
qPCR as a Selective Tool for Cytogenetics
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. The Study of Closely Related Species
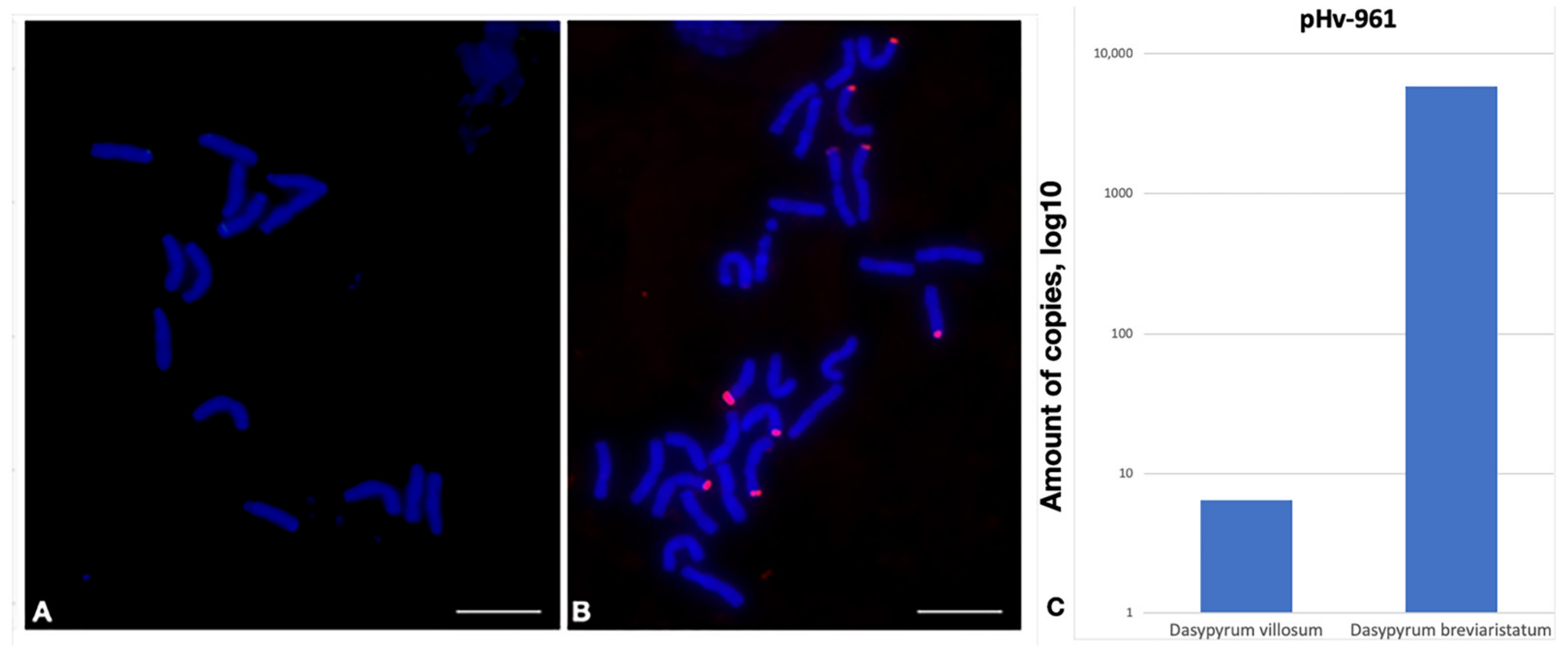
2.1.1. pHv-961
2.1.2. 19-202(a)
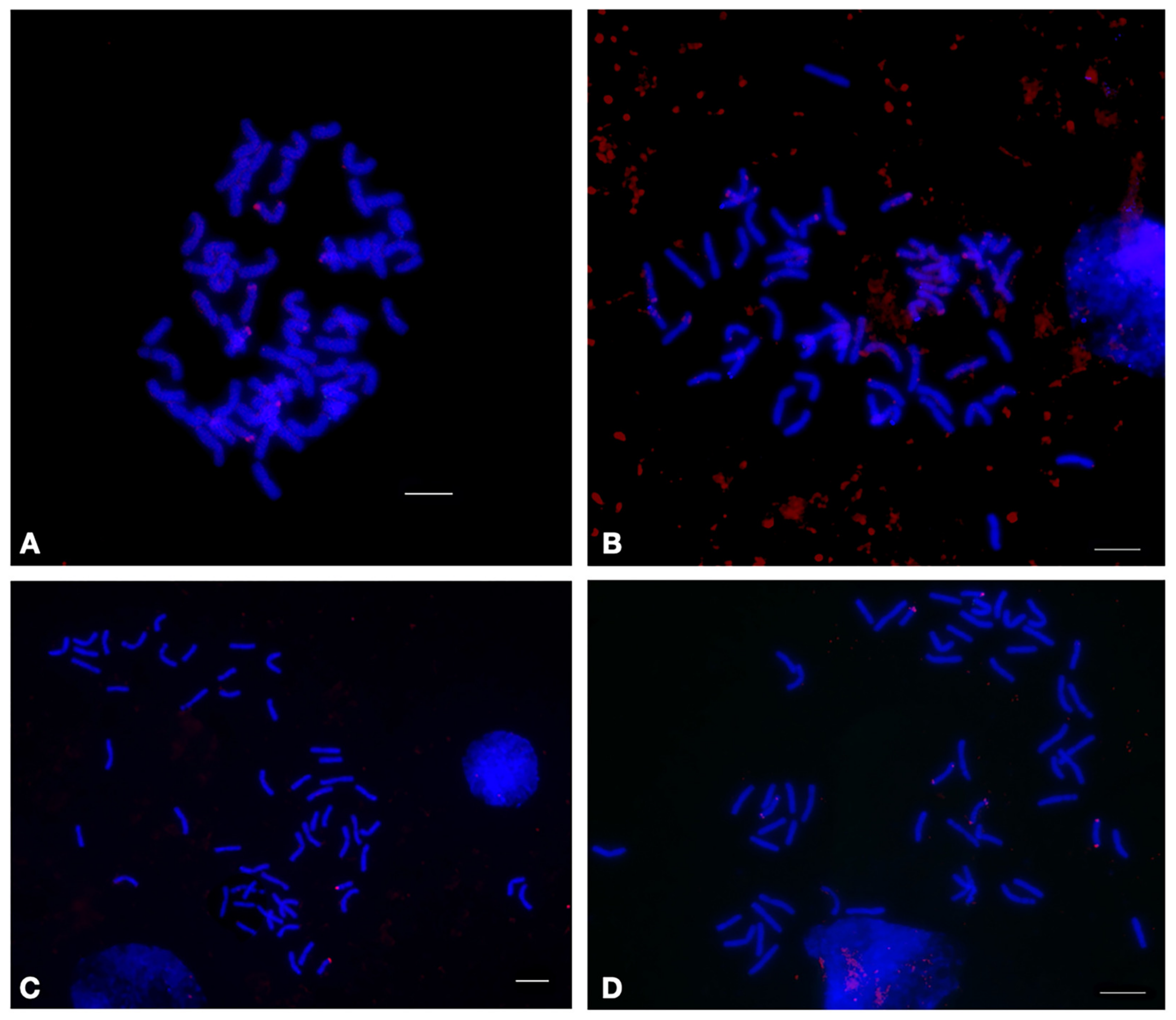
2.1.3. CL244
2.2. “Transfer of Repeats” Strategy for the Study of Closely Related Species
2.2.1. P720
2.2.2. 17-202
2.3. Analysis of Intraspecific Polymorphism
2.3.1. 17-62
