sm-RNase in citrus. Nat. Plants 2020, 6, 131–142. [Google Scholar] [CrossRef]
- Wu, L.; Williams, J.S.; Sun, L.; Kao, T. Sequence analysis of the Petunia inflata S-locus region containing 17 S-Locus F-Box genes and the S-RNase gene involved in self-incompatibility. Plant J. 2020, 104, 1348–1368. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hao, S.; Wang, L.; Fang, W.; Wang, Y.; Li, X. Late-acting self-incompatibility in tea plant (Camellia sinensis). Biologia 2012, 67, 347–351. [Google Scholar] [CrossRef]
- Ford, C.S.; Wilkinson, M.J. Confocal observations of late-acting self-incompatibility in Theobroma cacao L. Sex. Plant Reprod. 2012, 25, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, P.; De Graaf, B.H.J.; Zhang, H.; Jiao, H.; Tang, C.; Zhang, S.; Wu, J. Phosphatidic Acid Counteracts S-RNase Signaling in Pollen by Stabilizing the Actin Cytoskeleton. Plant Cell 2018, 30, 1023–1039. [Google Scholar] [CrossRef]
- Wilkins, K.A.; Bancroft, J.; Bosch, M.; Ings, J.; Smirnoff, N.; Franklin-Tong, V.E. Reactive Oxygen Species and Nitric Oxide Mediate Actin Reorganization and Programmed Cell Death in the Self-Incompatibility Response of Papaver. Plant Physiol. 2011, 156, 404–416. [Google Scholar] [CrossRef]
- Serrano, I.; Romero-Puertas, M.C.; Sandalio, L.M.; Olmedilla, A. The role of reactive oxygen species and nitric oxide in programmed cell death associated with self-incompatibility. J. Exp. Bot. 2015, 66, 2869–2876. [Google Scholar] [CrossRef]
- Del Duca, S.; Aloisi, I.; Parrotta, L.; Cai, G. Cytoskeleton, Transglutaminase and Gametophytic Self-Incompatibility in the Malinae (Rosaceae). Int. J. Mol. Sci. 2019, 20, 209. [Google Scholar] [CrossRef]
- Chang, Y.; Gong, W.; Xu, J.; Gong, H.; Song, Q.; Xiao, S.; Yuan, D. Integration of semi-in vivo assays and multi-omics data reveals the effect of galloylated catechins on self-pollen tube inhibition in Camellia oleifera. Hortic. Res. 2023, 10, uhac248. [Google Scholar] [CrossRef]
- Qin, N.; Gao, Y.; Cheng, X.; Yang, Y.; Wu, J.; Wang, J.; Li, S.; Xing, G. Genome-wide identification of CLE gene family and their potential roles in bolting and fruit bearing in cucumber (Cucumis sativus L.). BMC Plant Biol. 2021, 21, 143. [Google Scholar] [CrossRef]
