
Pyramiding of Four Broad Spectrum Bacterial Blight Resistance Genes in Cross Breeds of Basmati Rice
 , ,
, ,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
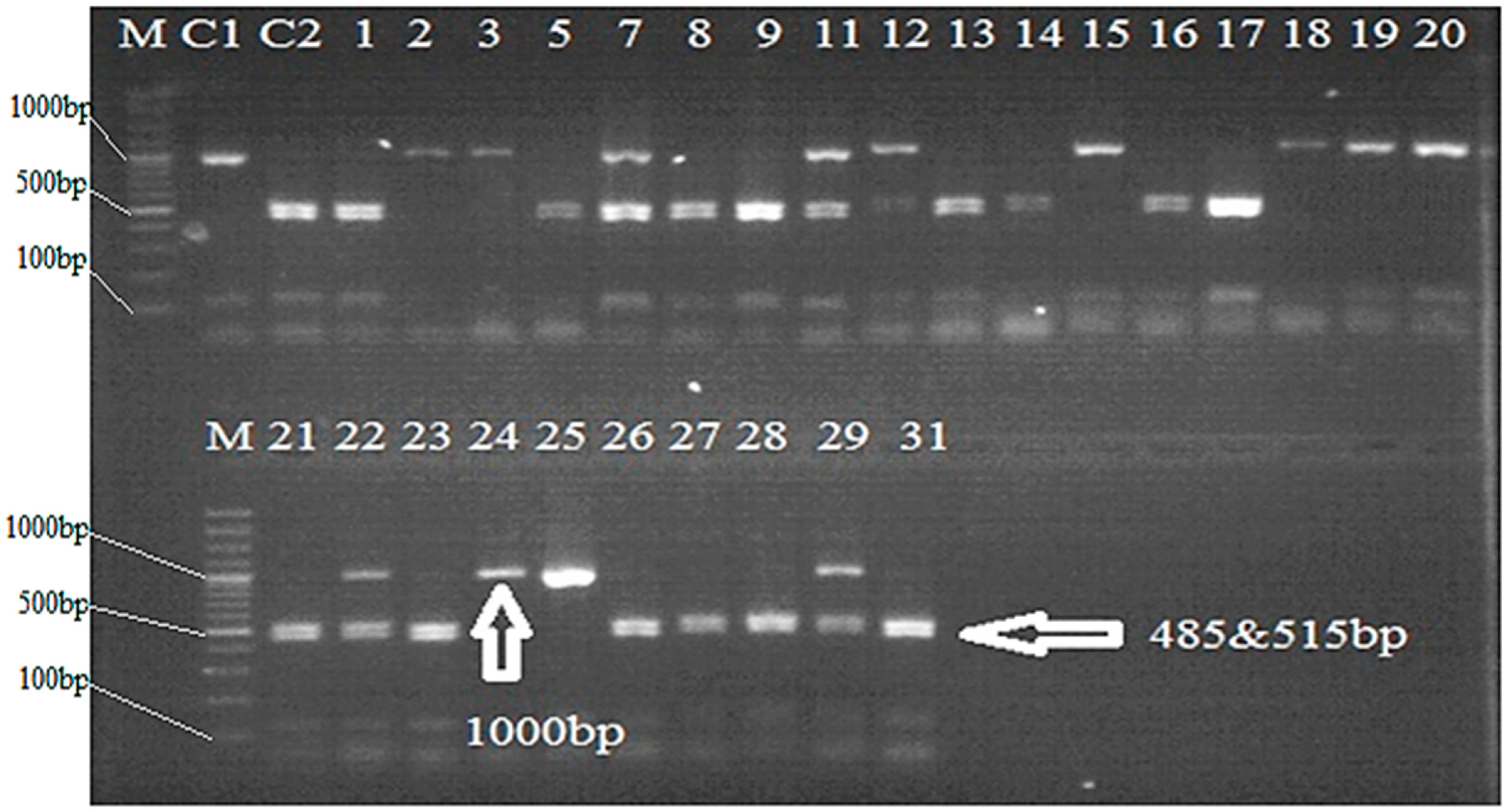
2.1. PCR Analysis
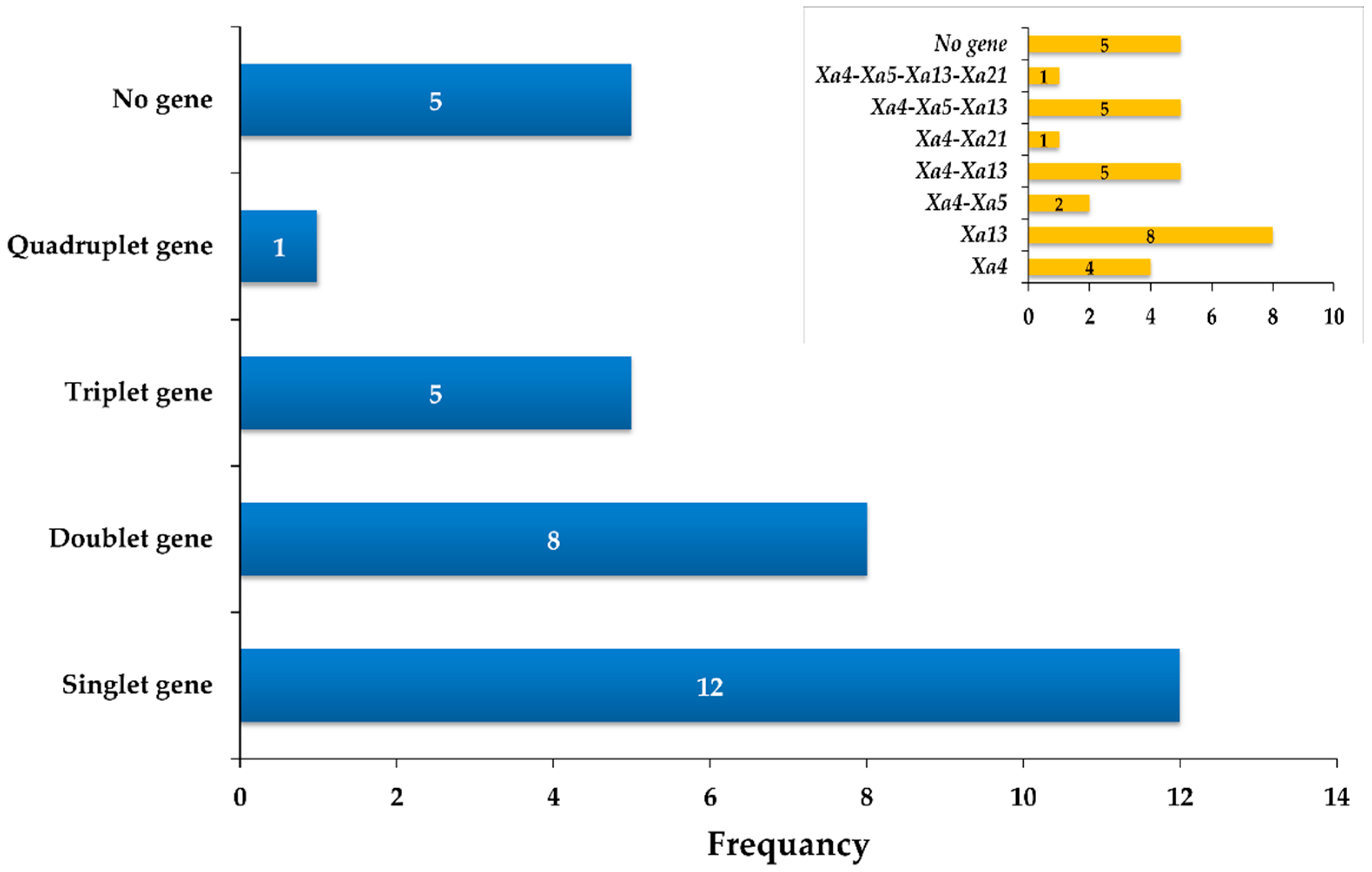
2.2. Identification of Pyramided Lines in BC1F6 Population
2.3. Responses of RILs to BB Isolates
3. Discussion
4. Materials and Methods
4.1. Pyramiding BB-R Genes into Basmati-385
4.2. Extraction of Genomic DNA
4.3. PCR Amplification of Bacterial Blight R Genes
4.4. Xanthomonas oryzae pv. oryzae (Xoo) Strain Isolation
4.5. Preparation of Inoculum from Pure Cultures
4.6. Data Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linares, O.F. African rice (Oryza glaberrima): History and future potential. Proc. Natl. Acad. Sci. USA 2002, 99, 16360–16365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noreen, R.; Khan, S.; Rabbani, A.; Kanwal, A.; Uzair, B. Screening of different rice (Oryza sativa L.) varieties for genetic diversity and bacterial blight R gene. Pak. J. Bot. 2020, 52, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- USDA. World Agricultural Production. Foreign Agricultural Service, Office of Global Analysis. United States Department of Agriculture; 2020. Available online: https://www.fas.usda.gov/data/world-agricultural-production (accessed on 18 November 2022).
