
Characterization of Neochloris oleoabundans under Different Cultivation Modes and First Results on Bioactivity of Its Extracts against HCoV-229E Virus
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
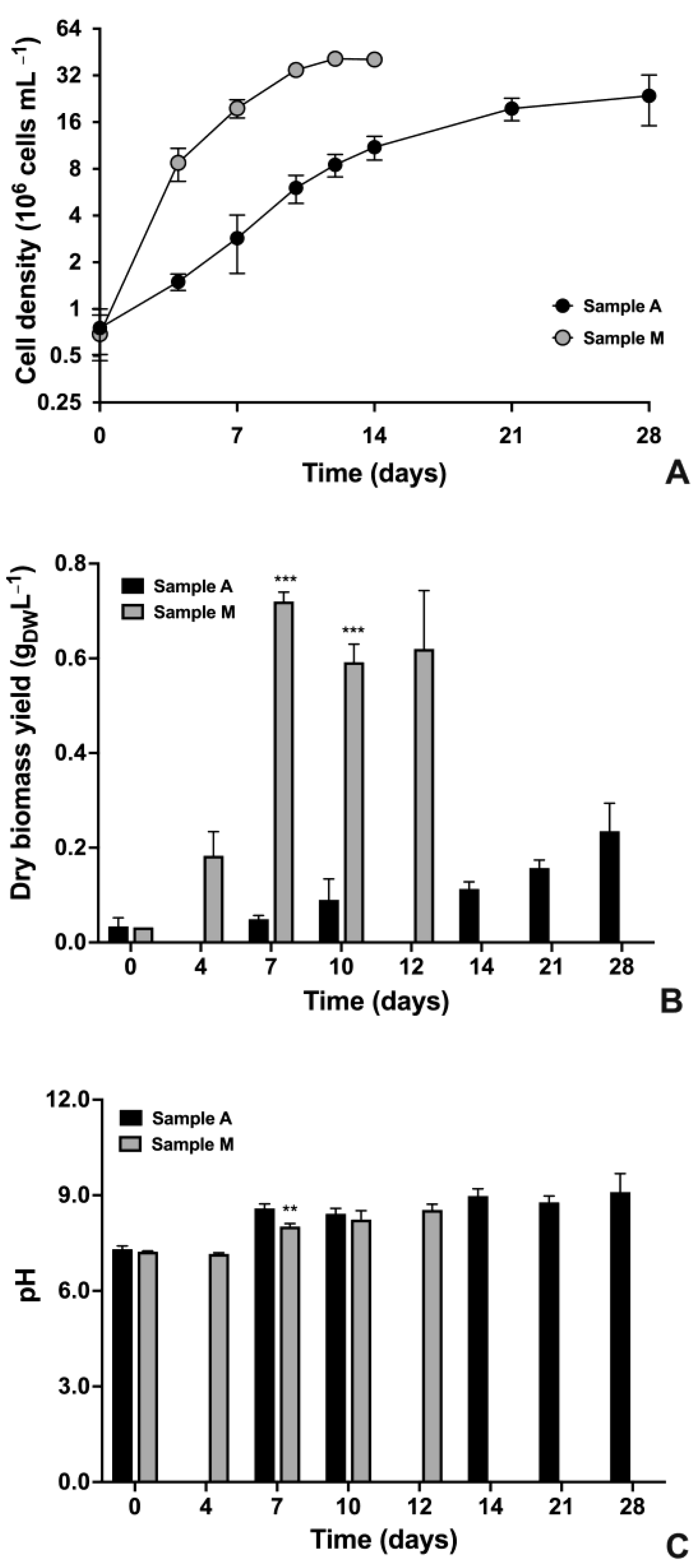
2.1. Growth Parameters
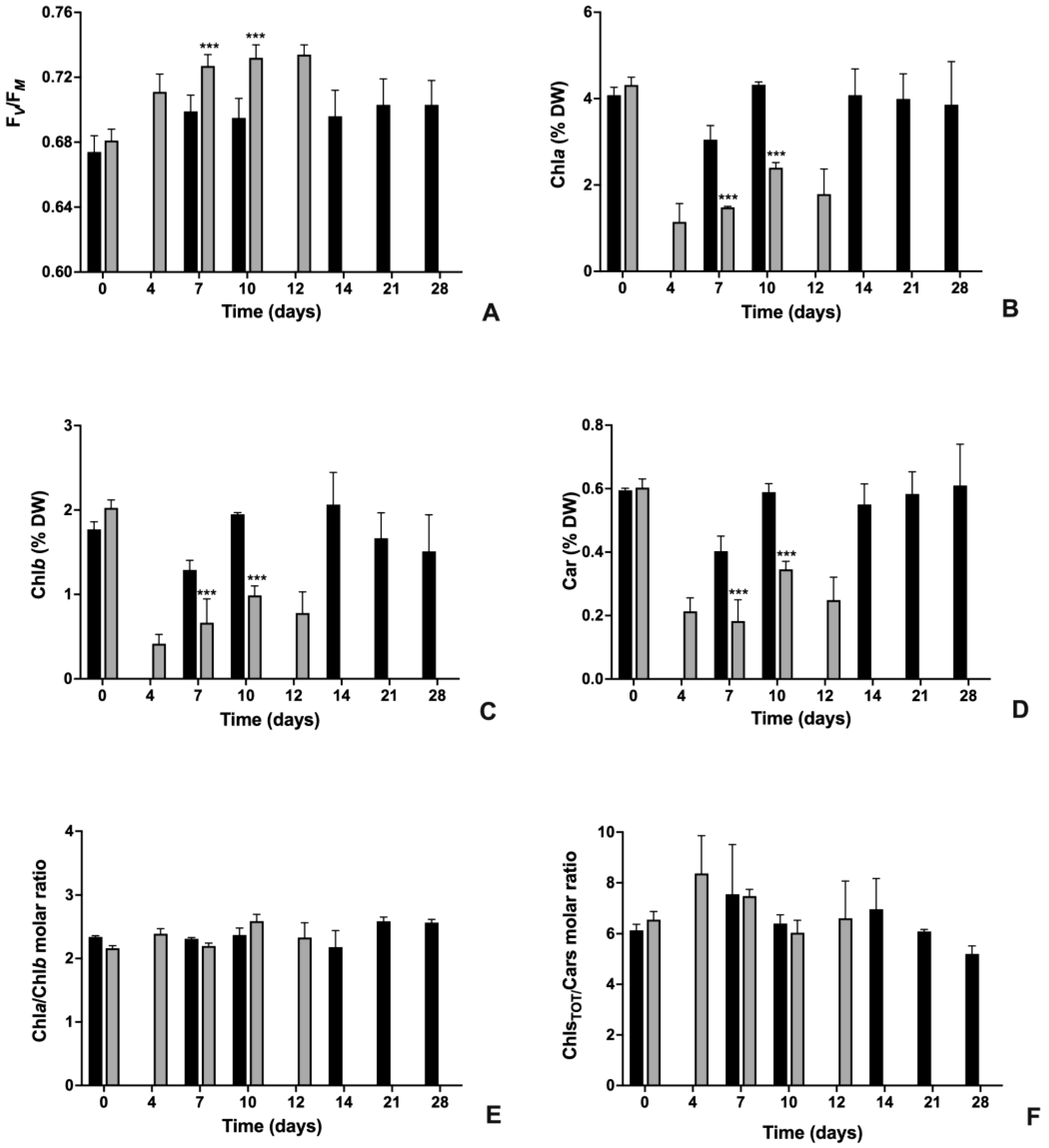
2.2. Photosynthetic Parameters and Chemical Composition of Algae
2.2.1. PSII Maximum Quantum Yield and Photosynthetic Pigment Content
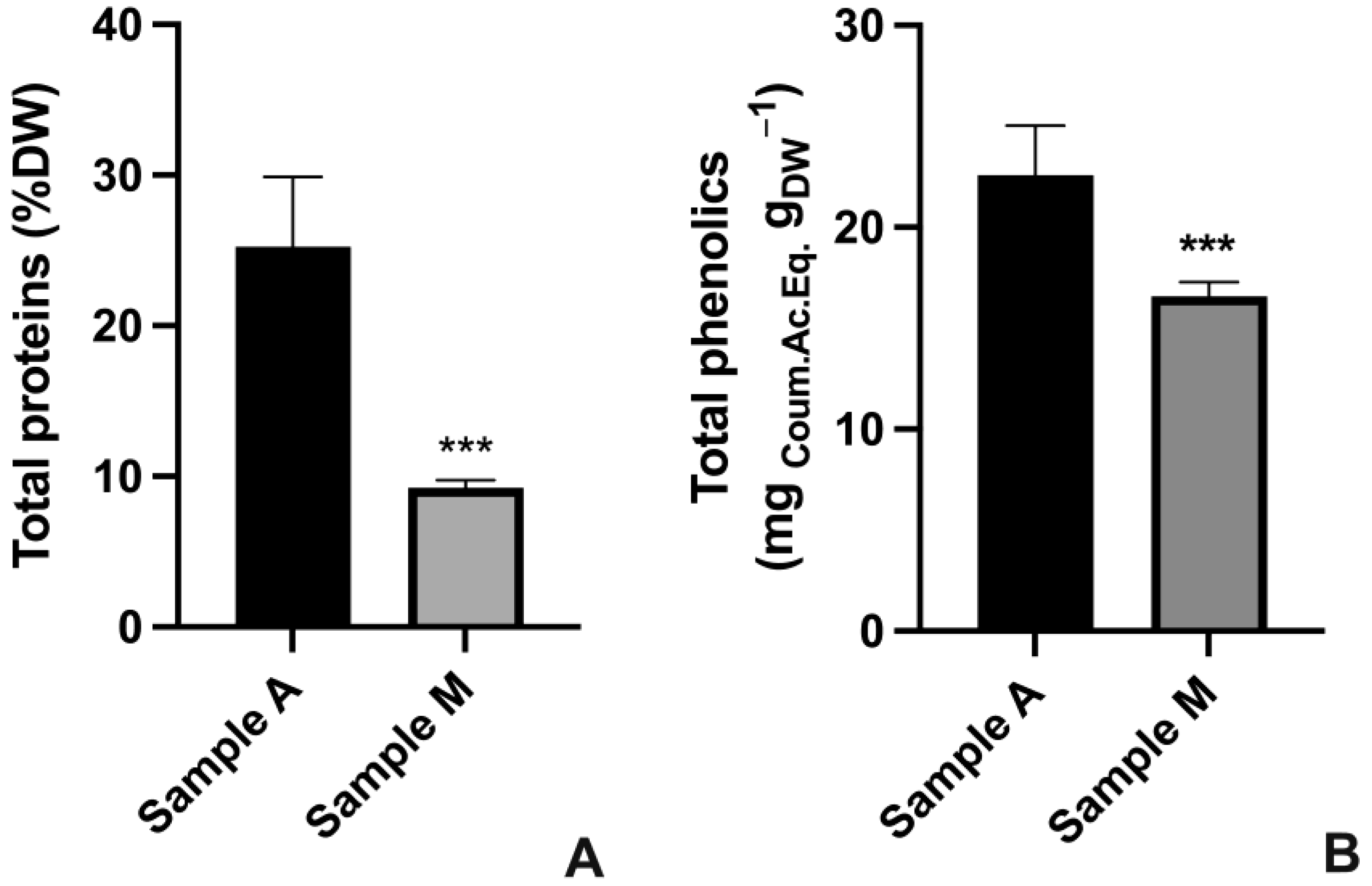
2.2.2. Quantification of Total Proteins and Total Phenolics
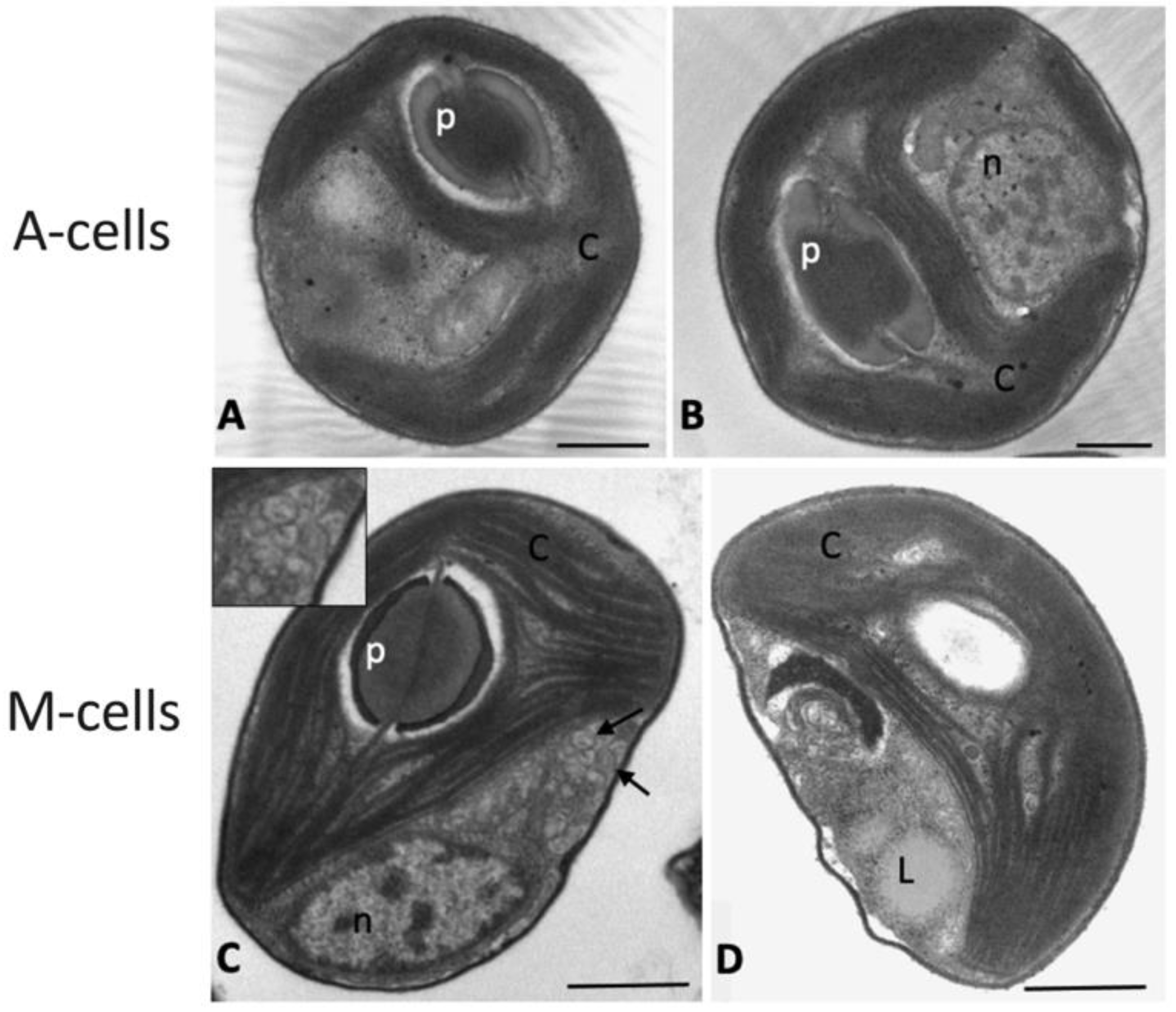
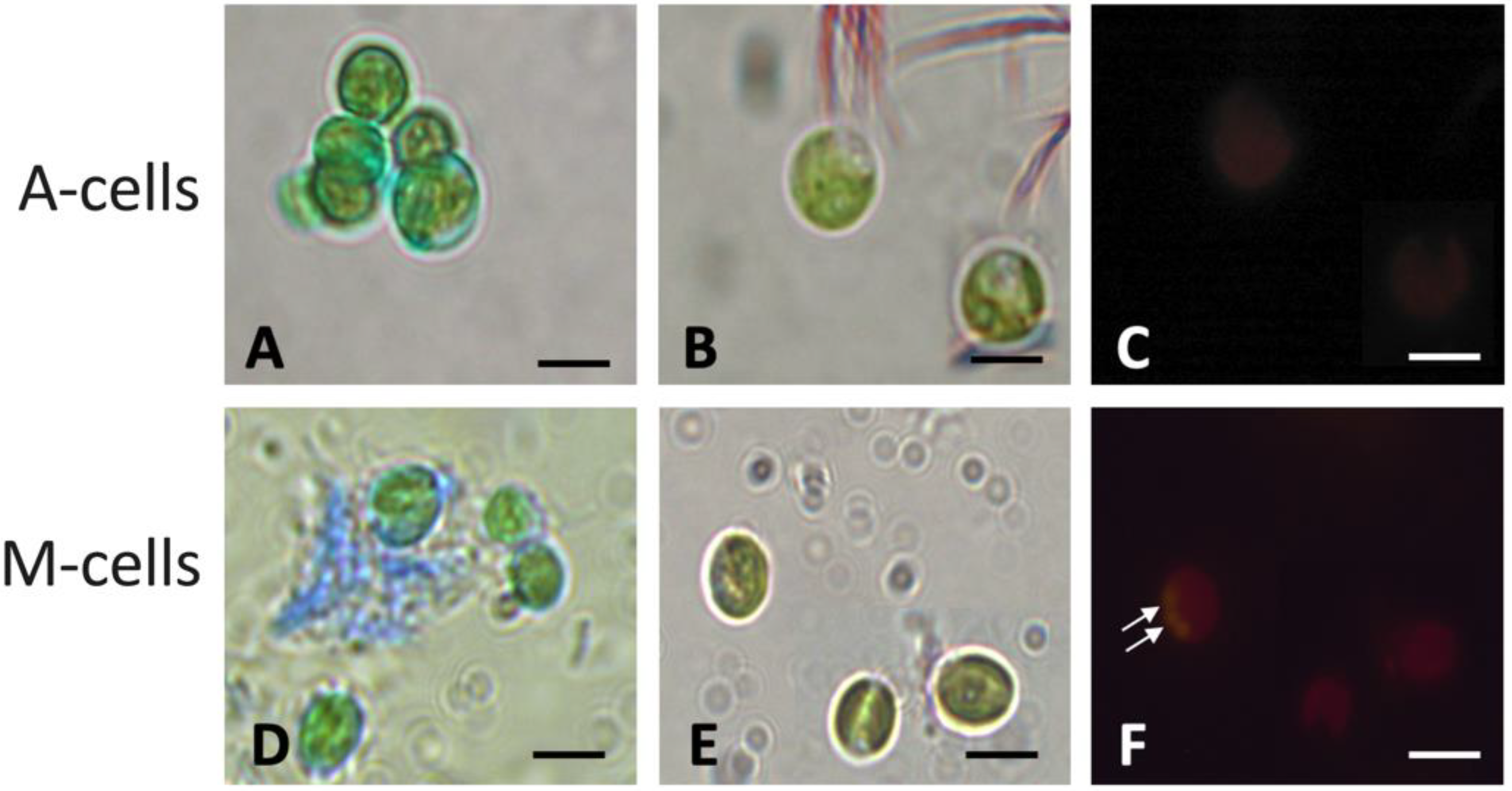
2.3. Observations of Algae Cell Morphology
2.4. Chemical Characterization of Microalgae Extracts Used for Antiviral Tests
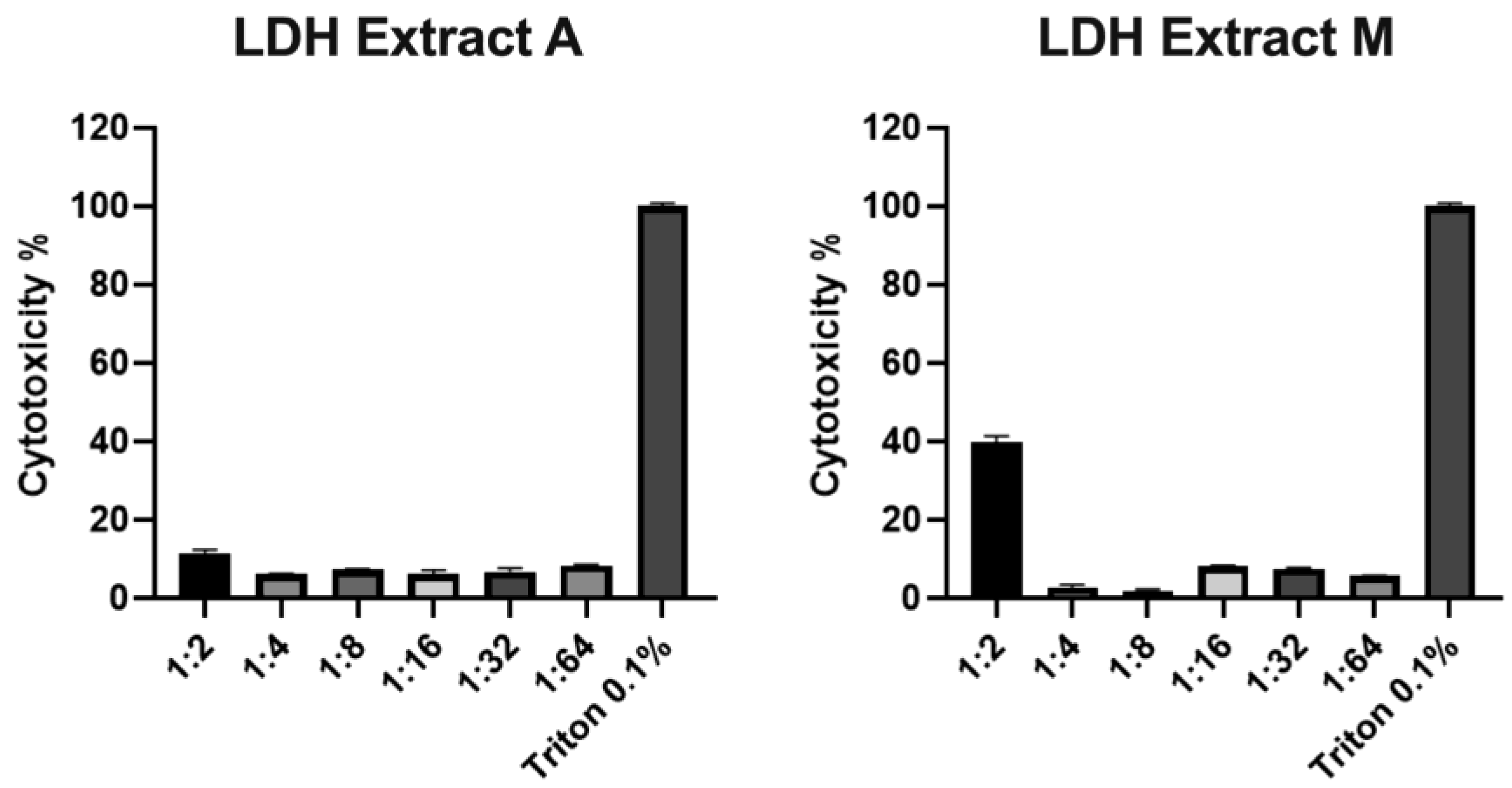
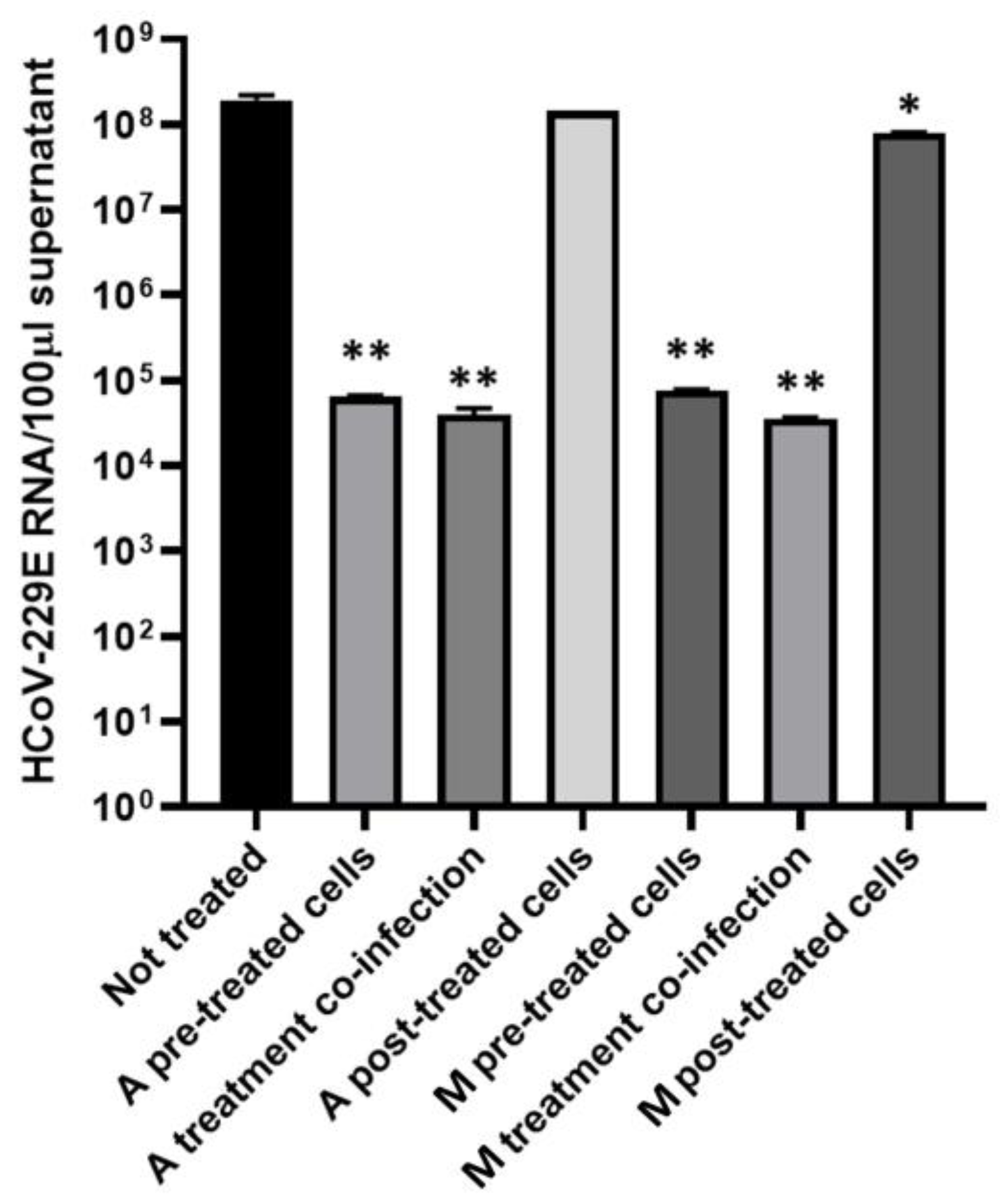
2.5. Antiviral Activity of Microalgae Extracts
3. Discussion
4. Materials and Methods
4.1. Algal Material and Experimental Design
4.2. Growth Evaluations: Cell Density and Dry Biomass of Algae Cultures and pH of Algae Culture Medium
4.3. Evaluation of Algae PSII Maximum Quantum Yield
4.4. Photosynthetic Pigment Extraction and Quantification
4.5. Total Proteins Extraction and Quantification
4.6. Total Phenolics Extraction and Quantification
4.7. Morphological Observations of the Algae
4.7.1. Transmission Electron Microscopy (TEM)
4.7.2. Light Microscopy
4.8. Preparation of Algal Whole Extracts for Antiviral Tests
4.9. Chemical Characterization of Extracts Used for Antiviral Tests
4.10. Cells and Virus
4.10.1. Cell Viability
4.10.2. Antiviral Activity Test
4.11. Statistical Data Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ebenezer, V.; Medlin, L.K.; Ki, J.S. Molecular detection, quantification, and diversity evaluation of microalgae. Mar. Biotechnol. 2012, 14, 129–142. [Google Scholar] [CrossRef]
- Singh, S.K.; Kaur, R.; Bansal, A.; Kapur, S.; Sundaram, S. Biotechnological exploitation of cyanobacteria and microalgae for bioactive compounds. In Biotechnological Production of Bioactive Compounds; Verma, M.L., Chandel, A.K., Eds.; Elsevier: Prayagraj/Jalandhar, India, 2019; pp. 221–259. [Google Scholar] [CrossRef]
- Hopes, A.; Mock, T. Evolution of Microalgae and Their Adaptations in Different Marine Ecosystems; eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2015. [Google Scholar] [CrossRef]
- Metting, F.B., Jr. Biodiversity and application of microalgae. J. Ind. Microbiol. 1996, 17, 477–489. [Google Scholar] [CrossRef]
- Maglie, M.; Baldisserotto, C.; Guerrini, A.; Sabia, A.; Ferroni, L.; Pancaldi, S. A co-cultivation process of Nannochloropsis oculata and Tisochrysis lutea induces morpho-physiological and biochemical variations potentially useful for biotechnological purposes. J. Appl. Phycol. 2021, 33, 2817–2832. [Google Scholar] [CrossRef]
