

Transgressive Biochemical Response to Water Stress in Interspecific Eggplant Hybrids
 , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction
2. Results
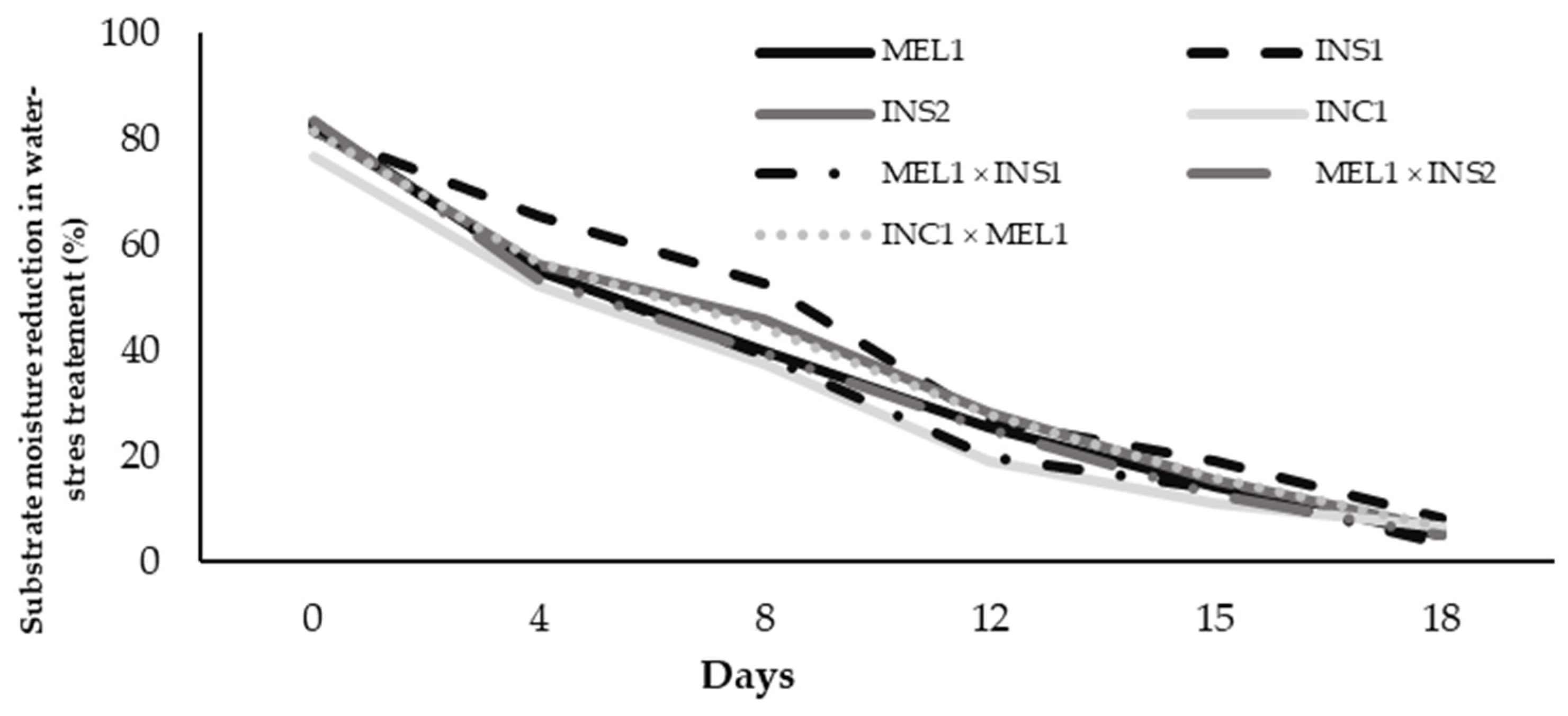
2.1. Substrate Moisture during the Water Stress Treatments
2.2. Growth Response of S. melongena, S. insanum and S. incanum during Water Stress
2.3. Biochemical Parameters Quantified in Parental Accessions
2.4. Comparison of the Response of the Hybrids with Respect to Their Parents
2.5. Criteria for Assessing Plant Tolerance
3. Discussion
3.1. Growth and Biochemical Responses of Parental Species
3.2. Evaluation of Hybrids’ Responses
3.3. Future Perspectives
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design and Growth Conditions
4.3. Measurement of Growth Parameters and Water Use Efficiency
4.4. Biochemical Analyses
4.4.1. Malondialdehyde (MDA) Determination
4.4.2. Total Phenolic Compounds (TPC)
4.4.3. Total Flavonoids (TF)
4.4.4. Proline (Pro) Determination
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vorontsova, M.S.; Knapp, S. A Revision of the “Spiny Solanums,” Solanum Subgenus Leptostemonum (Solanaceae), in Africa and Madagascar. Syst. Bot. Monogr. 2016, 99, 432. [Google Scholar]
- Sharma, M.; Kaushnik, P. Biochemical composition of eggplant fruits: A review. Appl. Sci. 2021, 11, 7078. [Google Scholar] [CrossRef]
- Bouhajeb, R.; Selmi, S.; Nakbi, A.; Jlassi, I.; Montevecchi, G.; Flamini, G.; Zarrad, I.; Dabbou, S. Chemical composition analysis, antioxidant, and antibacterial activities of eggplant leaves. Chem. Biodivers. 2020, 17, e2000405. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Production Databases. 2017. Available online: http://www.fao.org/faostat/en/#home (accessed on 18 April 2022).