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Gnanamanickam, S.S. Rice and its importance to human life. In Biological Control of Rice Diseases; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–11. [Google Scholar]
- Gul, A.; Xiumin, W.; Chandio, A.A.; Rehman, A.; Siyal, S.A.; Asare, I. Tracking the effect of climatic and non-climatic elements on rice production in Pakistan using the ARDL approach. Environ. Sci. Pollut. Res. 2022, 29, 31886–31900. [Google Scholar] [CrossRef] [PubMed]
- Woolfe, M.; Steele, K. The Authenticity of Basmati Rice—A Case Study. In DNA Techniques to Verify Food Authenticity; RSC Publishing: Cambridge, UK, 2019; pp. 207–218. [Google Scholar]
- Ahmad, M.; Akhtar, M.; Anwar, M. Basmati rice: Progress, issues and prospects for Pakistan. In Proceedings of the International Seminar in Rice Crop, Lahore, Pakistan, 2–3 October 2005; pp. 1–7. [Google Scholar]
- Bashir, K.; Khan, N.M.; Rasheed, S.; Salim, M. Indica rice varietal development in Pakistan: An overview. Water Environ. 2007, 5, 73–81. [Google Scholar] [CrossRef]
- Akram, M. Aromatic rices of Pakistan—A review. Pak. J. Agric. Res. 2009, 22, 154–160. [Google Scholar]
- Saleem, M.Y.; Mirza, J.I.; Haq, M.A. Combining ability analysis of some morpho-physiological traits in Basmati rice. Pak. J. Bot. 2010, 42, 3113–3123. [Google Scholar]
- Imran, M.; Masud, T.; Sakandar, H.A. Studies on physiochemical dimensions of two basmati (Super Basmati, Basmati 385) and two non-basmati (KS-282, IR-6) Pakistani commercial rice varieties. J. Glob. Innov. Agric. Soc. Sci. 2014, 2, 120–122. [Google Scholar] [CrossRef]
- Swings, J.; Van den Mooter, M.; Vauterin, L.; Hoste, B.; Gillis, M.; Mew, T.; Kersters, K. Reclassification of the Causal Agents of Bacterial Blight (Xanthomonas campestris pv. oryzae) and Bacterial Leaf Streak (Xanthomonas campestris pv. oryzicola) of Rice as Pathovars of Xanthomonas oryzae (ex Ishiyama 1922) sp. nov., nom. rev. Int. J. Syst. Evol. Microbiol. 1990, 40, 309–311. [Google Scholar] [CrossRef] [Green Version]
- Yasmin, S.; Hafeez, F.Y.; Mirza, M.S.; Rasul, M.; Arshad, H.M.; Zubair, M.; Iqbal, M. Biocontrol of bacterial leaf blight of rice and profiling of secondary metabolites produced by rhizospheric Pseudomonas aeruginosa BRp3. Front. Microbiol. 2017, 8, 1895. [Google Scholar] [CrossRef] [Green Version]
- Chukwu, S.; Rafii, M.; Ramlee, S.; Ismail, S.; Hasan, M.; Oladosu, Y.; Magaji, U.; Akos, I.; Olalekan, K.K. Bacterial leaf blight resistance in rice: A review of conventional breeding to molecular approach. Mol. Biol. Rep. 2019, 46, 1519–1532. [Google Scholar] [CrossRef] [PubMed]
- Waheed, M.; Inamullah, A.H.; Sirajuddin, A.H.; Khan, A.; Khan, A. Evaluation of rice genotypes for resistance against bacterial leaf blight. Pak. J. Bot. 2009, 41, 329–335. [Google Scholar]
- Ahsan, R.; Ullah, S.; Yaseen, I.; Fateh, F.S.; Fayyaz, M.; Asad, S.; Jamal, A.; Sufyan, M.; Zakria, M. Assessment of bacterial leaf blight incidence and severity in rice growing areas of Pakistan. Pak. J. Agric. Res. 2021, 34, 693–699. [Google Scholar] [CrossRef]
