
Floral Characterization of Pomegranate Genotypes to Improve Hybridization Efficiency
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Sex Ratio
2.2. Flower Morphology
2.2.1. Length of Flower
2.2.2. Heterostyly
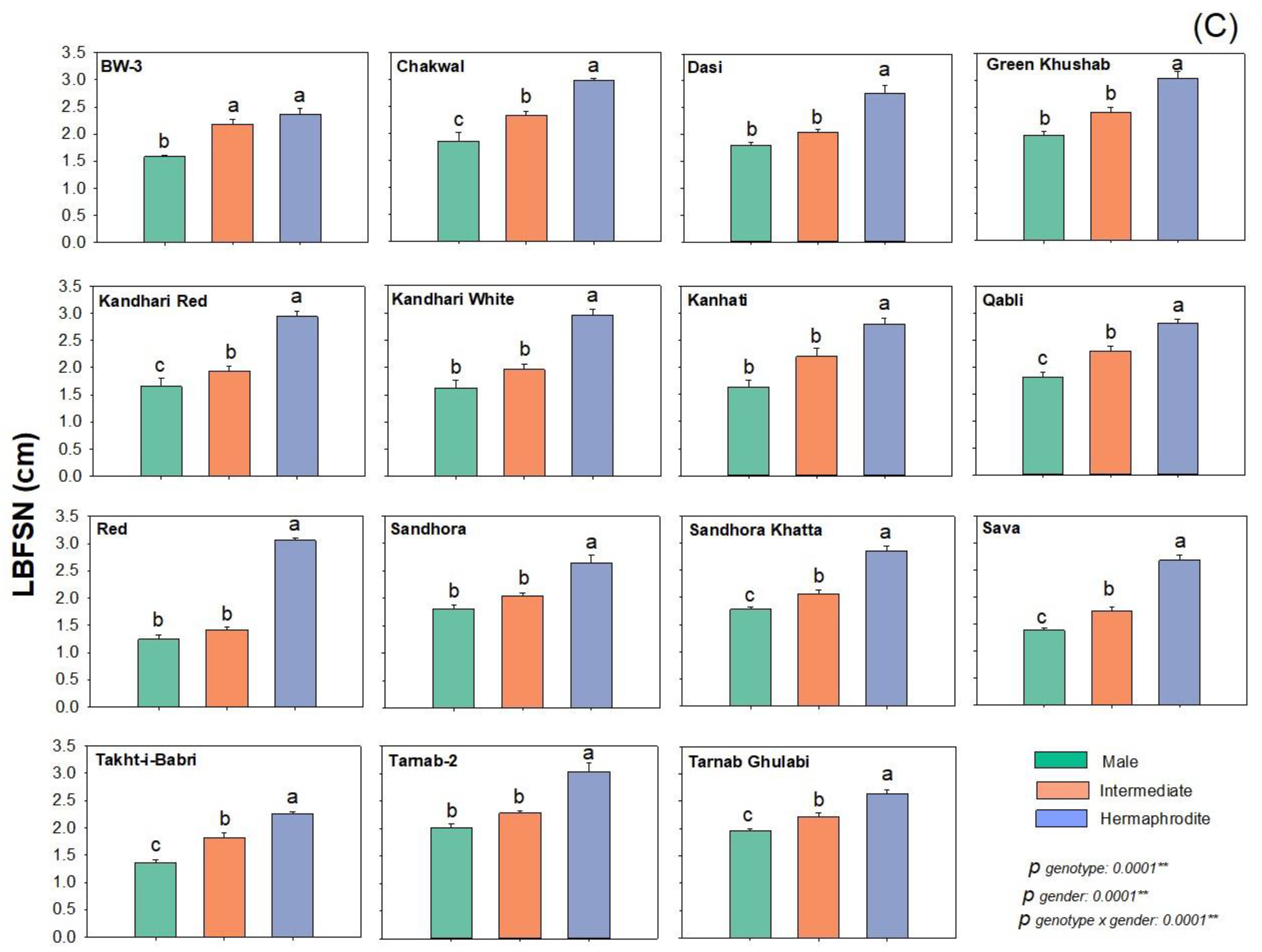
2.3. Flower Width
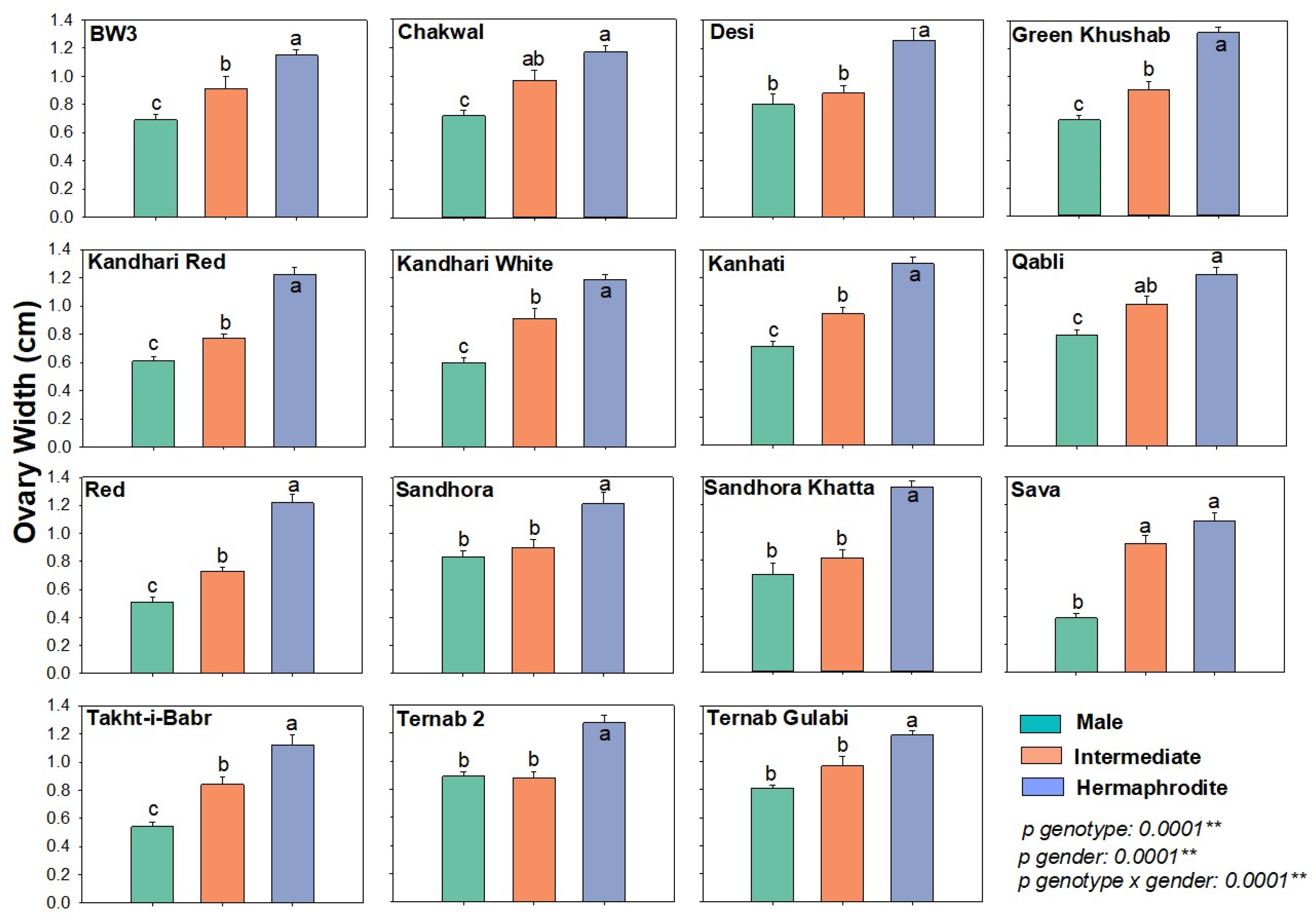
Ovary Width
2.4. Cross-Pollination
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growing Conditions
4.2. Flower Morphology
4.3. Cross Breeding Pollination and Fertization
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pike, S.M. Review of The Narcissus and the Pomegranate: An Archaeology of the Homeric Hymn to Demeter by Ann Suter. Pomegranate Int. J. Pagan. Stud. 2007, 7, 176. [Google Scholar] [CrossRef]
- Taylor, S.H. Pharmacological and Therapeutic Properties of an Ideal β-Blocker. J. Int. Med. Res. 1988, 16, 231–235. [Google Scholar]
- Ross, S.M. Pomegranate: Its Role in Cardiovascular Health. Holist. Nurs. Pract. 2009, 23, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Karimi, M.; Sadeghi, R.; Kokini, J. Pomegranate as a Promising Opportunity in Medicine and Nanotechnology. Trends Food Sci. Technol. 2017, 69, 59–73. [Google Scholar] [CrossRef]
- Ikram, S.; Shafqat, W.; Qureshi, M.A.; Din, S.U.; Rehman, S.U.; Mehmood, A.; Sajjad, Y.; Nafees, M. Causes and Control of Fruit Cracking in Pomegranate: A Review. J. Glob. Innov. Agric. Soc. Sci. 2020, 8, 183–190. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Henning, S.M.; Niu, Y.; Zhang, Y.; Nair, M.G.; Heber, D. In Vitro Antiproliferative, Apoptotic and Antioxidant Activities of Punicalagin, Ellagic Acid and a Total Pomegranate Tannin Extract Are Enhanced in Combination with Other Polyphenols as Found in Pomegranate Juice. J. Nutr. Biochem. 2005, 16, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Longtin, R. The Pomegranate: Nature’s Power Fruit? J. Natl. Cancer Inst. 2003, 95, 346–348. [Google Scholar] [CrossRef]