- Borowitzka, M.A. High-value products from microalgae—Their development and commercialization. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Mutanda, T.; Naidoo, D.; Bwapwa, J.K.; Anandraj, A. Biotechnological applications of microalgal oleagineous compounds: Current trends on microalgal bioprocessing of products. Front. Energy Res. 2020, 8, 598803. [Google Scholar] [CrossRef]
- Baldisserotto, C.; Sabia, A.; Guerrini, A.; Demaria, S.; Maglie, M.; Ferroni, L.; Pancaldi, S. Mixotrophic cultivation of Thalassiosira pseudonana with pure and crude glycerol: Impact on lipid profile. Algal Res. 2021, 54, 102194. [Google Scholar] [CrossRef]
- Baldisserotto, C.; Sabia, A.; Giovanardi, M.; Ferroni, L.; Maglie, M.; Pancaldi, S. Chlorophyta microalgae as dietary protein supplement: A comparative analysis of productivity related to photosynthesis. J. Appl. Phycol. 2022, 34, 1323–1340. [Google Scholar] [CrossRef]
- Baldisserotto, C.; Popovich, C.; Giovanardi, M.; Sabia, A.; Ferroni, L.; Constenla, D.; Leonardi, P.; Pancaldi, S. Photosynthetic aspects and lipid profile in the mixotrophic alga Neochloris oleoabundans as useful parameters for biodiesel production. Algal Res. 2016, 16, 255–265. [Google Scholar] [CrossRef]
- McClure, D.D.; Luiz, A.; Gerber, B.; Barton, G.W.; Kavanagh, J.M. An investigation into the effect of culture conditions on fucoxanthin production using the merine microalgae Phaeodactylum tricornutum. Algal Res. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Liu, H.; Su, J.; Lan, C.Q.; Zhong, M.; Hu, X. Production, isolation and bioactive estimation of extracellular polysaccharides of green microalga Neochloris oleoabundans. Algal Res. 2020, 48, 101883. [Google Scholar] [CrossRef]
- Liyanaarachchi, V.C.; Premaratne, M.; Ariyadasa, T.U.; Nimarshana, P.H.V. Two-stage cultivation of microalgae for production of high-value compounds and biofuels: A review. Algal Res. 2021, 57, 102353. [Google Scholar] [CrossRef]
- Zhu, L.D.; Li, Z.H.; Hiltunen, E. Strategies for lipid production improvement in microalgae as a biodiesel feedstock. BioMed. Res. Int. 2016, 2016, 8792548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, T.; Ramos, D.; García-Beltrán, T.; Brito-Bazan, M.; Galindo, E. Mixotrophic cultivation of microalgae: An alternative to produce high-value products. Biochem. Eng. J. 2021, 176, 108183. [Google Scholar] [CrossRef]
- Villanova, V.; Fortunato, A.E.; Singh, D.; Dal Bo, D.; Conte, M.; Obata, T.; Jouhet, J.; Fernie, A.R.; Marechal, E.; Falciatore, E.; et al. Investigating mixotrophic metabolism in the model diatom Phaeodactylum tricornutum. Philos. Trans. R. Soc. B 2017, 372, 20160404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morocho-Jácome, A.L.; Santos, B.B.D.; Carvalho, J.C.M.D.; Almeida, T.S.D.; Rijo, P.; Velasco, M.V.R.; Rosado, C.; Baby, A.M. Microalgae as sustainable, natural-oriented and vegan dermocosmetic bioactive ingredient: The case of Neochloris oleoabundans. Cosmetics 2022, 9, 9. [Google Scholar] [CrossRef]