- Alam, I.; Salimullah, M. Genetic engineering of eggplant (Solanum melongena L.): Progress, controversy and potential. Horticulturae 2021, 7, 78. [Google Scholar] [CrossRef]
- Fu, Q.S.; Yang, R.C.; Wang, H.S.; Zhao, B.; Zhou, C.L.; Ren, S.X.; Guo, Y.D. Leaf morphological and ultrastructural performance of eggplant (Solanum melongena L.) in response to water stress. Photosynthetica. 2013, 51, 109–114. [Google Scholar] [CrossRef]
- Darko, R.O.; Yuan, S.; Kumi, F.; Quaye, F. Effect of deficit irrigation on yield and quality of eggplant. Int. J. Agric. Environ. Biotechnol. 2019, 4, 1325–1333. [Google Scholar] [CrossRef]
- Kouassi, A.B.; Kouassi, K.B.A.; Sylla, Z.; Plazas, M.; Fonseka, R.M.; Kouassi, A.; Fonseka, A.; N’guetta, A.S.-P. Genetic parameters of drought tolerance for agromorphological traits in eggplant, wild relatives, and interspecific hybrids. Crop Sci. 2021, 61, 55–68. [Google Scholar] [CrossRef]
- Gramazio, P.; Prohes, J.; Plazas, M.; Mangino, G.; Herraiz, F.J.; Vilanova, S. Development and genetic characterization of advanced backcross materials and an introgression line population of Solanum incanum in a S. melongena background. Front. Plant Sci. 2017, 8, 1477. [Google Scholar] [CrossRef] [Green Version]
- Mastretta-Yanes, A.; Acevedo Gasman, F.; Burgeff, C.; Cano Ramírez, M.; Piñero, D.; Sarukhán, J. An initiative for the study and use of genetic diversity of domesticated plants and their wild relatives. Front. Plant Sci. 2019, 9, 209. [Google Scholar] [CrossRef] [Green Version]
- Warschefsky, E.J.; Rieseberg, L.H. Laying the groundwork for crop wild relative conservation in the United States. Proc. Natl. Acad. Sci. USA 2021, 118, e2024375118. [Google Scholar] [CrossRef]
- Kaushik, P. Line × tester analysis for morphological and fruit biochemical traits in eggplant (Solanum melongena L.) using wild relatives as testers. Agronomy 2019, 9, 185. [Google Scholar] [CrossRef]
- Davidar, P.; Snow, A.A.; Rajkumar, M.; Pasquet, R.; Daunay, M.C.; Mutegi, E. The potential for crop to wild hybridization in eggplant (Solanum melongena; Solanaceae) in southern India. Am. J. Bot. 2015, 102, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plazas, M.; Vilanova, S.; Gramazio, P.; Rodríguez-Burruezo, A.; Fita, A.; Herraiz, F.J.; Prohens, J. Interspecific hybridization between eggplant and wild relatives from different genepools. J. Am. Soc. Hortic. Sci. 2016, 141, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Fita, A.; Rodriguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef] [Green Version]
- Oladosu, Y.; Rafii, M.Y.; Arolu, F.; Chukwu, S.C.; Salisu, M.A.; Olaniyan, B.A.; Fagbohun, I.K.; Muftaudeen, T.K. Genetic diversity and utilization of cultivated eggplant germplasm in varietal improvement. Plants 2021, 10, 1714. [Google Scholar] [CrossRef]
- Pratap, A.; Das, A.; Kumar, S.; Gupta, S. Current perspectives on introgression Breeding in Food Legumes. Front. Plant Sci. 2021, 11, 589189. [Google Scholar] [CrossRef]
- Kaushik, P.; Prohens, J.; Vilanova, S.; Gramazio, P.; Plazas, M. Phenotyping of eggplant wild relatives and interspecific hybrids with conventional and phenomics descriptors provides insight for their potential utilization in breeding. Front. Plant Sci. 2016, 7, 677. [Google Scholar] [CrossRef] [Green Version]
- Hajjar, R.; Hodgkin, T. The use of wild relatives in crop improvement: A survey of developments over the last 20 years. Euphytica 2007, 156, 1–13. [Google Scholar] [CrossRef]