2.3.2. 19-202(b)
2.4. Individual Cases
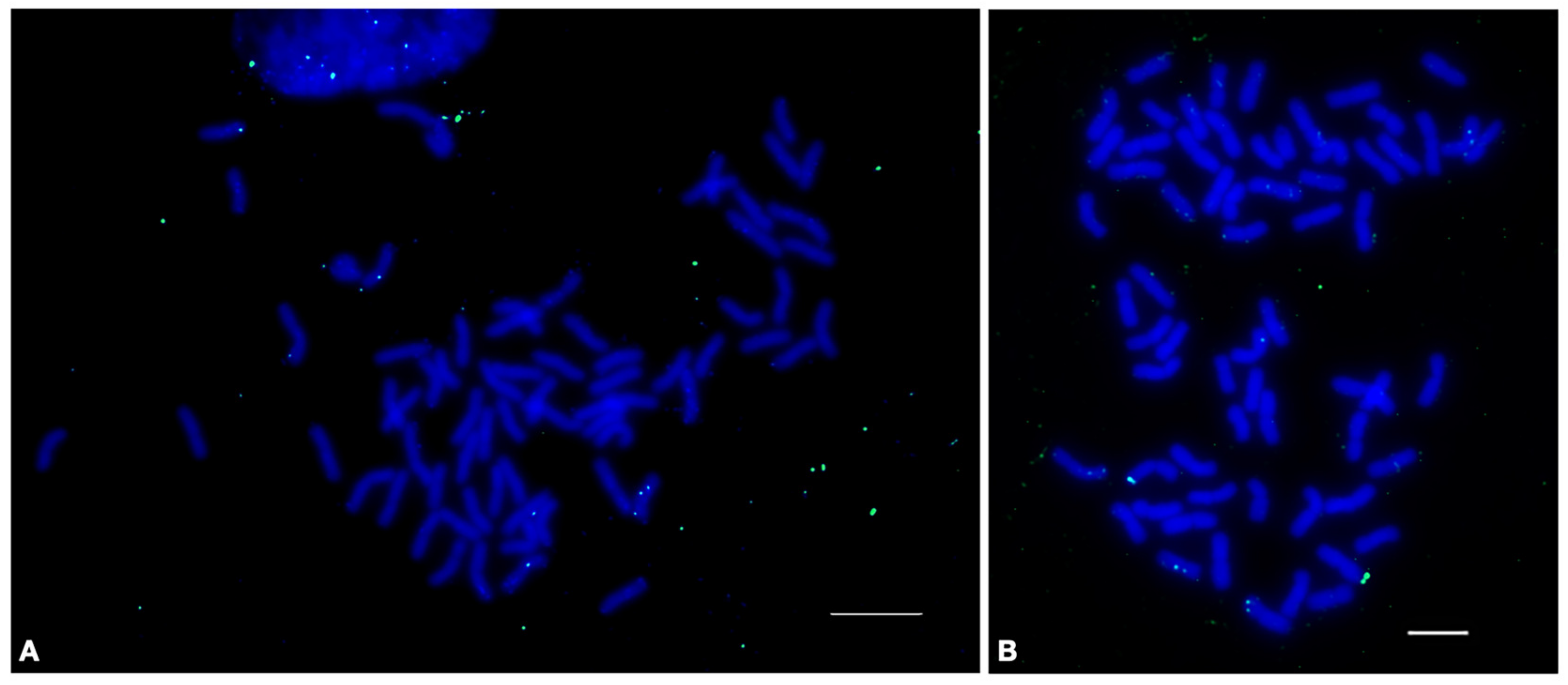
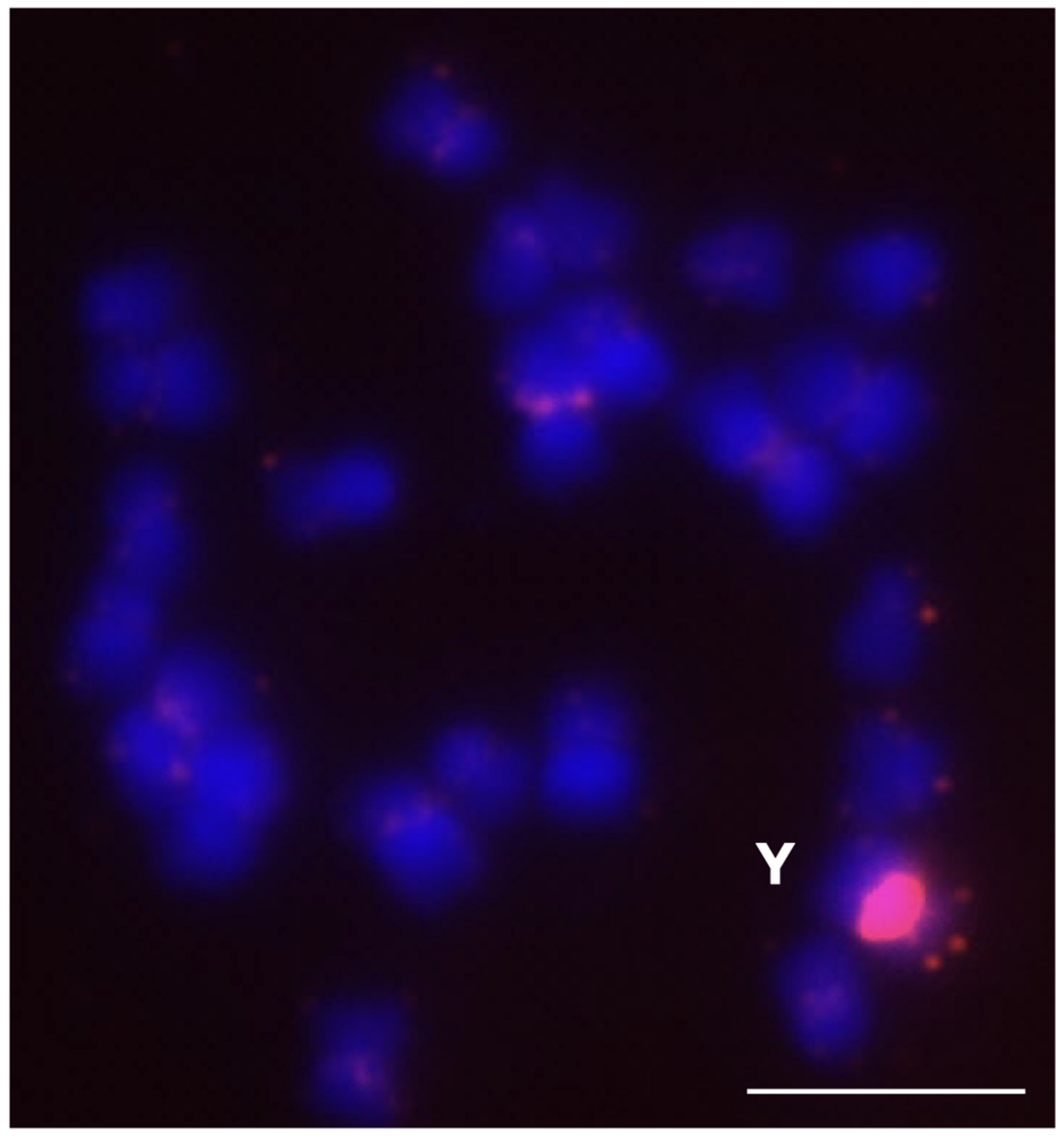
2.4.1. HRTR12
2.4.2. CL131
2.4.3. CL239
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Methods
- Finding repeats with RE2 and/or TRfinder
- qPCR with found tandem repeats
- FISH with selected tandem repeats
4.2.1. DNA Extraction
4.2.2. Sequencing and Preprocessing
4.2.3. Repeat Search
4.2.4. qPCR
4.2.5. FISH Protocols
5. Conclusions
- qPCR can be used as a downsampling tool for satellite repeats that are most promising for use as chromosomal markers. This approach makes it possible to develop chromosomal markers for one species based on whole-genome sequences of a related species.
- qPCR can detect tandem satellite repeats with several orders of magnitude difference in copy number between species, visualized by FISH. A correlation was shown between the results of bioinformatic analysis, qPCR, and FISH in most of the cases considered. In case of inconsistency in trends between qPCR and FISH results, we recommend checking the qPCR primers and designing other primers if necessary.
- Primers designed for the satellite repeat monomer can be used for both qPCR and probe preparation for the FISH procedure.