- Shah, S.A.A.; ul Hassan, S.S.; Bungau, S.; Si, Y.; Xu, H.; Rahman, M.D.H.; Behl, T.; Gitea, D.; Pavel, F.-M.; Corb Aron, R.A.; et al. Chemically diverse and biologically active secondary metabolites from marine Phylum Chlorophyta. Mar. Drugs 2020, 18, 493. [Google Scholar] [CrossRef]
- Carbone, D.A.; Pellone, P.; Lubritto, C.; Ciniglia, C. Evaluation of microalgae antiviral activity and their bioactive compounds. Antibiotics 2021, 10, 746. [Google Scholar] [CrossRef]
- Reynolds, D.; Huesemann, M.; Edmundson, S.; Sims, A.; Hurst, B.; Cady, S.; Beirne, N.; Freeman, J.; Berger, A. Viral inhibitors derived from macroalgae, microalgae, and cyanobacteria: A review of antiviral potential throughout pathogenesis. Algal Res. 2021, 57, 102331. [Google Scholar] [CrossRef]
- Sami, N.; Ahmad, R.; Fatma, T. Exploring algae and cyanobacteria as a promising natural source of antiviral drugs against SARS-CoV-2. Biomed. J. 2020, 44, 54–62. [Google Scholar] [CrossRef]
- Ribeiro, M.C.M.; Salles, T.S.; Ferreira Moreira, M.; Barbarino, E.; do Valle, A.F.; Peixoto Gimenes Couto, M.A. Antiviral activity of microalgae extracts against Mayaro virus. Algal Res. 2022, 61, 102577. [Google Scholar] [CrossRef]
- Santoyo, S.; Jaime, L.; Plaza, M.; Herrero, M.; Rodriguez-Meizoso, I.; Ibañez, E.; Reglero, G. Antiviral compounds obtained from microalgae commonly used as carotenoid sources. J. Appl. Phycol. 2012, 24, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Khavari, F.; Saidijam, M.; Taheri, M.; Nouri, F. Microalgae: Therapeutic potentials and applications. Mol. Biol. Rep. 2021, 48, 4757–4765. [Google Scholar] [CrossRef] [PubMed]
- Tornabene, T.G.; Holzer, G.; Lien, S.; Burris, N. Lipid composition of the nitrogen starved green alga Neochloris oleoabundans. Enzym. Microb. Technol. 1983, 5, 435–440. [Google Scholar] [CrossRef]
- Giovanardi, M.; Ferroni, L.; Baldisserotto, C.; Tedeschi, P.; Maietti, A.; Pantaleoni, L.; Pancaldi, S. Morphophysiological analyses of Neochloris oleoabundans (Chlorophyta) grown mixotrophically in a carbon-rich waste product. Protoplasma 2013, 250, 161–174. [Google Scholar] [CrossRef]
- Karthikeyan, S.; Kalaimurugan, K.; Prathima, A. Quality analysis studies on biodiesel production of Neochloris oleoabundans algae. Energy Sources Part A 2018, 40, 439–445. [Google Scholar] [CrossRef]
- Castro-Puyana, M.; Pérez-Sánchez, A.; Valdés, A.; Ibrahim, O.H.M.; Suarez-Álvarez, S.; Ferragus, J.A.; Micol, V.; Cifuentes, A.; Ibáñez, E.; García-Cañas, V. Pressurized liquid extraction of Neochloris oleoabundans for the recovery of bioactive carotenoids with anti-proliferative activity against human colon cancer cells. Food Res. 2017, 99, 1048–1055. [Google Scholar] [CrossRef] [Green Version]