- Bhavyasree, R.K.; Singh, S.; Singh, I. Advanced backcross strategy for alien introgression for productivity enhancing traits in chickpea (Cicer arietinum L.). Legume Res. 2018, 41, 379–383. [Google Scholar] [CrossRef]
- Harlan, J.R.; De Wet, J.M.J. Toward a rational classification of cultivated plants. Taxon 1971, 20, 509–517. [Google Scholar] [CrossRef]
- Ranil, R.H.G.; Prohens, J.; Aubriot, X.; Niran, H.M.L.; Plazas, M.; Fonseka, R.M.; Vilanova, S.; Fonseka, H.H.; Gramazio, P.; Knap, S. Solanum insanum L. (subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant (S. melongena L.): A review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genet. Resour. Crop Evol. 2017, 64, 1707–1722. [Google Scholar] [CrossRef]
- Page, A.; Gibson, J.; Meyer, R.S.; Chapman, M.A. Eggplant domestication: Pervasive gene flow, feralization, and transcriptomic divergence. Mol. Biol. Evol. 2019, 36, 1359–1372. [Google Scholar] [CrossRef] [PubMed]
- Barchi, L.; Rabanus-Wallace, M.T.; Prohens, J.; Toppino, L.; Padmarasu, S.; Portis, E.; Rotino, G.L.; Stein, N.; Lanteri, S.; Giuliano, G. Improved genome assembly and pan-genome provide key insights into eggplant domestication and breeding. Plant J. 2021, 107, 579–596. [Google Scholar] [CrossRef] [PubMed]
- Weese, T.L.; Bohs, L. Eggplant origins: Out of Africa, into the Orient. Taxon. 2010, 59, 49–56. [Google Scholar] [CrossRef]
- Syfert, M.M.; Castañeda-Álvarez, N.P.; Khoury, C.K.; Särkinen, T.; Sosa, C.C.; Achicanoy, H.A.; Bernau, V.; Prohens, J.; Daunay, M.-C.; Knapp, S. Crop wild relatives of the brinjal eggplant (Solanum melongena): Poorly represented in genebanks and many species at risk of extinction. Am. J. Bot. 2016, 103, 635–651. [Google Scholar] [CrossRef] [Green Version]
- Plazas, M.; González-Orenga, S.; Nguyen, H.T.; Morar, I.M.; Fita, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Growth and antioxidant responses triggered by water stress in wild relatives of eggplant. Sci. Hortic. 2022, 293, 110685. [Google Scholar] [CrossRef]
- Arafa, R.A.; Prohens, J.; Solberg, S.Ø.; Plazas, M.; Rakh, M. Breeding and genome mapping for resistance to biotic stress in eggplant. In Genomic Designing for Biotic Stress Resistant Vegetable Crops; Kole, C., Ed.; Springer: Cham, Germany, 2022. [Google Scholar] [CrossRef]
- Chaves, M.M.; Costa, J.M.; Saibo, N.J.M. Recent advances in photosynthesis under drought and salinity. Adv. Bot. Res. 2011, 57, 49–104. [Google Scholar]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Env. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Dudek, B.; Warskulat, A.-C.; Schneider, B. The occurrence of flavonoids and related compounds in flower sections of Papaver nudicaule. Plants 2016, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Martinez, V.; Mestrem, T.C.; Rubio, F.; Girones-Vilaplana, A.; Moreno, D.A.; Mittler, R.; Rivero, R.M. Accumulation of flavonols over hydroxycinnamic acids favors oxidative damage protection under abiotic stress. Front. Plant Sci. 2016, 7, 838. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in biotic and abiotic stress responses and microbial associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Apel, K.Y.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 25, 800. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Peleegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Khalil, F.; Rauf, S.; Monneveux, P.; Anwar, S.; Iqbal, Z. Genetic analysis of proline concentration under osmotic stress in sunflower (Helianthus annuus L.). Breed Sci. 2016, 66, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef]