- Arif, M.; Chilvers, G.; Day, S.; Naveed, S.; Woolfe, M.; Rodionova, O.Y.; Pomerantsev, A.; Kracht, O.; Brodie, C.; Mihailova, A.; et al. Differentiating Pakistani long-grain rice grown inside and outside the accepted Basmati Himalayan geographical region using a ‘one-class’ multi-element chemometric model. Food Control 2021, 123, 107827. [Google Scholar] [CrossRef]
- Islam, W.J. Management of plant virus diseases; farmer’s knowledge and our suggestions. Host Viruses 2017, 4, 28. [Google Scholar]
- Ali, H.; Abbasi, F.M.; Ahmad, H. Bacterial Blight, a serious threat to productivity of rice (Oryza Sativa L.), an overview. Int. J. Biosci. 2016, 9, 154–169. [Google Scholar]
- Noman, A.; Bashir, R.; Aqeel, M.; Anwer, S.; Iftikhar, W.; Zainab, M.; Zafar, S.; Khan, S.; Islam, W.; Adnan, M.; et al. Success of transgenic cotton (Gossypium hirsutum L.): Fiction or reality? Cogent Food Agric. 2016, 2, 1207844. [Google Scholar] [CrossRef]
- Kim, S.-M. Identification of novel recessive gene xa44 (t) conferring resistance to bacterial blight races in rice by QTL linkage analysis using an SNP chip. Theor. Appl. Genet. 2018, 131, 2733–2743. [Google Scholar] [CrossRef] [Green Version]
- Neelam, K.; Mahajan, R.; Gupta, V.; Bhatia, D.; Gill, B.K.; Komal, R.; Lore, J.S.; Mangat, G.S.; Singh, K. High-resolution genetic mapping of a novel bacterial blight resistance gene xa-45 (t) identified from Oryza glaberrima and transferred to Oryza sativa. Theor. Appl. Genet. 2020, 133, 689–705. [Google Scholar] [CrossRef]
- Pradhan, S.; Barik, S.; Nayak, D.; Pradhan, A.; Pandit, E.; Nayak, P.; Das, S.; Pathak, H. Genetics, molecular mechanisms and deployment of bacterial blight resistance genes in rice. Crit. Rev. Plant Sci. 2020, 39, 360–385. [Google Scholar] [CrossRef]
- Kim, S.-M.; Suh, J.-P.; Qin, Y.; Noh, T.-H.; Reinke, R.F.; Jena, K.K. Identification and fine-mapping of a new resistance gene, Xa40, conferring resistance to bacterial blight races in rice (Oryza sativa L.). Theor. Appl. Genet. 2015, 128, 1933–1943. [Google Scholar] [CrossRef]
- Bhasin, H.; Bhatia, D.; Raghuvanshi, S.; Lore, J.S.; Sahi, G.K.; Kaur, B.; Vikal, Y.; Singh, K. New PCR-based sequence-tagged site marker for bacterial blight resistance gene Xa38 of rice. Mol. Breed. 2012, 30, 607–611. [Google Scholar] [CrossRef]
- Kumar, P.N.; Sujatha, K.; Laha, G.; Rao, K.S.; Mishra, B.; Viraktamath, B.; Hari, Y.; Reddy, C.; Balachandran, S.; Ram, T. Identification and fine-mapping of Xa33, a novel gene for resistance to Xanthomonas oryzae pv. oryzae. Phytopathology 2012, 102, 222–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Liu, P.; Mei, L.; He, X.; Chen, L.; Liu, H.; Shen, S.; Ji, Z.; Zheng, X.; Zhang, Y. Xa7, a new executor R gene that confers durable and broad-spectrum resistance to bacterial blight disease in rice. Plant Commun. 2021, 2, 100143. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-Y.; Pi, L.-Y.; Wang, G.-L.; Gardner, J.; Holsten, T.; Ronald, P.C. Evolution of the rice Xa21 disease resistance gene family. Plant Cell 1997, 9, 1279–1287. [Google Scholar] [PubMed]
- Yang, D.; Sanchez, A.; Khush, G.; Zhu, Y.; Huang, N. Construction of a BAC contig containing the xa5 locus in rice. Theor. Appl. Genet. 1998, 97, 1120–1124. [Google Scholar] [CrossRef]
- Sun, X.; Yang, Z.; Wang, S.; Zhang, Q. Identification of a 47-kb DNA fragment containing Xa4, a locus for bacterial blight resistance in rice. Theor. Appl. Genet. 2003, 106, 683–687. [Google Scholar] [CrossRef]