- Jassim, S.A.A.; Naji, M.A. Novel antiviral agents: A medicinal plant perspective. J. Appl. Microbiol. 2003, 95, 412–427. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Matthews, T.J.; McDanal, C.B.; Bolognesi, D.P.; Greenberg, M.L. A molecular clasp in the human immunodeficiency virus (HIV) type 1 TM protein determines the anti-HIV activity of gp41 derivatives: Implication for viral fusion. J. Virol. 1995, 69, 3771–3775. [Google Scholar] [CrossRef]
- Payne, S. Chapter 17—Family Coronaviridae. In Viruses—from Understanding to Investigation; Academic Press: London, UK, 2017; pp. 149–158. [Google Scholar] [CrossRef]
- Yeo, C.; Kaushal, S.; Yeo, D. Enteric involvement of coronaviruses: Is faecal–oral transmission of SARS-CoV-2 possible? Lancet Gastroenterol. Hepatol. 2020, 5, 335–337. [Google Scholar] [CrossRef] [Green Version]
- Fang, L. Structure, function, and evolution of coronavirus Spike proteins. Ann. Rev. Virol. 2016, 29, 237–261. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; De LeClerq, E. Therapeutic options for the 2019 novel coronavirus (2019-nCoV). Nat. Rev. Drug Discov. 2020, 19, 149–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumla, A.; Chan, J.F.W.; Azhar, E.I.; Hui, D.S.C.; Yuen, K.-Y. Coronaviruses—Drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldisserotto, C.; Giovanardi, M.; Ferroni, L.; Pancaldi, S. Growth, morphology and photosynthetic responses of Neochloris oleoabundans during cultivation in a mixotrophic brackish medium and subsequent starvation. Acta Physiol. Plant. 2014, 36, 461–472. [Google Scholar] [CrossRef]
- Sabia, A.; Baldisserotto, C.; Biondi, S.; Marchesini, R.; Tedeschi, P.; Maietti, A.; Giovanardi, M.; Ferroni, L.; Pancaldi, S. Re-cultivation of Neochloris oleoabundans in exhausted autotrophic and mixotrophic media: The potential role of polyamines and free fatty acids. Appl. Microbiol. Biotechnol. 2015, 99, 10597–10609. [Google Scholar] [CrossRef]
- Giovanardi, M.; Poggioli, M.; Ferroni, L.; Lespinasse, M.; Baldisserotto, C.; Aro, E.-M.; Pancaldi, S. Higher packaging of thylakoid complexes ensures a preserved Photosystem II activity in mixotrophic Neochloris oleoabundans. Algal Res. 2017, 25, 322–332. [Google Scholar] [CrossRef]
- White, S.; Anandraj, A.; Bux, F. PAM fluorometry as a tool to assess microalgal nutrient stress and monitor cellular neutral lipids. Bioresour. Technol. 2011, 102, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Schuurmans, R.M.; van Alphen, P.; Schuurmans, J.M.; Matthijs, H.C.P.; Helligwerf, K.J. Comparison of the photosynthetic yield of cyanobacteria and green algae: Different methods give different answers. PLoS ONE 2015, 10, e0139061. [Google Scholar] [CrossRef] [PubMed]