- Jiménez, S.; Dridi, J.; Gutiérrez, D.; Moret, D.; Irigoyen, J.J.; Moreno, M.A.; Gogorcena, Y. Physiological, biochemical and molecular responses in four Prunus rootstocks submitted to drought stress. Tree Physiol. 2013, 33, 1061–1075. [Google Scholar] [CrossRef]
- Escalante-Magaña, C.; Aguilar-Caamal, L.F.; Echevarría-Machado, I.; Medina-Lara, F.; Sánchez Cach, L.; Martínez-Estévez, M. Contribution of Glycine Betaine and Proline to Water Deficit Tolerance in Pepper Plants. Hortscience 2019, 54, 1044–1054. [Google Scholar] [CrossRef] [Green Version]
- Gulen, H.; Kesici, M.; Cetinkaya, C.; Ergin, S. Proline and antioxidant enzyme activities in some strawberry cultivars under drought and recovery. Not. Bot. Horti. Agrobo. 2018, 46, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Lv, B.S.; Ma, H.Y.; Li, X.W.; Wei, L.X.; Lv, H.Y.; Yang, H.Y.; Jiang, C.J.; Liang, Z.W. Proline accumulation is not correlated with saline-alkaline stress tolerance in rice seedlings. Agron. J. 2015, 107, 51–60. [Google Scholar] [CrossRef]
- Silvente, S.; Sobolev, A.P.; Lara, M. Metabolite adjustments in drought tolerant and sensitive soybean genotypes in response to water stress. PloS ONE 2012, 7, e38554. [Google Scholar] [CrossRef] [Green Version]
- Arteaga, S.; Yabor, L.; Díez, M.J.; Prohens, J.; Boscaiu, M.; Vicente, O. The use of proline in screening for tolerance to drought and salinity in common bean (Phaseolus vulgaris L.) genotypes. Agronomy 2020, 10, 817. [Google Scholar] [CrossRef]
- Bechtold, U.; Field, B. Molecular mechanisms controlling plant growth during abiotic stress. J. Exp. Bot. 2018, 69, 2753–2758. [Google Scholar] [CrossRef] [Green Version]
- Toscano, S.; Ferrante, A.; Romano, D. Response of mediterranean ornamental plants to drought stress. Horticulturae 2019, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [Green Version]
- Rahma, N.A.F. Screening for drought tolerance in eggplants relatives and Interspecific hybrids (Solanum spp.). Master Thesis, Polytechnic University of Valencia, Valencia, Spain, 2016. Available online: https://riunet.upv.es/bitstream/handle/10251/68707/RAHMA (accessed on 3 November 2022).
- Munné-Bosch, S.; Alegre, L. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef]
- Zimmermann, P.; Zentgraf, U. The correlation between oxidative stress and leaf senescence during plant development. Cell. Mol. Biol. Lett. 2005, 10, 515–534. [Google Scholar]
- Díaz-Pérez, J.C.; Eaton, T.E. Eggplant (Solanum melongena L.) plant growth and fruit yield as affected by drip irrigation rate. HortScience. 2015, 50, 1709–1714. [Google Scholar] [CrossRef]
- Sarker, B.C.; Hara, M.; Uemura, M. Comparison of response of two C3 species to leaf water relation, proline synthesis, gas exchange and water use under periodic water stress. J. Plant Biol. 2004, 47, 33–41. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Env. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Hsu, S.Y.; Hsu, Y.T.; Kao, C.H. The effect of polyethylene glycol on proline accumulation in rice leaves. Biol. Plant. 2003, 46, 73–78. [Google Scholar] [CrossRef]
- Lei, Y.; Yin, C.; Li, C. Differences in some morphological, physiological, and biochemical responses to drought stress in two contrasting populations of Populus przewalskii. Physiol. Plant. 2006, 127, 182–191. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids. 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Kurniawati, S. Drought Stress Tolerance Mechanisms of Eggplant (Solanum spp.): Physiology and Molecular. Master’s Thesis, IPB University, Bogor, Indonesia, 2014. Available online: http://repository.ipb.ac.id/handle/123456789/68351 (accessed on 1 December 2022).