- Verma, N.; Mohanty, A.; Lal, A. Pomegranate Genetic Resources and Germplasm Conservation: A Review. Fruit Veg. Cereal Sci. Biotechnol. 2010, 4, 120–125. [Google Scholar]
- Jalikop, S.H. Pomegranate Breeding. Fruit, Veg. Cereal Sci. Biothec. 2010, 4, 26–34. [Google Scholar]
- Holland, D.; Bar-Ya’akov, I. Pomegranate (Punica Granatum L.) Breeding. In Advances in Plant Breeding Strategies: Fruits; Springer: Cham, Switzerland, 2018; Volume 3, ISBN 9783319919447. [Google Scholar]
- Holland, D.; Hatib, K.; Bar-ya, I. Pomegranate: Botany, Horticulture, Breeding. Hortic. Rev. (Am. Soc. Hortic. Sci). 2009, 35, 127–192. [Google Scholar]
- Wetzstein, H.Y.; Ravid, N.; Wilkins, E.; Martinelli, A.P. A Morphological and Histological Characterization of Bisexual and Male Flower Types in Pomegranate. J. Am. Soc. Hortic. Sci. 2011, 136, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Dhinesh Babu, K. Floral Biology of Pomegranate (Punica granatum L.). Glob. Sci. Books 2010, 4, 45–50. [Google Scholar]
- Teixeira da Silva, J.A.; Rana, T.S.; Narzary, D.; Verma, N.; Meshram, D.T.; Ranade, S.A. Pomegranate Biology and Biotechnology: A Review. Sci. Hortic. (Amsterdam). 2013, 160, 85–107. [Google Scholar] [CrossRef]
- Chaudhari, S.; Desai, U. Effect of Plant Growth-Regulators on Flower Sex in Pomegranate (Punica granatum). Indian J. Agric. Sci. 1993, 63, 34–35. [Google Scholar]
- Wetzstein, H.Y.; Yi, W.; Porter, J.A.; Ravid, N. Flower Position and Size Impact Ovule Number per Flower, Fruitset, and Fruit Size in Pomegranate. J. Am. Soc. Hortic. Sci. 2013, 138, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Wessinger, C.A.; Hileman, L.C. Parallelism in Flower Evolution and Development; Annual Reviews: Palo Alto, CA, USA, 2020. [Google Scholar]