- Ferroni, L.; Giovanardi, M.; Poggioli, M.; Baldisserotto, C.; Pancaldi, S. Enhanced photosynthetic linear electron flow in mixotrophic green microalga Ettlia oleoabundans UTEX1185. Plant Physiol. Biochem. 2018, 130, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Esteban, R.; Barrutia, O.; Artetxe, U.; Fernàndez-Marìn, B.; Hernàndez, A.; Garcìa-Plazaola, J.I. Internal and external factors affecting photosynthetic pigment composition in plants: A meta-analytical approach. New Phytol. 2015, 206, 268–280. [Google Scholar] [CrossRef]
- Canhoto, J.M. Biotecnologia Vegetal da Clonagem de Plantas à Transformacao Genética; Imprensa da Universidade de Coimbra/Coimbra University Press: Coimbra, Portugal, 2010. [Google Scholar] [CrossRef]
- Ahirwar, A.; Kesharwani, K.; Deka, R.; Muthukumar, S.; Khan, M.J.; Rai, A.; Vinayak, V.; Varjani, S.; Joshi, K.B.; Morjaia, S. Microalgal drugs: A promising therapeutic reserve for the future. J. Biotechnol. 2022, 349, 32–46. [Google Scholar] [CrossRef]
- Fabregas, J.; García, D.; Fernandez-Alonso, M.; Rocha, A.I.; Gómez-Puertas, P.; Escribano, J.M.; Otero, A.; Coll, J.M. In vitro inhibition of the replication of haemorragic septicemia virus (VHSV) and African swine fever virus (ASFV) by extracts from marine microalgae. Antivir. Res. 1999, 44, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Gorelova, O.; Baulina, O.; Ismagulova, T.; Kokabi, K.; Lobakova, E.; Salyakh, I.; Semenova, L.; Chivkunova, O.; Karpova, O.; Scherbakov, P.; et al. Stress-induced changes in the ultrastructure of the photosynthetic apparatus of green microalgae. Protoplasma 2019, 256, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, L.; Komiyama, K. Acidic Polysaccharide CH-1 Isolated from Chlorella pyronoidosa and the Use Thereof. U.S. Patent 4533548, 7 August 1985. [Google Scholar]
- Kanekiyo, K.; Hayashi, K.; Takenaka, H.; Lee, J.-B.; Hayashi, T. Anti-Herpes simplex virus target of an acidic polysaccharide, nostoflan, from the edible blue-green alga Nostoc flagelliforme. Biol. Pharm. Bull. 2007, 30, 1573–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaza, M.; Herrero, H.; Cifuentes, A.; Ibáñez, E. Innovative natural functional ingredients from microalgae. J. Agric. Food Chem. 2007, 57, 7150–7170. [Google Scholar] [CrossRef] [PubMed]
- Popovich, C.; Damiani, C.; Constenla, D.; Martìnez, A.M.; Freije, H.; Giovanardi, M.; Pancaldi, S.; Leonardi, P.I. Neochloris oleoabundans grown in enriched natural seawater for biodiesel feedstock: Evaluation of its growth and biochemical composition. Bioresour. Technol. 2012, 114, 87–293. [Google Scholar] [CrossRef] [PubMed]