- Plazas, M.; Nguyen, T.; González-Orenga, S.; Fita, A.; Vicente, O.; Prohens, J.; Boscaiu, M. Comparative analysis of the responses to water stress in eggplant (Solanum melongena) cultivars. Plant Physiol. Biochem. 2019, 143, 72–82. [Google Scholar] [CrossRef]
- Tani, E.; Kizis, D.; Markellou, E.; Papadakis, I.; Tsamadia, D.; Leventis, G.; Karapanaos, I. Cultivar-dependent responses of eggplant (Solanum melongena L.) to simultaneous Verticillium dahliae infection and drought. Front. Plant Sci. 2018, 9, 1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, B.C.; Hara, M.; Uemura, M. Proline synthesis, physiological responses and biomass yield of eggplants during and after repetitive soil moisture stress. Sci. Hort. 2005, 103, 387–402. [Google Scholar] [CrossRef]
- Heuer, B. Role of proline in plant response to drought and salinity. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar] [CrossRef]
- Pollastri, S.; Tattini, M. Flavonols: Old compounds for old roles. Ann. Bot. 2011, 108, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Vicente, O.; Boscaiu, M. Flavonoids: Antioxidant compounds for plant defence… and for a healthy human diet. Not. Bot. Horti Agrobo. 2017, 46, 1421. [Google Scholar] [CrossRef] [Green Version]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. J. Plant Physiol. 2011, 168, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Bautista, I.; Boscaiu, M.; Lidón, A.; Llinares, J.V.; Lull, C.; Donat, M.P.; Vicente, O. Environmentally induced changes in antioxidant phenolic compounds levels in wild plants. Acta Physiol. Plant. 2016, 38, 9. [Google Scholar] [CrossRef]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Nisha, P.; Abdul Nazar, P.; Jayamurthy, P. A comparative study on antioxidant activities of different varieties of Solanum melongena. Food Chem. Toxicol. 2009, 47, 2640–2644. [Google Scholar] [CrossRef]
- Ylstra, B.; Touraev, A.; Benito, R.M.; Stoger, E.; Van Tunen, A.J.; Vicente, O.; Mol, J.N.M.; Heberle-Bors, E. Flavonols stimulate development, germination, and tube growth of tobacco pollen. Plant Physiol. 1992, 100, 902–907. [Google Scholar] [CrossRef] [Green Version]
- Gould, K.; Lister, C. Flavonoid functions in plants. In Flavonoids: Chemistry, Biochemistry and Applications; Andersen, P.M., Markham, K.R., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 397–441. [Google Scholar] [CrossRef]
- Kashyap, A.; Garg, P.; Tanwar, K.; Sharma, J.; Gupta, N.C.; Thi Thu Ha, P.; Bhattacharya, R.C.; Mason, A.S. Strategies for utilization of crop wild relatives in plant breeding programs. Theor. Appl. Genet. 2022, 135, 4151–4167. [Google Scholar] [CrossRef]
- Prohens, J.; Gramazio, P.; Plazas, M.; Dempewolf, H.; Kilian, B.; Díez, M.J.; Fita, A.; Herraiz, F.J.; Rodríguez-Burruezo, A.; Knapp, S.; et al. Introgressiomics: A new approach for using crop wild relatives in breeding for adaptation to climate change. Euphytica 2017, 213, 158. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Liu, Y.; Zhang, Y.; Gu, R. Advances in research on the mechanism of heterosis in plants. Front. Plant Sci. 2021, 12, 745726. [Google Scholar] [CrossRef]
- Geng, X.; Qu, Y.; Jia, Y.; He, S.; Pan, Z.; Wang, L.; Du, X. Assessment of heterosis based on parental genetic distance estimated with SSR and SNP markers in upland cotton (Gossypium hirsutum L.). BMC Genomics 2021, 22, 123. [Google Scholar] [CrossRef]
- Betran, F.J.; Ribaut, J.M.; Beck, D.; de Leon, D.G. Genetic diversity, specific combining ability, and heterosis in tropical maize under stress and nonstress environments. Crop Sci. 2003, 43, 797–806. [Google Scholar] [CrossRef]