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yang, F.; Chu, Z.; Wang, G.-L.; White, F.F. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef]
- Niño-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef]
- Cheema, K.K.; Grewal, N.K.; Vikal, Y.; Sharma, R.; Lore, J.S.; Das, A.; Bhatia, D.; Mahajan, R.; Gupta, V.; Bharaj, T.S. A novel bacterial blight resistance gene from Oryza nivara mapped to 38 kb region on chromosome 4L and transferred to Oryza sativa L. Genet. Res. 2008, 90, 397–407. [Google Scholar] [CrossRef]
- Zhang, F.; Zhuo, D.L.; Zhang, F.; Huang, L.Y.; Wang, W.S.; Xu, J.L.; vera Cruz, C.; Li, Z.K.; Zhou, Y.L. Xa39, a novel dominant gene conferring broad-spectrum resistance to Xanthomonas oryzae pv. oryzae in rice. Plant Pathol. 2015, 64, 568–575. [Google Scholar] [CrossRef]
- Sabar, M.; Akhter, M.; Bibi, T.; Riaz, A.; Haider, Z.; Khan, A.R.; Bibi, A. Basmati rice lines development carrying multiple bacterial blight resistance genes pyramided using the marker-assisted backcross breeding approach. Mol. Breed. 2019, 39, 155. [Google Scholar] [CrossRef]
- Nayak, D.; Pandit, E.; Mohanty, S.; Barik, D.; Pradhan, S.K. Marker-assisted selection in back cross progenies for transfer of bacterial leaf blight resistance genes into a popular lowland rice cultivar. ORYZA Int. J. Rice 2015, 52, 163–172. [Google Scholar]
- Hsu, Y.-C.; Chiu, C.-H.; Yap, R.; Tseng, Y.-C.; Wu, Y.-P. Pyramiding bacterial blight resistance genes in Tainung82 for broad-spectrum resistance using marker-assisted selection. Int. J. Mol. Sci. 2020, 21, 1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, I.; Ali, H.; Islam, M.; Ullah, W.; Haris, M.; Khan, M.Q.; Shafiq-ur-Rehman, K.K.; Khan, K.; Ghani, B. Molecular analysis of bacterial blight resistance gene XA7 in advance population of rice using STS markers. Int. J. Biosci. 2020, 17, 1–10. [Google Scholar]
- Yugander, A.; Sundaram, R.; Ladhalakshmi, D.; Hajira, S.; Prakasam, V.; Prasad, M.; Sheshu Madhav, M.; Ravindra Babu, V.; Laha, G.S. Virulence profiling of Xanthomonas oryzae pv. oryzae isolates, causing bacterial blight of rice in India. Eur. J. Plant Pathol. 2017, 149, 171–191. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Nayak, D.K.; Mohanty, S.; Behera, L.; Barik, S.R.; Pandit, E.; Lenka, S.; Anandan, A. Pyramiding of three bacterial blight resistance genes for broad-spectrum resistance in deepwater rice variety, Jalmagna. Rice 2015, 8, 19. [Google Scholar] [CrossRef]
- Swathi, G.; Durga Rani, C.V.; Md, J.; Madhav, M.S.; Vanisree, S.; Anuradha, C.; Kumar, N.R.; Kumar, N.; Kumari, K.A.; Bhogadhi, S.C.; et al. Marker-assisted introgression of the major bacterial blight resistance genes, Xa21 and xa13, and blast resistance gene, Pi54, into the popular rice variety, JGL1798. Mol. Breed. 2019, 39, 58. [Google Scholar] [CrossRef]