- Thormar, H.; Isaacs, C.E.; Brown, H.R.; Barshatzky, M.R.; Pessolano, T. Inactivation of enveloped viruses and killing of cells by fatty acids and monoglycerides. Antimicrob. Agents Chemother. 1987, 31, 27–31. [Google Scholar] [CrossRef]
- Subhash, G.V.; Raja, G.; Kumar, K.; Sapre, A.; Dasgupta, S. Possible prevention of Covid-19 by using linoleic acid (C18) rich algae oil. AIJR 2020. preprints. [Google Scholar] [CrossRef]
- Matich, E.K.; Butryn, D.M.; Ghafari, M.; del Solar, V.; Camgoz, E.; Pfeifer, B.A.; Aga, D.S.; Haznedaroglu, B.Z.; Atill-Gokcumen, G.E. Mass spectrometry-based metabolomics of value-added biochemicals from Ettlia oleoabundans. Algal Res. 2016, 19, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Inès, M.; Dhouba, G. Glycolipid biosurfactants: Potential related biochemical and biotechnological applications. Carbohydr. Res. 2015, 416, 59–69. [Google Scholar] [CrossRef]
- Loya, S.; Reshef, V.; Mizrachi, E.; Silberstein, S.; Rachamim, Y.; Carmeli, S.; Hizi, A. The inhibition of the reverse transcriptase of HIV-1 by the natural sulfoglycolipids from cyanobacteria: Contribution of different moieties to their high potency. J. Nat. Prod. 1998, 61, 891–895. [Google Scholar] [CrossRef]
- Matsumoto, K.; Sakai, H.; Takeuchi, R.; Tsuchiya, K.; Ohta, K.; Sugawara, F.; Abe, M.; Sakaguchi, K. Effective form of sulfoquinovosyldiacyglycerol (SQDG) vesicles for DNA polymerase inhibition. Colloids Surf. B Biointerfaces 2005, 46, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Chirasuwan, N.; Chaiklahan, R.; Kittakoo, P.; Chanasattru, W.; Ruengjitchatchawalya, M.; Tantichereon, M.; Bunnag, B. Anti HSV-1 activity of sulphoquinovosyl diacylglycerol isolated from Spirulina platensis. Sci. Asia 2009, 35, 137–141. [Google Scholar] [CrossRef]
- Safi, C.; Pollio, A.; Olivieri, G. Neochloris oleoabundans from nature to industry: A comprehensive review. Rev. Environ. Sci. Biotechnol. 2021, 20, 943–958. [Google Scholar] [CrossRef]
- Crepin, A.; Caffarri, S. Functions and evolution of Lhcb isoforms composing LHCII, the major light harvesting complex of Photosystem II of green eukaryotic organisms. Curr. Protein Pept. Sci. 2018, 19, 699–713. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.J.; Cavalcanti, V.L.R.; Porto, A.L.F.; Gama, W.A.; Brandão-Costa, R.M.P.; Pedrosa Bezerra, R. The green microalgae Tetradesmus obliquus (Scenedesmus acutus) as lectin source in the recognition of AB0 blood type: Purification and characterization. J. Appl. Phycol. 2020, 32, 103–110. [Google Scholar] [CrossRef]
- Cavalcanti, V.L.R.; Brandão-Costa, R.M.P.; Pontual, E.V.; de Andrade, A.F.; Alves, L.C.; Porto, A.L.F.; Bezerra, R.P. Chlorella vulgaris lectin kills Aedes aegypti larvae. Algal Res. 2021, 56, 102290. [Google Scholar] [CrossRef]
- Roopashi, A.N.; Savitha, J. Screening of freshwater microalgae species for occurrence of lectins and their carbohydrate-binding specificity. J. Appl. Biol. Sci. 2021, 16, 24–34. [Google Scholar] [CrossRef]