- Sheng, H.; Zhang, S.; Wei, Y.; Chen, S. Exogenous Application of Low-Concentration Sugar Enhances Brassinosteroid Signaling for Skotomorphogenesis by Promoting BIN2 Degradation. Int. J. Mol. Sci. 2021, 22, 13588. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Tang, C.; Wang, R.; Gu, C.; Wu, X.; Hu, S.; Jiao, J.; Zhang, S. Transcriptome and phytohormone analysis reveals a comprehensive phytohormone and pathogen defence response in pear self-/cross-pollination. Plant Cell Rep. 2017, 36, 1785–1799. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Gu, Z.; Li, W.; Wang, A.; Yuan, H.; Yang, Q.; Li, T. Apple MdABCF assists in the transportation of S-RNase into pollen tubes. Plant J. 2014, 78, 990–1002. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.; Nakajima, R.; Iwano, M.; Kanaoka, M.M.; Kimura, S.; Takeda, S.; Kawarazaki, T.; Senzaki, E.; Hamamura, Y.; Higashiyama, T.; et al. Ca2+-Activated Reactive Oxygen Species Production by Arabidopsis RbohH and RbohJ Is Essential for Proper Pollen Tube Tip Growth. Plant Cell 2014, 26, 1069–1080. [Google Scholar] [CrossRef]
- Li, C.-Y.; Zhang, N.; Guan, B.; Zhou, Z.-Q.; Mei, F.-Z. Reactive oxygen species are involved in cell death in wheat roots against powdery mildew. J. Integr. Agric. 2019, 18, 1961–1970. [Google Scholar] [CrossRef]
- Yoon, G.M.; Dowd, P.E.; Gilroy, S.; McCubbin, A.G. Calcium-Dependent Protein Kinase Isoforms in Petunia Have Distinct Functions in Pollen Tube Growth, Including Regulating Polarity. Plant Cell 2006, 18, 867–878. [Google Scholar] [CrossRef]
- Entani, T.; Kubo, K.-I.; Isogai, S.; Fukao, Y.; Shirakawa, M.; Isogai, A.; Takayama, S. Ubiquitin-proteasome-mediated degradation of S-RNase in a solanaceous cross-compatibility reaction. Plant J. 2014, 78, 1014–1021. [Google Scholar] [CrossRef]
- Abdallah, D.; Baraket, G.; Perez, V.; Ben Mustapha, S.; Salhi-Hannachi, A.; Hormaza, J.I. Analysis of Self-Incompatibility and Genetic Diversity in Diploid and Hexaploid Plum Genotypes. Front. Plant Sci. 2019, 10, 896. [Google Scholar] [CrossRef]