- Detected differences in qPCR copy number of satellite repeats allows for comparative analysis between closely related species, different populations of the same species, distant hybrids, and dioecious plants.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Komuro, S.; Endo, R.; Shikata, K.; Kato, A. Genomic and chromosomal distribution patterns of various repeated DNA sequences in wheat revealed by a fluorescence in situ hybridization procedure. Genome 2013, 56, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Danilova, T.V.; Friebe, B.; Gill, B.S. Single-copy gene fluorescence in situ hybridization and genome analysis: Acc-2 loci mark evolutionary chromosomal rearrangements in wheat. Chromosoma 2012, 121, 597–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuadrado, A.; Vitellozzi, F.; Jouve, N.; Ceoloni, C. Fluorescence in situ hybridization with multiple repeated DNA Probes applied to the analysis of wheat-rye chromosome pairing. Theor. Appl. Genet. 1997, 94, 347–355. [Google Scholar] [CrossRef]
- Qi, Z.; Du, P.; Qian, B.; Zhuang, L.; Chen, H.; Chen, T.; Shen, J.; Guo, J.; Feng, Y.; Pei, Z. Characterization of a wheat–Thinopyrum bessarabicum (T2JS-2BS·2BL) translocation line. Theor. Appl. Genet. 2010, 121, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Zhang, Z.H.; Liu, J.H.; Lu, J.Y.; Huang, S.W.; Jin, W.W. Distribution of the tandem repeat sequences and karyotyping in cucumber (Cucumis sativus L.) by fluorescence in situ hybridization. Cytogenet. Genome Res. 2008, 122, 80–88. [Google Scholar] [CrossRef]
- Lang, T.; Li, G.; Wang, H.; Yu, Z.; Chen, Q.; Yang, E.; Fu, S.; Tang, Z.; Yang, Z. Physical location of tandem repeats in the wheat genome and application for chromosome identification. Planta 2019, 249, 663–675. [Google Scholar] [CrossRef]
- Kroupin, P.Y.; Kuznetsova, V.M.; Nikitina, E.A.; Martirosyan, Y.T.; Karlov, G.I.; Divashuk, M.G. Development of new cytogenetic markers for Thinopyrum ponticum (Podp.) Z.-W. Liu & R.-C. Wang. Comp. Cytogenet. 2019, 13, 231–243. [Google Scholar] [CrossRef]
- Jiang, J.; Gill, B.S. Current status and the future of fluorescence in situ hybridization (FISH) in plant genome research. Genome 2006, 49, 1057–1068. [Google Scholar] [CrossRef]
- De Preter, K.; Speleman, F.; Combaret, V.; Lunec, J.; Laureys, G.; Eussen, B.H.J.; Francotte, N.; Board, J.; Pearson, A.D.J.; De Paepe, A.; et al. Quantification of MYCN, DDX1, and NAG gene copy number in neuroblastoma using a real-time quantitative PCR assay. Mod. Pathol. 2002, 15, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Ding, J.; Zhang, C.; Jia, J.; Weng, H.; Liu, W.; Zhang, D. Estimating the copy number of transgenes in transformed rice by real-time quantitative PCR. Plant Cell Rep. 2005, 23, 759–763. [Google Scholar] [CrossRef]
- Pös, O.; Radvanszky, J.; Styk, J.; Pös, Z.; Buglyó, G.; Kajsik, M.; Budis, J.; Nagy, B.; Szemes, T. Copy number variation: Methods and clinical applications. Appl. Sci. 2021, 11, 819. [Google Scholar] [CrossRef]
- Kalendar, R.; Raskina, O.; Belyayev, A.; Schulman, A.H. Long tandem arrays of Cassandra retroelements and their role in genome dynamics in plants. Int. J. Mol. Sci. 2020, 21, 2931. [Google Scholar] [CrossRef]
- Baruch, O.; Kashkush, K. Analysis of copy-number variation, insertional polymorphism, and methylation status of the Tiniest Class I (TRIM) and Class II (MITE) transposable element families in various rice strains. Plant Cell Rep. 2012, 31, 885–893. [Google Scholar] [CrossRef]