- Nan, X.; Huihui, Z.; Haixiu, Z.; Yining, W.; Jinbo, L.; Li, X.; Zepeng, Y.; Wenxu, Z.; Yi, Q.; Guangyu, S. The response of photosynthetic functions of F1 cutting seedlings from Physocarpus amurensis Maxim (♀) × Physocarpus opulifolius “Diabolo” (♂) and the parental seedlings to salt Stress. Front Plant Sci. 2018, 9, 714. [Google Scholar] [CrossRef]
- Farooq, M.A.; Shakeel, A.; Chattha, W.S.; Tahir, M. Two-year study on combining ability and heterotic potential in functional traits under salt stress in upland cotton (Gossypium hirsutum). Plant Breed. 2020, 139, 139–1221. [Google Scholar] [CrossRef]
- Zhao, Y.; Hu, F.; Zhang, X.; Wei, Q.; Dong, j.; Bo, C.; Cheng, B.; Ma, Q. Comparative transcriptome analysis reveals important roles of nonadditive genes in maize hybrid An’nong 591 under heat stress. BMC Plant Biol. 2019, 19, 273. [Google Scholar] [CrossRef] [Green Version]
- Trovato, M.; Forlani, G.; Signorelli, S.; Funck, D. Proline metabolism and its functions in development and stress tolerance. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants; Hossain, M., Kumar, V., Burritt, D., Fujita, M., Mäkelä, P., Eds.; Springer: Cham, Germany, 2019. [Google Scholar] [CrossRef]
- Forlani, G.; Trovato, M.; Funck, D.; Signorelli, S. Regulation of proline accumulation and its molecular and physiological functions in stress defence. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants; Hossain, M.A., Kumar, V., Burritt, D.J., Fujita, M., Mäkelä, P.S.A., Eds.; Springer: Cham, Germany, 2019; pp. 73–97. [Google Scholar]
- de Oliveira, M.M.T.; Shuhua, L.; Kumbha, D.S.; Zurgil, U.; Raveh, E.; Tel-Zur, N. Performance of Hylocereus (Cactaceae) species and interspecific hybrids under high-temperature stress. Plant Physiol. Biochem. 2020, 153, 30–39. [Google Scholar] [CrossRef]
- Bajpai, P.K.; Reichelt, M.; Augustine, R.; Gershenzon, J.; Bisht, N.C. Heterotic patterns of primary and secondary metabolites in the oilseed crop Brassica juncea. Heredity 2019, 123, 318–336. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, T.; Verma, M.K.; Singh, S.K.; Thakre, M.; Sharma, R.R.; Jaiswal, S. Heterosis and heterobeltiosis for morpho-physical, phenolics, flavonoids and antioxidants in grape (Vitis vinifera) hybrids. Indian J. Agric. Sci. 2017, 87, 759–764. [Google Scholar]
- Pabuayon, I.C.; Kitazumi, A.; Cushman, K.R.; Singh, R.K.; Gregorio, G.B.; Dhatt, B.; Zabet-Moghaddam, M.; Walia, H.; de Los Reyes, B.G. Novel and transgressive salinity tolerance in recombinant inbred lines of rice created by physiological coupling-uncoupling and network rewiring effects. Front. Plant Sci. 2021, 12, 615277. [Google Scholar] [CrossRef] [PubMed]
- Kouassi, B.; Prohens, J.; Gramazio, P.; Kouassi, A.B.; Vilanova, S.; Galán-Ávila, A.; Herraiz, F.J.; Kouassi, A.; Segui, J.M.; Plazas, M. Development of backcross generations and new interspecific hybrid combinations for introgression breeding in eggplant (Solanum melongena). Sci. Hort. 2016, 213, 199–207. [Google Scholar] [CrossRef]
- Ranil, R.H.; Niran, H.M.L.; Plazas, M.; Fonseka, R.; Fonseka, H.H.; Vilanova, S.; Andújar, I.; Gramazio, P.; Fita, A.; Prohens, J. Improving seed germination of the eggplant rootstock Solanum torvum by testing multiple factors using an orthogonal array design. Sci. Hort. 2015, 193, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Rasband, W.S.; ImageJ. U.S. National Institutes of Health, Bethesda, MD, USA, 1997–2018. Available online: https://imagej.nih.gov/ij/ (accessed on 2 November 2022).
- Hodges, D.M.; Delong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. Available online: https://www.ajevonline.org/content/16/3/144 (accessed on 2 November 2022).