- Honsho, C.; Ushijima, K.; Anraku, M.; Ishimura, S.; Yu, Q.; Gmitter, F.G., Jr.; Tetsumura, T. Association of T2/S-RNase With Self-Incompatibility of Japanese Citrus Accessions Examined by Transcriptomic, Phylogenetic, and Genetic Approaches. Front. Plant Sci. 2021, 12, 638321. [Google Scholar] [CrossRef]
- Qin, X.M.; Zhang, Y.; Liu, Y.J.; Ni Guo, D.; Li, H.M. Molecular mechanisms underlying the participation of Ribonuclease T2 gene into self-incompatibility of Citrus grandis var. Shatianyu Hort. Cell. Mol. Biol. 2018, 64, 1153. [Google Scholar] [CrossRef] [PubMed]
- MacIntosh, G.C.; Hillwig, M.S.; Meyer, A.; Flagel, L. RNase T2 genes from rice and the evolution of secretory ribonucleases in plants. Mol. Genet. Genom. 2010, 283, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Yang, W.; Su, S.; Fu, L.; Yi, H.; Chen, C.; Deng, X.; Chai, L. Genome-wide identification and functional analysis of S-RNase involved in the self-incompatibility of citrus. Mol. Genet. Genom. 2016, 292, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Luhtala, N.; Parker, R. T2 Family ribonucleases: Ancient enzymes with diverse roles. Trends Biochem. Sci. 2010, 35, 253–259. [Google Scholar] [CrossRef]
- Manghwar, H.; Hussain, A.; Ali, Q.; Liu, F. Brassinosteroids (BRs) Role in Plant Development and Coping with Different Stresses. Int. J. Mol. Sci. 2022, 23, 1012. [Google Scholar] [CrossRef]
- Zhang, J.; Qian, J.-Y.; Bian, Y.-H.; Liu, X.; Wang, C.-L. Transcriptome and Metabolite Conjoint Analysis Reveals the Seed Dormancy Release Process in Callery Pear. Int. J. Mol. Sci. 2022, 23, 2186. [Google Scholar] [CrossRef]
- Song, Y.; Feng, L.; Alyafei, M.A.M.; Jaleel, A.; Ren, M. Function of Chloroplasts in Plant Stress Responses. Int. J. Mol. Sci. 2021, 22, 13464. [Google Scholar] [CrossRef]
- Lu, M.; Zhou, J.; Liu, Y.; Yang, J.; Tan, X. CoNPR1 and CoNPR3.1 are involved in SA- and MeSA- mediated growth of the pollen tube in Camellia oleifera. Physiol. Plant. 2021, 172, 2181–2190. [Google Scholar] [CrossRef]
- Zakharova, E.V.; Timofeeva, G.V.; Fateev, A.D.; Kovaleva, L.V. Caspase-like proteases and the phytohormone cytokinin as determinants of S-RNAse–based self-incompatibility–induced PCD in Petunia hybrida L. Protoplasma 2020, 258, 573–586. [Google Scholar] [CrossRef]