- Shams, I.; Raskina, O. Intraspecific and intraorganismal copy number dynamics of retrotransposons and tandem repeat in Aegilops speltoides Tausch (Poaceae, Triticeae). Protoplasma 2018, 255, 1023–1038. [Google Scholar] [CrossRef]
- Divashuk, M.G.; Karlov, G.I.; Kroupin, P.Y. Copy number variation of transposable elements in Thinopyrum intermedium and its diploid relative species. Plants 2019, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Kraitshtein, Z.; Yaakov, B.; Khasdan, V.; Kashkush, K. Genetic and epigenetic dynamics of a retrotransposon after allopolyploidization of wheat. Genetics 2010, 186, 801–812. [Google Scholar] [CrossRef] [Green Version]
- Yaakov, B.; Kashkush, K. Mobilization of Stowaway-like MITEs in newly formed allohexaploid wheat species. Plant Mol. Biol. 2012, 80, 419–427. [Google Scholar] [CrossRef]
- Pereira, C.M.; Stoffel, T.J.R.; Callegari-Jacques, S.M.; Hua-Van, A.; Capy, P.; Loreto, E.L.S. The somatic mobilization of transposable element mariner-Mos1 during the Drosophila lifespan and its biological consequences. Gene 2018, 679, 65–72. [Google Scholar] [CrossRef]
- Caballero-López, V.; Lundberg, M.; Sokolovskis, K.; Bensch, S. Transposable elements mark a repeat-rich region associated with migratory phenotypes of willow warblers (Phylloscopus trochilus). Mol. Ecol. 2022, 31, 1128–1141. [Google Scholar] [CrossRef]
- Coan, R.; Martins, C. Landscape of transposable elements focusing on the B chromosome of the cichlid fish Astatotilapia latifasciata. Genes 2018, 9, 269. [Google Scholar] [CrossRef]
- Papaceit, M.; Ávila, V.; Aguadé, M.; García-Dorado, A. The Dynamics of the Roo transposable element in mutation-accumulation lines and segregating populations of Drosophila melanogaster. Genetics 2007, 177, 511–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiel, E.E.; Cabrero, J.; Camacho, J.P.M.; López-León, M. Gypsy, RTE and Mariner transposable elements populate Eyprepocnemis plorans genome. Genetica 2012, 140, 365–374. [Google Scholar] [CrossRef]
- Divashuk, M.G.; Khuat, T.M.L.; Kroupin, P.Y.; Kirov, I.V.; Romanov, D.V.; Kiseleva, A.V.; Khrustaleva, L.I.; Alexeev, D.G.; Zelenin, A.S.; Klimushina, M.V.; et al. Variation in copy number of Ty3/Gypsy centromeric retrotransposons in the genomes of Thinopyrum intermedium and its diploid progenitors. PLoS ONE 2016, 11, e0154241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suntronpong, A.; Singchat, W.; Kruasuwan, W.; Prakhongcheep, O.; Sillapaprayoon, S.; Muangmai, N.; Somyong, S.; Indananda, C.; Kraichak, E.; Peyachoknagul, S.; et al. Characterization of centromeric satellite DNAs (MALREP) in the asian swamp eel (Monopterus albus) suggests the possible origin of repeats from transposable elements. Genomics 2020, 112, 3097–3107. [Google Scholar] [CrossRef] [PubMed]
- Kroupin, P.; Kuznetsova, V.; Romanov, D.; Kocheshkova, A.; Karlov, G.; Dang, T.X.; Khuat, T.M.L.; Kirov, I.; Alexandrov, O.; Polkhovskiy, A.; et al. Pipeline for the rapid development of cytogenetic markers using genomic data of related species. Genes 2019, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Kato, A. High-Density Fluorescence in situ hybridization signal detection on barley (Hordeum vulgare L.) chromosomes with improved probe screening and reprobing procedures. Genome 2011, 54, 151–159. [Google Scholar] [CrossRef]
- Baum, B.R.; Edwards, T.; Johnson, D.A. What does the Nr5S DNA multigene family tell us about the genomic relationship between Dasypyrum breviaristatum and D. villosum (Triticeae: Poaceae)? Mol. Genet. Genomics 2014, 289, 553–565. [Google Scholar] [CrossRef]