- Plazas, M.; Prohens, J.; Cuñat, A.N.; Vilanova, S.; Gramazio, P.; Herraiz, F.J.; Andújar, I. Reducing capacity, chlorogenic acid content and biological activity in a collection of scarlet (Solanum aethiopicum) and gboma (S. macrocarpon) eggplants. Int. J. Mol. Sci. 2014, 15, 17221–17241. [Google Scholar] [CrossRef] [Green Version]
- Zhishen, J.; Mengchen, T.; Jianmin, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Comstock, R.E.; Robinson, A.; Gowen, J.W. Estimation of average dominance of genes. In Heterosis; Gowen, J.W., Ed.; Iowa State College Press: Ames, IA, USA, 1952; pp. 494–516. [Google Scholar]
- Hayman, B.I. Interaction, heterosis and diallel crosses. Genetics 1957, 42, 336–355. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effects | gl | RFW | RDW | RWC | SH | Lno | LA | LFW | LDW | LWC |
|---|---|---|---|---|---|---|---|---|---|---|
| MAIN | ||||||||||
| A: Genotype | 3 | 20.97 * | 1.08 *** | 11.04 NS | 65.12 *** | 12.33 ** | 3812 ** | 71.65 ** | 1.22 NS | 14.9 NS |
| B: Treatment | 1 | 62.70 ** | 0.16 NS | 1407 *** | 64.85 *** | 38.12 *** | 37963 *** | 1174 *** | 12.17 *** | 105 *** |
| INTERACTION | ||||||||||
| AB | 3 | 7.48 NS | 0.14NS | 128.24 *** | 3.89 NS | 1.63 NS | 1047 NS | 25.81 NS | 0.09 NS | 95.18 ** |
| RESIDUALS | 30 | 5.88 | 0.21 | 11.53 | 3.97 | 2.30 | 594 | 12.35 | 0.66 | 15.10 |
| Effects | gl | MDA | TPC | TF | Pro |
|---|---|---|---|---|---|
| MAIN | |||||
| A: Genotype | 3 | 9134 *** | 483 *** | 41.64 *** | 1230 NS |
| B: Treatment | 1 | 9858 *** | 150 *** | 71.55 *** | 184558 *** |
| INTERACTION | |||||
| AB | 3 | 1257 * | 15.42 NS | 6.64 NS | 2674 NS |
| RESIDUALS | 30 | 6.10 | 2.28 | 3622 |
| MEL1 × INS1 | MEL1 × INS2 | INC1 × MEL1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F1 | Pm | Ratio | P | F1 | Pm | Ratio | P | F1 | Pm | Ratio | P | ||
| Leaf Area | C | 153.05 | 123.83 | 1.24 | 0.84 | 162.30 | 151.95 | 1.07 | 1.57 | 148.27 | 154.67 | 0.96 | −1.66 |
| S | 59.54 | 72.90 | 0.82 | −0.94 | 69.40 | 79.51 | 0.87 | −1.33 | 60.24 | 79.60 | 0.76 * | −2.57 | |
| Stem Height | C | 18.60 | 14.85 | 1.25 * | 1.42 | 14.10 | 15.10 | 0.93 | −0.35 | 14.60 | 14.60 | 1.00 | 0 |
| S | 15.50 | 12.20 | 1.27 | 1.83 | 14.10 | 13.55 | 1.04 | 0.18 | 15.36 | 11.75 | 1.31 * | 2.67 | |
| Total Water Content | C | 78.72 | 79.41 | 0.99 | −0.20 | 82.32 | 81.78 | 1.01 | 0.52 | 80.89 | 81.59 | 0.99 | −0.57 |
| S | 60.33 | 69.46 | 0.87 * | −3.92 | 60.56 | 67.23 | 0.90 | −62.46 | 63.56 | 67.02 | 0.95 | −31.44 | |
| Leaf Water Content | C | 82.18 | 81.83 | 1.00 | 0.14 | 83.55 | 83.13 | 1.00 | 0.34 | 82.69 | 82.61 | 1.00 | 0.05 |
| S | 59.09 | 69.34 | 0.85 * | −2.03 | 54.76 | 65.11 | 0.84 | −12.61 | 63.55 | 67.06 | 0.95 | −1.27 | |
| Root Water Content | C | 78.69 | 79.99 | 0.98 | −0.44 | 84.49 | 83.17 | 1.02 | 1.91 | 84.20 | 83.84 | 1.00 | 0.26 |
| S | 65.43 | 73.87 | 0.89 | −24.2 | 64.07 | 70.55 | 0.91 | −12.61 | 69.00 | 68.09 | 1.01 | 0.24 | |
| Leaf Fresh Weight | C | 29.89 | 20.35 | 1.47 * | 1.98 | 24.59 | 24.07 | 1.02 | 0.46 | 22.03 | 21.67 | 1.02 | 0.10 |
| S | 11.22 | 9.41 | 1.19 | 2.34 | 12.16 | 10.41 | 1.17 | 7.82 | 9.28 | 8.95 | 1.04 | 0.27 | |