- Liu, C.; Xiao, P.; Jiang, F.; Wang, S.; Liu, Z.; Song, G.; Li, W.; Lv, T.; Li, J.; Wang, D.; et al. Exogenous gibberellin treatment improves fruit quality in self-pollinated apple. Plant Physiol. Biochem. 2022, 174, 11–21. [Google Scholar] [CrossRef]
- Gu, Z.; Li, W.; Doughty, J.; Meng, D.; Yang, Q.; Yuan, H.; Li, Y.; Chen, Q.; Yu, J.; Liu, C.S.; et al. A gamma-thionin protein from apple, MdD1, is required for defence against S-RNase-induced inhibition of pollen tube prior to self/non-self recognition. Plant Biotechnol. J. 2019, 17, 2184–2198. [Google Scholar] [CrossRef] [PubMed]
- Kucera, B.; Cohn, M.A.; Leubner-Metzger, G. Plant hormone interactions during seed dormancy release and germination. Seed Sci. Res. 2005, 15, 281–307. [Google Scholar] [CrossRef]
- Chang, E.; Deng, N.; Zhang, J.; Liu, J.; Chen, L.; Zhao, X.; Abbas, M.; Jiang, Z.; Shi, S. Proteome-Level Analysis of Metabolism- and Stress-Related Proteins during Seed Dormancy and Germination in Gnetum parvifolium. J. Agric. Food Chem. 2018, 66, 3019–3029. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, L.-Y.; Zhou, P.; Liao, Z.; Lin, H.; Yu, Q.; Ming, R. Sex biased expression of hormone related genes at early stage of sex differentiation in papaya flowers. Hortic. Res. 2021, 8, 147. [Google Scholar] [CrossRef]
- Baker, R.P.; Hasenstein, K.H.; Zavada, M.S. Hormonal Changes after Compatible and Incompatible Pollination in Theobroma cacao L. Hortscience 1997, 32, 1231–1234. [Google Scholar] [CrossRef]
- Guo, Z.; Yuan, X.; Li, L.; Zeng, M.; Yang, J.; Tang, H.; Duan, C. Genome-Wide Analysis of the ATP-Binding Cassette (ABC) Transporter Family in Zea mays L. and Its Response to Heavy Metal Stresses. Int. J. Mol. Sci. 2022, 23, 2109. [Google Scholar] [CrossRef]
- Niu, L.; Li, H.; Song, Z.; Dong, B.; Cao, H.; Liu, T.; Du, T.; Yang, W.; Amin, R.; Wang, L.; et al. The functional analysis of ABCG transporters in the adaptation of pigeon pea (Cajanus cajan) to abiotic stresses. PeerJ 2021, 9, e10688. [Google Scholar] [CrossRef]