- Nikitina, E.; Kuznetsova, V.; Kroupin, P.; Karlov, G.I.; Divashuk, M.G. Development of specific Thinopyrum cytogenetic markers for wheat-wheatgrass hybrids using sequencing and qPCR Data. Int. J. Mol. Sci. 2020, 21, 4495. [Google Scholar] [CrossRef]
- Pestsova, E.G.; Goncharov, N.P.; Salina, E.A. Elimination of a tandem repeat of telomeric heterochromatin during the evolution of wheat. Theor. Appl. Genet. 1998, 97, 1380–1386. [Google Scholar] [CrossRef]
- Kroupin, P.Y.; Badaeva, E.D.; Sokolova, V.M.; Chikida, N.N.; Belousova, M.K.; Surzhikov, S.A.; Nikitina, E.A.; Kocheshkova, A.A.; Ulyanov, D.S.; Ermolaev, A.S.; et al. Aegilops crassa Boiss. repeatome characterized using low-coverage ngs as a source of new FISH markers: Application in phylogenetic studies of the Triticeae. Front. Plant Sci. 2022, 13, 980764. [Google Scholar] [CrossRef]
- Mahelka, V.; Kopecký, D.; Paštová, L. On the Genome constitution and evolution of intermediate wheatgrass (Thinopyrum intermedium: Poaceae, Triticeae). BMC Evol. Biol. 2011, 11, 127. [Google Scholar] [CrossRef] [Green Version]
- Kishii, M.; Wang, R.R.-C.; Tsujimoto, H. GISH Analysis revealed new aspect of genomic constitution of Thinopyrum intermedium. Czech J. Genet. Plant Breed. 2005, 41, 92–95. [Google Scholar] [CrossRef] [Green Version]
- Puterova, J.; Razumova, O.; Martinek, T.; Alexandrov, O.; Divashuk, M.; Kubat, Z.; Hobza, R.; Karlov, G.; Kejnovsky, E. Satellite DNA and transposable elements in seabuckthorn (Hippophae rhamnoides), a dioecious plant with small Y and large X chromosomes. Genome Biol. Evol. 2017, 9, 197–212. [Google Scholar] [CrossRef] [Green Version]
- Shams, I.; Raskina, O. Supernumerary B Chromosomes and plant genome changes: A snapshot of wild populations of Aegilops speltoides Tausch (Poaceae, Triticeae). Int. J. Mol. Sci. 2020, 21, 3768. [Google Scholar] [CrossRef]
- Wang, G.; He, Q.; Macas, J.; Novák, P.; Neumann, P.; Meng, D.; Zhao, H.; Guo, N.; Han, S.; Zong, M.; et al. Karyotypes and distribution of tandem repeat sequences in Brassica nigra determined by fluorescence in situ hybridization. Cytogenet. Genome Res. 2017, 152, 158–165. [Google Scholar] [CrossRef]
- Tang, Z.X.; Yang, Z.J.; Fu, S.L.; Yang, M.Y.; Li, G.R.; Zhang, H.Q.; Tan, F.Q.; Ren, Z. A New long terminal repeat (LTR) sequence allows to identify J genome from JS and St genomes of Thinopyrum Intermedium. J. Appl. Genet. 2011, 52, 31–33. [Google Scholar] [CrossRef] [Green Version]
- Yaakov, B.; Meyer, K.; Ben-David, S.; Kashkush, K. Copy number variation of transposable elements in Triticum–Aegilops genus suggests evolutionary and revolutionary dynamics following allopolyploidization. Plant Cell Rep. 2013, 32, 1615–1624. [Google Scholar] [CrossRef]
- Belyayev, A.; Kalendar, R.; Brodsky, L.; Nevo, E.; Schulman, A.H.; Raskina, O. Transposable elements in a marginal plant population: Temporal fluctuations provide new insights into genome evolution of wild diploid wheat. Mob. DNA 2010, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Rogers, S.O.; Bendich, A.J. Extraction of total cellular DNA from plants, algae and fungi. In Plant Molecular Biology Manual; Springer: Dordrecht, The Netherlands, 1994; pp. 183–190. [Google Scholar]
- Doyle, J.J. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Novák, P.; Ávila Robledillo, L.; Koblížková, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Kroupin, P.Y.; Divashuk, M.G.; Belov, V.I.; Glukhova, L.I.; Aleksandrov, O.S.; Karlov, G.I. Comparative molecular cytogenetic characterization of partial wheat-wheatgrass hybrids. Russ. J. Genet. 2011, 47, 432–437. [Google Scholar] [CrossRef]