- Pang, R.; Tao, J.-Y.; Zhang, S.-L.; Zhao, L.; Yue, X.; Wang, Y.-F.; Ye, P.; Dong, J.-H.; Zhu, Y.; Wu, J.-G. In vitro antiviral activity of lutein against Hepatitis B virus. Phytother. Res. 2010, 24, 1627–1630. [Google Scholar] [CrossRef]
- Urreta, I.; Janices, I.; Ibañez, E.; Castro-Puyana, M.; Castañon, S.; Suárez-Alvarez, S. Valorization of Neochloris oleoabundans biomassa as source of biodiesel by concurrent production of lipids and carotenoids. Algal Res. 2014, 5, 16–22. [Google Scholar] [CrossRef]
- da Silva Ferreira, V.; Sant’Anna, C. Impact of culture conditions on the chlorophyll content of microalgae for biotechnological applications. World J. Microbiol. Biotechnol. 2017, 33, 20. [Google Scholar] [CrossRef]
- Saide, A.; Lauritano, C.; Ianora, A. Pheophorbide a: State of the art. Mar. Drugs 2021, 18, 257. [Google Scholar] [CrossRef] [PubMed]
- Bouslama, L.; Hayashi, K.; Lee, J.B.; Ghorbel, A.; Hayashi, T. Potent virucidal effect of pheophorbide a and pyropheophorbide a on enveloped viruses. J. Nat. Med. 2011, 65, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Paraiso, I.L.; Revel, J.S.; Stevens, J.F. Potential use of polyphenols in the battle against COVID-19. Curr. Opin. Food Sci. 2020, 32, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Qiu, R.; Gao, S.; Lopez, P.A.; Ogden, K.L. Effects of pH on cell growth, lipid production and CO2 addition of microalgae Chlorella sorokiniana. Algal Res. 2017, 28, 192–199. [Google Scholar] [CrossRef]
- González-Camejo, J.; Robles, A.; Seco, A.; Ferrer, J.; Ruano, M.V. On-line monitoring of photosynthetic activity based on pH data to assess microalgae cultivation. J. Environ. Manag. 2020, 276, 111343. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef]
- Baldisserotto, C.; Demaria, S.; Accoto, O.; Marchesini, R.; Zanella, M.; Benetti, L.; Avolio, F.; Maglie, M.; Ferroni, L.; Pancaldi, S. Removal of nitrogen and phosphorus from thickening effluent of an urban wastewater treatment plant by an isolated green microalga. Plants 2020, 9, 1802. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Lowry, H.O.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Haoujar, I.; Cacciola, F.; Abrini, J.; Mangraviti, D.; Giuffrida, D.; El Majdoub, Y.O.; Kounnoun, A.; Miceli, N.; Taviano, M.F.; Mondello, L.; et al. The contribution of carotenoids, phenolic compounds, and flavonoids to the antioxidative properties of marine microalgae isolated from Mediterranean Morocco. Molecules 2019, 24, 4037. [Google Scholar] [CrossRef] [Green Version]