- Qu, H.; Guan, Y.; Wang, Y.; Zhang, S. PLC-Mediated Signaling Pathway in Pollen Tubes Regulates the Gametophytic Self-incompatibility of Pyrus Species. Front. Plant Sci. 2017, 8, 1164. [Google Scholar] [CrossRef]
- Zhou, B.; Liu, Y.; Chen, Z.; Liu, D.; Wang, Y.; Zheng, J.; Liao, X.; Zhou, R. Comparative Transcriptome Analysis Reveals the Cause for Accumulation of Reactive Oxygen Species During Pollen Abortion in Cytoplasmic Male-Sterile Kenaf Line 722HA. Int. J. Mol. Sci. 2019, 20, 5515. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2013, 65, 1229–1240. [Google Scholar] [CrossRef]
- Tiwari, B.S.; Belenghi, B.; Levine, A. Oxidative Stress Increased Respiration and Generation of Reactive Oxygen Species, Resulting in ATP Depletion, Opening of Mitochondrial Permeability Transition, and Programmed Cell Death. Plant Physiol. 2002, 128, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, S.; Sun, H.; Liu, C.; Li, X.; Liu, Y.; Lyu, D.; Du, G. Production of reactive oxygen species by PuRBOHF is critical for stone cell development in pear fruit. Hortic. Res. 2021, 8, 249. [Google Scholar] [CrossRef] [PubMed]
- Hiscock, S.J.; Bright, J.; McInnis, S.M.; Desikan, R.; Hancock, J.T. Signaling on the Stigma: Potential New Roles for ROS and NO in Plant Cell Signaling. Plant Signal. Behav. 2007, 2, 23–24. [Google Scholar] [CrossRef]
- Qin, Y.; Yang, Z. Rapid tip growth: Insights from pollen tubes. Semin. Cell Dev. Biol. 2011, 22, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Konrad, K.R.; Wudick, M.M.; Feijó, J.A. Calcium regulation of tip growth: New genes for old mechanisms. Curr. Opin. Plant Biol. 2011, 14, 721–730. [Google Scholar] [CrossRef]
- Peer, L.A. Self-Incompatibility Patterns and Signal Transduction. In Plant Signaling: Understanding the Molecular Crosstalk; Hakeem, K.R., Rehman, R.U., Tahir, I., Eds.; Springer: New Delhi, India, 2014; pp. 327–343. ISBN 978-81-322-1541-7. [Google Scholar]
- Xu, C.; Li, M.; Wu, J.; Guo, H.; Li, Q.; Zhang, Y.; Chai, J.; Li, T.; Xue, Y. Identification of a canonical SCFSLF complex involved in S-RNase-based self-incompatibility of Pyrus (Rosaceae). Plant Mol. Biol. 2012, 81, 245–257. [Google Scholar] [CrossRef]