- Badaeva, E.D.; Ruban, A.S.; Aliyeva-Schnorr, L.; Municio, C.; Hesse, S.; Houben, A. In situ hybridization to plant chromosomes. In Fluorescence In Situ hybridization (FISH); Springer: Berlin/Heidelberg, Germany, 2017; pp. 477–494. [Google Scholar]
- Kirov, I.; Divashuk, M.; Van Laere, K.; Soloviev, A.; Khrustaleva, L. An easy “SteamDrop” method for high quality plant chromosome preparation. Mol. Cytogenet. 2014, 7, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuznetsova, V.M.; Razumova, O.V.; Karlov, G.I.; Dang, T.X.; Kroupin, P.Y.; Divashuk, M.G. Some peculiarities in application of denaturating and non-denaturating in situ hybridization on chromosomes of cereals. Moscow Univ. Biol. Sci. Bull. 2019, 74, 75–80. [Google Scholar] [CrossRef]
- Alexandrov, O.S.; Karlov, G.I. Molecular cytogenetic analysis and genomic organization of major DNA repeats in castor bean (Ricinus communis L.). Mol. Genet. Genomics 2016, 291, 775–787. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession | Purpose (qPCR/FISH/Sequencing) | Case (Paragraph) |
|---|---|---|---|
| Ae. crassa | K-2485 | qPCR, FISH | CL239 |
| Ae. crassa | AE 742 | qPCR, FISH, sequencing | CL244, CL131, CL239 |
| Ae. crassa | AE 1649 | qPCR, FISH | CL239 |
| Ae. tauschii | K-112 | qPCR, FISH, sequencing | CL244, CL131, CL239 |
| Ae. tauschii | Clae 3 | qPCR | P720 |
| D. breviaristatum | PI 516547 | qPCR, FISH | pHv-961, 19-202(a) |
| D. villosum | PI 21717 | qPCR, FISH | pHv-961 |
| H. rhamnoides | Botanicheskaya lyubitelskaya | FISH, sequencing | HRTR12 |
| H. rhamnoides | Darlovo 7 (Male) | qPCR | HRTR12 |
| H. rhamnoides | Darlovo 10 (Female) | qPCR | HRTR12 |
| H. rhamnoides | Krasnoyarsk 3 (Female) | qPCR | HRTR12 |
| H. rhamnoides | Moshkovo 2 (Male) | qPCR | HRTR12 |
| H. rhamnoides | Dolzhanskaya 11 (Male) | qPCR | HRTR12 |
| H. rhamnoides | Dolzhanskaya 18 (Female) | qPCR | HRTR12 |
| H. rhamnoides | Pollinator 1 | FISH, sequencing | HRTR12 |
| S. cereale | EM1 | qPCR | P720 |
| T. aestivum | Chinese Spring | qPCR, FISH | CL244 |
| T. aestivum | Ivolga | qPCR | P720 |
| T. aestivum | Chinese Spring | qPCR, FISH | CL131, CL239 |
| T. aestivum | Nemchinovskaya 56 | qPCR | 17-202, 17-62 |
| Th. bessarabicum | PI 201890 | qPCR, FISH, sequencing | CL244, CL239 |
| Th. intermedium | PI 401200 | qPCR, FISH | 19-202(a), 17-202, 17-62 |
| Th. ponticum | PI 693508 | qPCR, FISH | 19-202(b) |
| Th. ponticum | 1158A/19 | qPCR, FISH, sequencing | 19-202(b) |
| Th. ponticum | PI 547313 | qPCR, FISH | 19-202(b) |
| Th. ponticum | PI 636523 | qPCR, FISH, sequencing | 19-202(a), 19-202(b), 17-202, 17-62 |
| Th. sartorii | PI 531745 | qPCR, FISH | 19-202(a) |
| Triticosecale | Solovey Kharkovskiy | FISH | P720 |
| WWGH | ZP26 | qPCR, FISH | 17-62 |
| WWGH | 166 | qPCR, FISH | 17-62 |
| WWGH | 548 | qPCR, FISH | 17-202, 17-62 |
| WWGH | 4044 | qPCR, FISH | 17-62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Divashuk, M.G.; Nikitina, E.A.; Sokolova, V.M.; Yurkina, A.I.; Kocheshkova, A.A.; Razumova, O.V.; Karlov, G.I.; Kroupin, P.Y. qPCR as a Selective Tool for Cytogenetics. Plants 2023, 12, 80. https://doi.org/10.3390/plants12010080
Divashuk MG, Nikitina EA, Sokolova VM, Yurkina AI, Kocheshkova AA, Razumova OV, Karlov GI, Kroupin PY. qPCR as a Selective Tool for Cytogenetics. Plants. 2023; 12(1):80. https://doi.org/10.3390/plants12010080
Chicago/Turabian StyleDivashuk, Mikhail G., Ekaterina A. Nikitina, Victoria M. Sokolova, Anna I. Yurkina, Alina A. Kocheshkova, Olga V. Razumova, Gennady I. Karlov, and Pavel Yu. Kroupin. 2023. "qPCR as a Selective Tool for Cytogenetics" Plants 12, no. 1: 80. https://doi.org/10.3390/plants12010080