- Ramírez-Rosas, K.; Aguirre-Jaimes, A.; Ramírez-Morillo, I.M.; García-Franco, J.G. Floral Biology and Potential Hybridization of Three Sympatric Epiphytic Bromeliads in Veracruz, Mexico. Plant Species Biol. 2020, 35, 197–209. [Google Scholar] [CrossRef]
- Soares, T.L.; de Jesus, O.N.; de Souza, E.H.; de Oliveira, E.J. Reproductive Biology and Pollen–Pistil Interactions in Passiflora Species with Ornamental Potential. Sci. Hortic. (Amsterdam) 2015, 197, 339–349. [Google Scholar] [CrossRef]
- Zhang, L.; Ampatzidis, Y.; Whiting, M.D. Sweet Cherry Floral Organ Size Varies with Genotype and Temperature. Sci. Hortic. (Amsterdam) 2015, 182, 156–164. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.; Li, H.; Niu, J.; Xue, H.; Liu, B.; Wang, Q.; Luo, X.; Zhang, F.; Zhao, D.; et al. Transcriptomic Analysis Reveals Candidate Genes for Female Sterility in Pomegranate Flowers. Front. Plant Sci. 2017, 8, 1430. [Google Scholar] [CrossRef] [Green Version]
- Bawa, K.S.; Webb, C.J. Floral Variation and Sexual Differentiation in Muntingia Calabura (Elaeocarpaceae), a Species with Hermaphrodite Flowers. Evolution (N. Y.) 1983, 37, 1271. [Google Scholar] [CrossRef]
- Freeman, D.C.; McArthur, E.D.; Harper, K.T.; Blauer, A.C. Influence of Environment on the Floral Sex Ratio of Monoecious Plants. Evolution (N. Y.) 1981, 35, 194. [Google Scholar] [CrossRef]
- Gokbayrak, Z.; Engin, H. Influence of Plant Growth Regulators on Sex Differentiation and Floral Characteristics of Pomegranate Flowers: A Case of Brassinosteroids. Acta Soc. Bot. Pol. 2018, 87, 3589. [Google Scholar] [CrossRef]