- Sun, L.; Williams, J.S.; Li, S.; Wu, L.; Khatri, W.A.; Stone, P.G.; Keebaugh, M.D.; Kao, T.-H. S-Locus F-Box Proteins Are Solely Responsible for S-RNase-Based Self-Incompatibility of Petunia Pollen. Plant Cell 2018, 30, 2959–2972. [Google Scholar] [CrossRef]
- Romero, C.; Vilanova, S.; Burgos, L.; Vicente, M.; Badenes, M.L. Analysis of the S-locus structure in Prunus armeniaca L. Identification ofS-haplotype specific S-RNase and F-box genes. Plant Mol. Biol. 2004, 56, 145–157. [Google Scholar] [CrossRef]
- Seth, R.; Bhandawat, A.; Parmar, R.; Singh, P.; Kumar, S.; Sharma, R.K. Global Transcriptional Insights of Pollen-Pistil Interactions Commencing Self-Incompatibility and Fertilization in Tea [Camellia sinensis (L.) O. Kuntze]. Int. J. Mol. Sci. 2019, 20, 539. [Google Scholar] [CrossRef]
- Liao, T.; Yuan, D.-Y.; Zou, F.; Gao, C.; Yang, Y.; Zhang, L.; Tan, X.-F. Self-Sterility in Camellia oleifera May Be Due to the Prezygotic Late-Acting Self-Incompatibility. PLoS ONE 2014, 9, e99639. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]









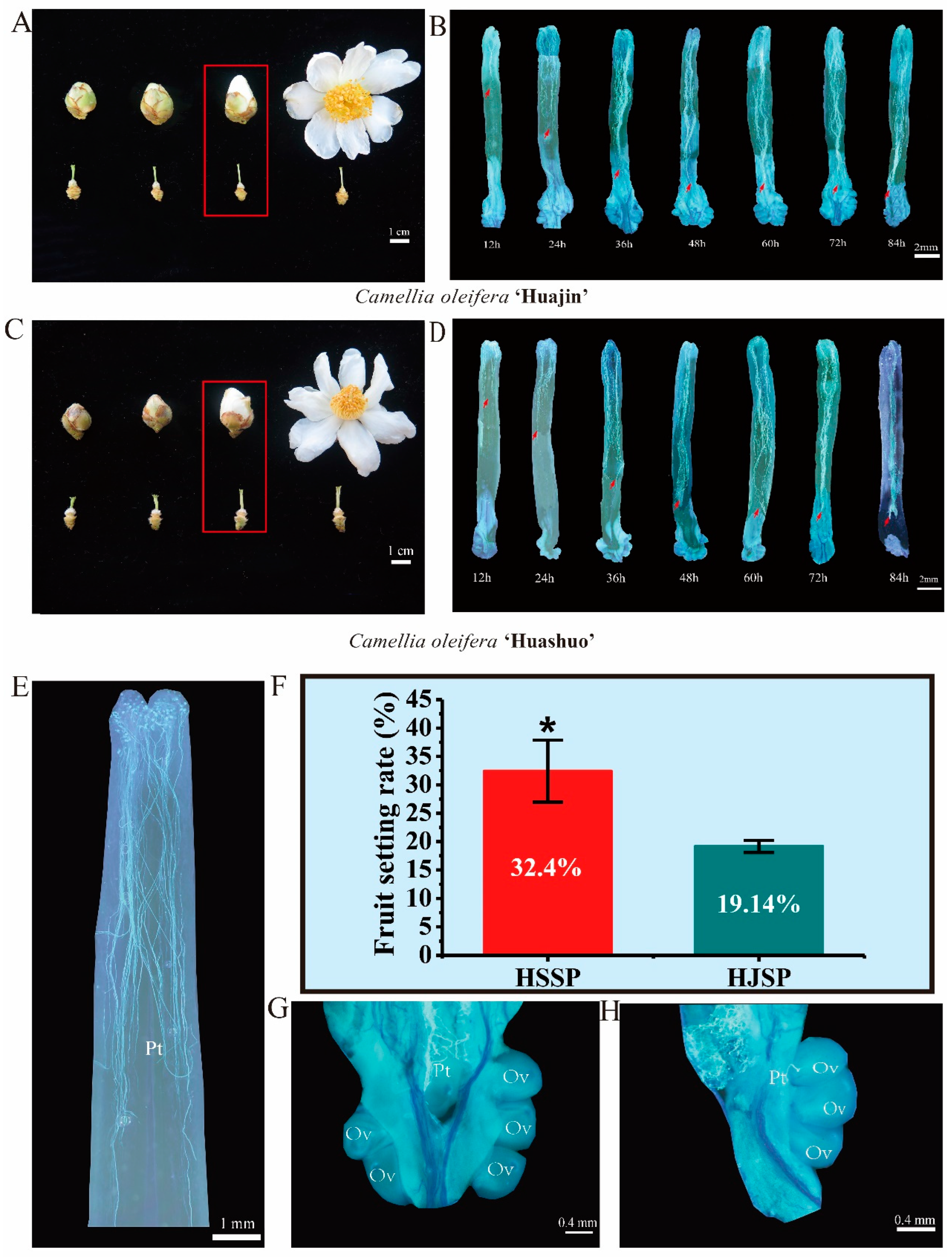{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
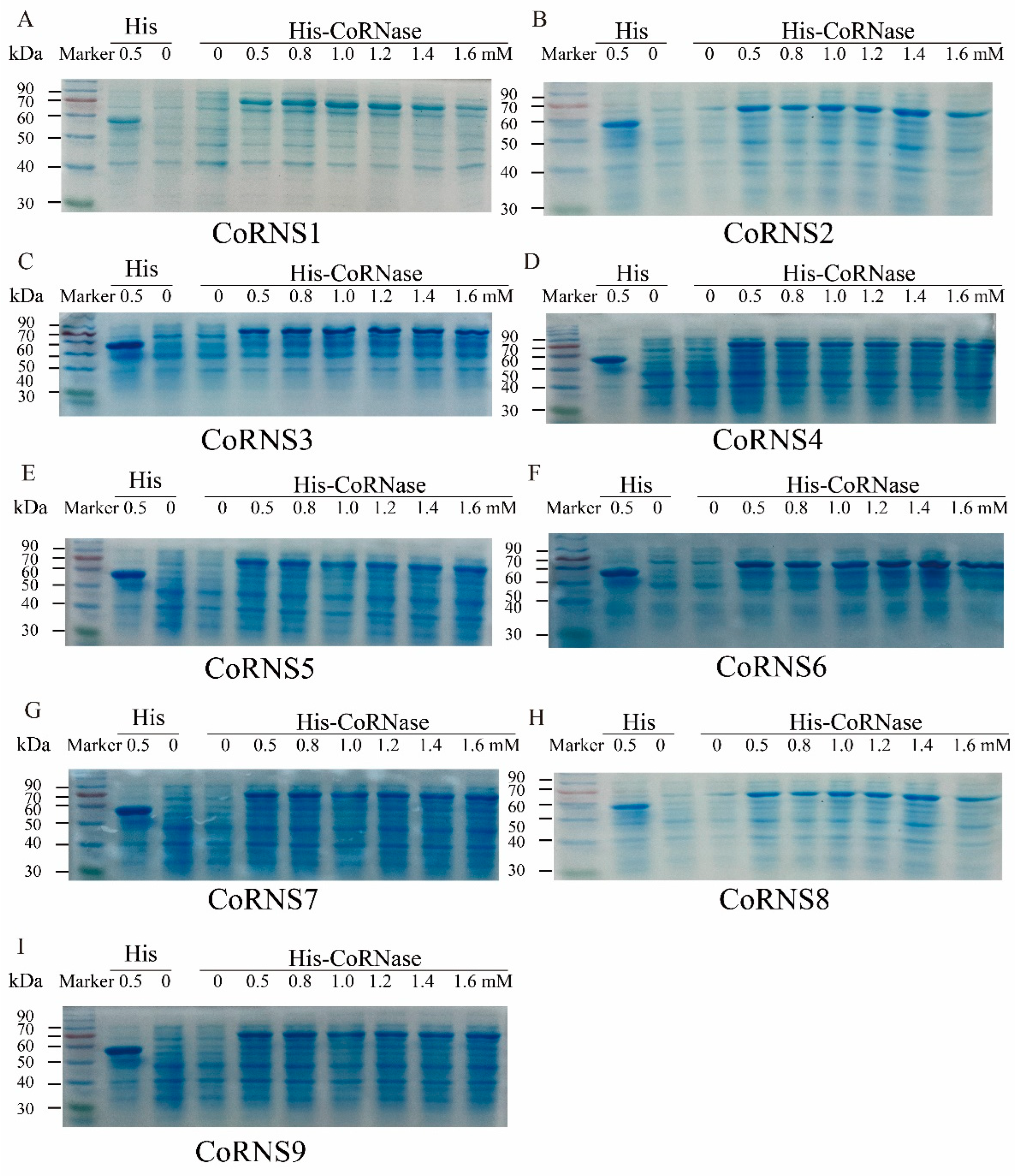{kind=link}
{kind=link}
{kind=link}
| Sample | Raw_Reads (Mb) | Clean_Reads (Mb) | Clean_Bases (Gb) | Q20 | Q30 | Unique_Map | Multi_Map | Total_Map |
|---|---|---|---|---|---|---|---|---|
| HJNP-1 | 66.78 | 66.50 | 9.92 | 98.00% | 94.16% | 39,267,036 (59.08%) | 21,089,363 (31.73%) | 60,356,399 (90.81%) |
| HJNP-2 | 69.22 | 69.02 | 10.10 | 98.10% | 94.41% | 40,222,163 (58.31%) | 21,918,936 (31.78%) | 62,141,099 (90.09%) |
| HJNP-3 | 61.92 | 61.71 | 9.02 | 97.87% | 93.82% | 36,100,019 (58.54%) | 19,422,333 (31.50%) | 55,522,352 (90.04%) |
| HJSP-1 | 47.98 | 47.89 | 7.15 | 98.33% | 94.74% | 28,096,446 (58.71%) | 15,250,400 (31.87%) | 43,346,846 (90.57%) |
| HJSP-2 | 50.43 | 50.33 | 7.52 | 98.15% | 94.26% | 29,748,589 (59.14%) | 15,790,304 (31.39%) | 45,538,893 (90.53%) |
| HJSP-3 | 52.69 | 52.60 | 7.86 | 98.07% | 94.04% | 30,664,191 (58.34%) | 16,592,787 (31.57%) | 47,256,978 (89.90%) |
| HSNP-1 | 61.12 | 60.95 | 9.09 | 97.94% | 93.91% | 38,592,078 (63.37%) | 18,130,734 (29.77%) | 56,722,812 (93.14%) |