- Suh, J.-P.; Cho, Y.-C.; Won, Y.-J.; Ahn, E.-K.; Baek, M.-K.; Kim, M.-K.; Kim, B.-K.; Jena, K.K. Development of resistant gene-pyramided japonica rice for multiple biotic stresses using molecular marker-assisted selection. Plant Breed. Biotechnol. 2015, 3, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Das, G.; Rao, G.J.N. Molecular marker assisted gene stacking for biotic and abiotic stress resistance genes in an elite rice cultivar. Front. Plant Sci. 2015, 6, 698. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Sidhu, J.; Huang, N.; Vikal, Y.; Li, Z.; Brar, D.; Dhaliwal, H.; Khush, G.S. Pyramiding three bacterial blight resistance genes (xa5, xa13 and Xa21) using marker-assisted selection into indica rice cultivar PR106. Theor. Appl. Genet. 2001, 102, 1011–1015. [Google Scholar] [CrossRef]
- Sabar, M.; Bibi, T.; Farooq, H.U.; Haider, Z.; Naseem, I.; Mahmood, A.; Akhter, M. Molecular screening of rice (Oryza sativa L.) germplasm for Xa4, xa5 and Xa21 bacterial leaf blight (BLB) resistant genes using linked marker approach. Afr. J. Biotechnol. 2016, 15, 2317–2324. [Google Scholar]
- Dash, A.K.; Rao, R.N.; Rao, G.; Verma, R.L.; Katara, J.L.; Mukherjee, A.K.; Singh, O.N.; Bagchi, T.B. Phenotypic and marker-assisted genetic enhancement of parental lines of Rajalaxmi, an elite rice hybrid. Front. Plant Sci. 2016, 7, 1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eker, T.; Sari, D.; Sari, H.; Tosun, H.S.; Toker, C. A kabuli chickpea ideotype. Sci. Rep. 2022, 12, 1611. [Google Scholar] [CrossRef]
- Sundaram, R.; Laha, G.; Viraktamath, B.; Sujatha, K.; Natarajkumar, P.; Hari, Y.; Srinivasa Rao, K.; Reddy, C.; Balachandran, S.; Madhav, M.S.; et al. Marker assisted breeding for development of bacterial blight resistant rice. J. Plant Pathol. 2011, 500, 154–182. [Google Scholar]
- Dokku, P.; Das, K.; Rao, G.J.N. Pyramiding of four resistance genes of bacterial blight in Tapaswini, an elite rice cultivar, through marker-assisted selection. Euphytica 2013, 192, 87–96. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Baksh, S.Y.; Mukherjee, A.K.; Mohanty, S.P. Development of flash-flood tolerant and durable bacterial blight resistant versions of mega rice variety ‘Swarna’ through marker-assisted backcross breeding. Sci. Rep. 2019, 9, 12810. [Google Scholar] [CrossRef] [Green Version]
- Guvvala, L.D.; Koradi, P.; Shenoy, V.; Marella, L.S. Improvement of resistance to bacterial blight through marker assisted backcross breeding and field validation in rice (Oryza sativa). Res. J. Biol. 2013, 1, 52–66. [Google Scholar]
- Baliyan, N.; Malik, R.; Rani, R.; Mehta, K.; Vashisth, U.; Dhillon, S.; Boora, K.S. Integrating marker-assisted background analysis with foreground selection for pyramiding bacterial blight resistance genes into Basmati rice. Comptes Rendus Biol. 2018, 341, 1–8. [Google Scholar] [CrossRef]
- Suh, J.-P.; Jeung, J.-U.; Noh, T.-H.; Cho, Y.-C.; Park, S.-H.; Park, H.-S.; Shin, M.-S.; Kim, C.-K.; Jena, K.K. Development of breeding lines with three pyramided resistance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice 2013, 6, 5. [Google Scholar] [CrossRef]
- Rashid, M.M.; Nihad, S.A.I.; Khan, M.A.I.; Haque, A.; Ara, A.; Ferdous, T.; Hasan, M.A.I.; Latif, M.A. Pathotype profiling, distribution and virulence analysis of Xanthomonas oryzae pv. oryzae causing bacterial blight disease of rice in Bangladesh. J. Phytopathol. 2021, 169, 438–446. [Google Scholar] [CrossRef]