| Leaf Dry Weight | C | 5.17 | 3.39 | 1.52 * | 4.79 | 3.93 | 3.94 | 1.00 | 0.06 | 4.10 | 3.76 | 1.09 | 54.74 |
| S | 3.55 | 2.41 | 1.47 * | 4.34 | 3.58 | 2.76 | 1.30 | 9.71 | 2.94 | 2.56 | 1.15 | 3.34 | |
| Root Fresh Weight | C | 11.96 | 7.55 | 1.58 * | 2.15 | 13.54 | 9.30 | 1.46 * | 13.77 | 11.22 | 8.76 | 1.28 * | 2.91 |
| S | 7.50 | 6.07 | 1.24 | 1.64 | 6.06 | 6.68 | 0.91 | −2.34 | 8.80 | 5.04 | 1.75 * | 1.98 | |
| Root Dry Weight | C | 2.55 | 1.60 | 1.60 * | 3.24 | 2.11 | 1.67 | 1.26 | 2.00 | 1.76 | 1.49 | 1.19 | 0.65 |
| S | 2.65 | 1.62 | 1.63 | 3.42 | 2.17 | 1.93 | 1.12 | 24.20 | 2.71 | 1.54 | 1.76 * | 3.10 | |
| Total Fresh Weight | C | 41.85 | 29.99 | 1.40 * | 2.03 | 38.13 | 33.37 | 1.14 | 3.35 | 33.25 | 30.44 | 1.09 | 0.65 |
| S | 18.72 | 15.47 | 1.21 | 1.97 | 18.22 | 17.08 | 1.07 | 27.59 | 18.08 | 13.99 | 1.29 * | 1.30 | |
| Total Dry Weight | C | 7.72 | 4.87 | 1.59 * | 5.25 | 6.04 | 5.49 | 1.10 | 7.20 | 5.87 | 5.16 | 1.14 | 2.84 |
| S | 6.20 | 4.031 | 1.54 * | 3.85 | 5.75 | 4.69 | 1.23 | 11.25 | 5.65 | 4.10 | 1.38 * | 3.15 | |
| Foliar MDA | C | 37.33 | 66.43 | 0.56 * | −0.99 | 39.45 | 87.11 | 0.45 * | −5.48 | 42.43 | 66.16 | 0.64 * | −1.00 |
| S | 69.29 | 85.76 | 0.81 | −0.43 | 84.02 | 129.25 | 0.65 * | −8.50 | 89.36 | 101.44 | 0.88 | −0.54 | |
| Foliar Proline | C | 33.90 | 28.91 | 1.17 | 2.65 | 15.42 | 31.58 | 0.49 * | −3.83 | 6.64 | 33.49 | 0.20 * | −4.50 |
| S | 231.15 | 169.23 | 1.37 | 7.37 | 243.03 | 185.77 | 1.31 | 6.40 | 157.63 | 192.06 | 0.82 | −2.26 | |
| Foliar Total Phenol. | C | 16,28 | 16,28 | 1.00 | 0 | 18.98 | 22.23 | 0.85 * | −0.71 | 20.95 | 15.45 | 1.36 * | 2.40 |
| S | 19.91 | 19.58 | 1.02 | 0.097 | 24.34 | 28.17 | 0.86 | −0.74 | 25.18 | 19.28 | 1.31 * | 1.59 | |
| Foliar Total Flav. | C | 4.21 | 4.58 | 0.92 | −3.05 | 5.00 | 5.82 | 0.86 | −0.73 | 5.55 | 4.29 | 1.30 * | 3.06 |
| S | 7.06 | 7.00 | 1.01 | 0.04 | 9.38 | 10.01 | 0.94 | −0.43 | 9.57 | 7.04 | 1.36 * | 1.68 | |
Note: | |||||||||||||
| 0.2 | 1 | 1.8 | |||||||||||
| Species | Accession | Germplasm Code | Origin | Genetic Pool | Hybrids |
|---|---|---|---|---|---|
| S. melongena | MEL1 | BBS-118/B | Côte d’Ivoire | ||
| S. insanum | INS1 | SLKINS-1 | Sri Lanka | Primary | MEL1 × INS1 |
| S. insanum | INS2 | SLKINS-2 | Sri Lanka | Primary | MEL1 × INS2 |
| S. incanum | INC1 | MM664 | Israel | Secondary | INC1 × MEL1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Orenga, S.; Plazas, M.; Ribera, E.; Pallotti, C.; Boscaiu, M.; Prohens, J.; Vicente, O.; Fita, A. Transgressive Biochemical Response to Water Stress in Interspecific Eggplant Hybrids. Plants 2023, 12, 194. https://doi.org/10.3390/plants12010194
González-Orenga S, Plazas M, Ribera E, Pallotti C, Boscaiu M, Prohens J, Vicente O, Fita A. Transgressive Biochemical Response to Water Stress in Interspecific Eggplant Hybrids. Plants. 2023; 12(1):194. https://doi.org/10.3390/plants12010194
Chicago/Turabian StyleGonzález-Orenga, Sara, Mariola Plazas, Elvira Ribera, Claudia Pallotti, Monica Boscaiu, Jaime Prohens, Oscar Vicente, and Ana Fita. 2023. "Transgressive Biochemical Response to Water Stress in Interspecific Eggplant Hybrids" Plants 12, no. 1: 194. https://doi.org/10.3390/plants12010194