| HSNP-2 | 62.36 | 62.18 | 9.27 | 97.77% | 93.62% | 39,226,394 (63.13%) | 18,632,072 (29.98%) | 57,858,466 (93.11%) |
| HSNP-3 | 72.42 | 72.21 | 10.76 | 97.95% | 94.07% | 44,810,287 (62.10%) | 22,744,249 (31.52%) | 67,554,536 (93.62%) |
| HSSP-1 | 44.88 | 44.80 | 6.70 | 98.28% | 94.59% | 28,700,957 (64.11%) | 13,046,307 (29.14%) | 41,747,264 (93.25%) |
| HSSP-2 | 61.69 | 61.58 | 9.21 | 98.12% | 94.19% | 39,366,886 (63.97%) | 17,537,945 (28.50%) | 56,904,831 (92.47%) |
| HSSP-3 | 57.18 | 57.09 | 8.64 | 98.26% | 94.52% | 36,229,624 (63.51%) | 16,581,192 (29.07%) | 52,810,816 (92.58%) |
| Name | ID | Length of CDS | Amino Acid | Molecular Weight | PI | Signal Peptide |
|---|---|---|---|---|---|---|
| CoRNS1 | oil_tea_GLEAN_10114714 | 748 | 315 | 34.82 | 5.6 | yes |
| CoRNS2 | oil_tea_GLEAN_10391745 | 429 | 142 | 15.65 | 6.02 | yes |
| CoRNS3 | oil_tea_GLEAN_10141428 | 480 | 159 | 17.75 | 5.39 | yes |
| CoRNS4 | oil_tea_GLEAN_10391744 | 448 | 143 | 16.0 | 4.94 | no |
| CoRNS5 | oil_tea_GLEAN_10004004 | 480 | 159 | 17.39 | 8.6 | no |
| CoRNS6 | oil_tea_GLEAN_10401437 | 753 | 250 | 28.39 | 8.77 | yes |
| CoRNS7 | oil_tea_GLEAN_10185682 | 684 | 227 | 24.97 | 4.59 | yes |
| CoRNS8 | oil_tea_GLEAN_10400090 | 717 | 262 | 26.54 | 5.42 | yes |
| CoRNS9 | oil_tea_GLEAN_10340465 | 457 | 146 | 16.1 | 5.78 | yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Lu, M.; Zhou, J.; Wang, S.; Long, Y.; Xu, Y.; Tan, X. Transcriptome Analysis of the Late-Acting Self-Incompatibility Associated with RNase T2 Family in Camellia oleifera. Plants 2023, 12, 1932. https://doi.org/10.3390/plants12101932
Li C, Lu M, Zhou J, Wang S, Long Y, Xu Y, Tan X. Transcriptome Analysis of the Late-Acting Self-Incompatibility Associated with RNase T2 Family in Camellia oleifera. Plants. 2023; 12(10):1932. https://doi.org/10.3390/plants12101932
Chicago/Turabian StyleLi, Chang, Mengqi Lu, Junqin Zhou, Sen Wang, Yi Long, Yan Xu, and Xiaofeng Tan. 2023. "Transcriptome Analysis of the Late-Acting Self-Incompatibility Associated with RNase T2 Family in Camellia oleifera" Plants 12, no. 10: 1932. https://doi.org/10.3390/plants12101932