- Yugander, A.; Ershad, M.; Muthuraman, P.P.; Prakasam, V.; Ladhalakshmi, D.; Sheshu Madhav, M.; Srinivas Prasad, M.; Sundaram, R.M.; Laha, G.S. Understanding the variability of rice bacterial blight pathogen, Xanthomonas oryzae pv. oryzae in Andhra Pradesh, India. J. Basic Microbiol. 2022, 62, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-Q.; Angeles, E.; Abenes, M.; Khush, G.; Huang, N. RAPD and RFLP mapping of the bacterial blight resistance gene xa-13 in rice. Theor. Appl. Genet. 1996, 93, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.; Brar, D.; Huang, N.; Li, Z.; Khush, G.S. Sequence tagged site marker-assisted selection for three bacterial blight resistance genes in rice. Crop Sci. 2000, 40, 792–797. [Google Scholar] [CrossRef]
- Shanti, M.; Shenoy, V.; Devi, G.L.; Kumar, V.M.; Premalatha, P.; Kumar, G.N.; Shashidhar, H.; Zehr, U.; Freeman, W.H. Marker-assisted breeding for resistance to bacterial leaf blight in popular cultivar and parental lines of hybrid rice. J. Plant Pathol. 2010, 92, 495–501. [Google Scholar]
- Bojun, M.; Wenming, W.; Bin, Z.; Yongli, Z.; Lihuang, Z.; Wenxue, Z. Studies of PCR marker for the rice bacterial blight resistance gene Xa-4. Yi Chuan Hered. 1999, 21, 9–12. [Google Scholar]
- Yoshimura, S.; Yoshimura, A.; Iwata, N.; McCouch, S.R.; Abenes, M.L.; Baraoidan, M.R.; Mew, T.W.; Nelson, R.J. Tagging and combining bacterial blight resistance genes in rice using RAPD and RFLP markers. Mol. Breed. 1995, 1, 375–387. [Google Scholar] [CrossRef]
- Ronald, P.C.; Albano, B.; Tabien, R.; Abenes, L.; Wu, K.-S.; McCouch, S.; Tanksley, S.D. Genetic and physical analysis of the rice bacterial blight disease resistance locus, Xa21. Mol. Gen. Genet. 1992, 236, 113–120. [Google Scholar] [CrossRef]
- Kauffman, H.E. An improved technique for evaluat-ing resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 1973, 57, 537–541. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Xa-4 | xa-5 | xa-13 | Xa-21 |
|---|---|---|---|---|
| Line-1 | + | - | + | - |
| Line-2 | + | - | - | - |
| Line-3 | + | - | - | - |
| Line-4 | + | + | - | - |
| Line-5 | - | - | + | - |
| Line-6 | + | + | - | - |
| Line-7 | - | - | +/- | - |
| Line-8 | + | + | + | - |
| Line-9 | + | - | + | - |
| Line-10 | - | - | - | - |
| Line-11 | + | - | +/- | - |
| Line-12 | + | - | +/- | - |
| Line-13 | - | - | + | - |
| Line-14 | - | - | + | - |
| Line-15 | - | - | - | - |
| Line-16 | + | + | + | - |
| Line-17 | + | - | + | - |
| Line-18 | + | - | - | - |
| Line-19 | + | - | - | - |
| Line-20 | + | - | - | + |
| Line-21 | + | + | + | - |
| Line-22 | - | - | +/- | - |
| Line-23 | + | + | + | + |
| Line-24 | - | - | - | - |
| Line-25 | - | - | - | - |
| Line-26 | - | - | + | - |
| Line-27 | + | + | + | - |
| Line-28 | + | + | + | - |
| Line-29 | - | - | +/- | - |
| Line-30 | - | - | - | - |
| Line-31 | - | - | + | - |
| Basmati-385 | + | - | - | - |
| IRBB-59 | - | + | + | + |
| Genotypes | Lesion Length (%) | Reaction to Isolates | ||||
|---|---|---|---|---|---|---|
| Xoo1 | Xoo2 | Xoo3 | Xoo1 | Xoo2 | Xoo3 | |
| Line-1 | 1.3 J | 10.6 J–L | 8.5 K–M | HR | R | R |
| Line-2 | 15 G–J | 21.4 G–K | 18.8 G–L | MR | MR | MR |
| Line-3 | 13.6 G–J | 25.2 F–J | 19.6 G–L | MR | MR | MR |
| Line-4 | 18.6 F–I | 15.6 I–L | 29.9 D–G | MR | MR | MS |
| Line-5 | 9.1 H–J | 21.1 G–K | 31.7 C–G | R | MR | MS |
| Line-6 | 6 IJ | 8 KL | 12.8 I–M | R | R | R |