- Baldisserotto, C.; Ferroni, L.; Medici, V.; Pagnoni, A.; Pellizzari, M.; Fasulo, M.P.; Fagioli, F.; Bonora, A.; Pancaldi, S. Specific intra-tissue responses to manganese in the floating lamina of Trapa natans. Plant Biol. 2004, 6, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Mowry, R.W.; Scott, J.E. Observation of the basophilia of amyloids. Histochemie 1967, 10, 8–32. [Google Scholar] [CrossRef] [PubMed]
- Fagan, C.; Dapson, R.W.; Horobin, R.W.; Kiernan, J.A. Revised tests and standards for biological stain commission certification of alcian blue dyes. Biotech. Histochem. 2020, 95, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Discart, V.; Bilad, M.; Vankelecom, I.F. Critical evaluation of the determination methods for transparent exopolymer particles, agents of membrane fouling. Crit. Rev. Environ. Sci. Technol. 2015, 45, 167–192. [Google Scholar] [CrossRef]
- Vergnes, J.B.; Gernigon, V.; Guiraud, P.; Formosa-Dague, C. Bicarbonate concentration induces production of exopolysaccharides by Arthrospira platensis that mediate bioflocculation and enhance flotation harvesting efficiency. ACS Sustain. Chem. Eng. 2019, 7, 13796–13804. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract A | Extract M | |
|---|---|---|
| Chla (μg mL−1) | 215.7 ± 1.69 | 65.71 ± 9.69 *** |
| Chlb (μg mL−1) | 97.20 ± 0.33 | 38.03 ± 3.83 *** |
| Cars (μg mL−1) | 14.59 ± 0.21 | 4.47 ± 2.18 ** |
| Total phenolics (μgCoum.Ac.Eq. mL−1) | 55.21 ± 10.16 | 42.27 ± 4.95 ** |
| Total proteins (μg μL−1) | 1.47 ± 0.28 | 2.86 ± 0.56 *** |
| Total lipids (μg μL−1) Fatty acids composition (% total FA) | - | 1.01 ± 0.08 |
| Palmitic acid (16:0) | - | 54.45 ± 4.22 |
| Stearic acid (18:0) | - | 13.37 ± 1.28 |
| Oleic acid (18:1) | - | 11.38 ± 1.65 |
| Linoleico (18:2) | - | 20.79 ± 0.70 |
| PFU mL−1 | |
|---|---|
| Starting inoculum (T = 0) | 5.2 × 104 |
| Untreated HCoV-229E (T = 1 h) | 3.85 × 104 |
| HCoV-229E + extract A | 0 |
| HCoV-229E + extract M | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldisserotto, C.; Gentili, V.; Rizzo, R.; Di Donna, C.; Ardondi, L.; Maietti, A.; Pancaldi, S. Characterization of Neochloris oleoabundans under Different Cultivation Modes and First Results on Bioactivity of Its Extracts against HCoV-229E Virus. Plants 2023, 12, 26. https://doi.org/10.3390/plants12010026
Baldisserotto C, Gentili V, Rizzo R, Di Donna C, Ardondi L, Maietti A, Pancaldi S. Characterization of Neochloris oleoabundans under Different Cultivation Modes and First Results on Bioactivity of Its Extracts against HCoV-229E Virus. Plants. 2023; 12(1):26. https://doi.org/10.3390/plants12010026
Chicago/Turabian StyleBaldisserotto, Costanza, Valentina Gentili, Roberta Rizzo, Chiara Di Donna, Luna Ardondi, Annalisa Maietti, and Simonetta Pancaldi. 2023. "Characterization of Neochloris oleoabundans under Different Cultivation Modes and First Results on Bioactivity of Its Extracts against HCoV-229E Virus" Plants 12, no. 1: 26. https://doi.org/10.3390/plants12010026