| Line-7 | 31.2 C–F | 50.4 A–C | 40.1 C–E | MS | MS | MS |
| Line-8 | 3.9 IJ | 3.4 L | 7.1 LM | R | R | R |
| Line-9 | 13.2 G–J | 11.9 J–L | 11.7 J–M | MR | R | R |
| Line-10 | 31.5 C–F | 30.6 E–I | 24.1 F–J | MS | MS | MR |
| Line-11 | 23.3 E–H | 12.5 J–L | 15.3 H–M | MR | R | MR |
| Line-12 | 35.8 B–E | 40.1 B–F | 56.8 AB | MS | MS | S |
| Line-13 | 34 B–F | 20.1 H–K | 34.3 C–F | MS | MR | MS |
| Line-14 | 22.9 E–H | 29.6 E–I | 43.7 B–D | MR | MS | MS |
| Line-15 | 59.8 A | 52.7 AB | 66.5 A | S | S | S |
| Line-16 | 3.3 IJ | 6 KL | 13.6 I–M | HR | R | MR |
| Line-17 | 12.8 G–J | 9.6 J–L | 12.4 J–M | R | R | R |
| Line-18 | 36.2 B–E | 40.9 B–F | 28.8 E–H | MS | MS | MS |
| Line-19 | 36.8 B–E | 36.1 C–G | 27.2 E–I | MS | MS | MS |
| Line-20 | 11.5 H–J | 12.7 J–L | 13.8 I–M | R | R | MR |
| Line-21 | 4 IJ | 7.4 KL | 9.1 K–M | R | R | R |
| Line-22 | 27.7 D–G | 39.1 B–F | 30.7 C–G | MS | MS | MS |
| Line-23 | 1.4 J | 2.3 L | 2.4 M | HR | HR | HR |
| Line-24 | 47.2 A–C | 42.4 A–E | 45.1 BC | MS | MS | MS |
| Line-25 | 37.8 B–E | 58 A | 44.9 BC | MS | S | MS |
| Line-26 | 44.5 A–C | 19.7 H–K | 22.8 F–K | MS | MR | MR |
| Line-27 | 2.1 J | 2.9 L | 8.6 K–M | HR | HR | R |
| Line-28 | 4.1 IJ | 1 L | 7.2 LM | R | HR | R |
| Line-29 | 60.1 A | 31.1 E–I | 40.5 C–E | S | MS | MS |
| Line-30 | 42.5 B–D | 32.1 D–H | 29.4 D–H | MS | MS | MS |
| Line-31 | 46.3 A–C | 34.5 D–H | 34.4 C–F | MS | MS | MS |
| Basmati-385 | 49.1 AB | 47.9 A–D | 56.7 AB | MS | MS | S |
| IRBB-59 | 7.3 H–J | 1.6 L | 2 M | R | HR | HR |
| Range | 60.1–1.3 | 58–1 | 66.5–2 | |||
| Grand mean | 24.1 | 23.7 | 25.8 | |||
| Primer | Primer Sequence (5′ to 3′) | Linked Gene | Reference |
|---|---|---|---|
| MP 1, 2 | (F) ATCGATCGATCTTCACGAGG | Xa4 | [60] |
| (R) TGCTATAAAAGGCATTCGGG | |||
| RZ207 | (F) GCCTCGAGCATCATCATCAG | xa5 | [61] |
| (R) ATCAACCTGCACTTGCCTGG | |||
| RG136 | (F) TCCCAGAAAGCTACTACAGC | xa13 | [57] |
| (R) GCAGACTCCAGTTTGACTTC | |||
| pTA248 | (F)AGACGCGGAAGGGTGGTTCC CGGA | Xa21 | [62] |
| (R) AGACCGGTAATCGAAAGATGAAA |
| Groups | Lesion Percentage | Disease Rating Scale |
|---|---|---|
| HR (Highly Resistant) | 0–3 | 1 |
| R (Resistant) | 4–12 | 3 |
| MR (Moderately Resistant) | 12–25 | 4 |
| MS (Moderately Susceptible) | 25–50 | 5 |
| S (Susceptible) | 51–87 | 7 |
| HS (Highly Susceptible) | 87–100 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, I.; Ali, H.; Mahmood, T.; Khan, M.N.; Haris, M.; Shah, H.; Mihoub, A.; Jamal, A.; Saeed, M.F.; Mancinelli, R.; et al. Pyramiding of Four Broad Spectrum Bacterial Blight Resistance Genes in Cross Breeds of Basmati Rice. Plants 2023, 12, 46. https://doi.org/10.3390/plants12010046
Ullah I, Ali H, Mahmood T, Khan MN, Haris M, Shah H, Mihoub A, Jamal A, Saeed MF, Mancinelli R, et al. Pyramiding of Four Broad Spectrum Bacterial Blight Resistance Genes in Cross Breeds of Basmati Rice. Plants. 2023; 12(1):46. https://doi.org/10.3390/plants12010046
Chicago/Turabian StyleUllah, Irfan, Hamid Ali, Tariq Mahmood, Mudassar Nawaz Khan, Muhammad Haris, Hussain Shah, Adil Mihoub, Aftab Jamal, Muhammad Farhan Saeed, Roberto Mancinelli, and et al. 2023. "Pyramiding of Four Broad Spectrum Bacterial Blight Resistance Genes in Cross Breeds of Basmati Rice" Plants 12, no. 1: 46. https://doi.org/10.3390/plants12010046