- Melgarejo, P.; Legua, P.; Martinez, M.; Martinez, J.J. Contribution to a Better Knowledge of the Quality of Pomegranate Pollen (Punica granatum L.). Options Méditerranéennes Série A Séminaires Méditerranéens 2000, 115–121. [Google Scholar]
- Solomon, B.P. Environmentally Influenced Changes in Sex Expression in an Andromonoecious Plant. Ecology 1985, 66, 1321–1332. [Google Scholar] [CrossRef]
- Sutherland, S. Floral Sex Ratios, Fruit-Set, and Resource Allocation in Plants. Ecology 1986, 67, 991–1001. [Google Scholar] [CrossRef]
- Albal, A.; Azad, G.; Shrotri, S.; Gowda, V. When to Be a Male? Role of Resource-Limitation and Pollinators in Determining Floral Sex in an Andromonoecious Spiderwort. bioRxiv 2020. [Google Scholar] [CrossRef]
- Engin, H.; Gökbayrak, Z. Effects of Plant Growth Regulators on Sex Expression and Flower Development in Pomegranates. Erwerbs-Obstbau 2019, 61, 23–27. [Google Scholar] [CrossRef]
- Din, S.; Wani, R.A.; Nisar, F.; Farwah, S.; Rizvi, S.; Wani, T.F.; Nisar, S. Fruit Set and Development: Pre-Requisites and Enhancement in Temperate Fruit Crops. J. Pharmacogn. Phytochem. 2019, 8, 1203–1216. [Google Scholar]
- Muñoz-Sanz, J.V.; Zuriaga, E.; Cruz-García, F.; McClure, B.; Romero, C. Self-(in) Compatibility Systems: Target Traits for Crop-Production, Plant Breeding, and Biotechnology. Front. Plant Sci. 2020, 11, 195. [Google Scholar] [CrossRef] [Green Version]
- Martíneza, J.J.; Melgarejo, P.; Hernández, F.; Legua, P. Pollen-Pistil Affinity of Eight New Pomegranate Clones (Punica granatum L.). In International Symposium on Pomegranate and Minor Mediterranean Fruits; ISHS: Leuven, Belgium, 2009; Volume 818, pp. 175–180. [Google Scholar]
- Karimi, H.R.; Mirdehghan, S.H. Effects of Self, Open, and Supplementary Pollination on Growth Pattern and Characteristics of Pomegranate Fruit. Int. J. Fruit Sci. 2015, 15, 382–391. [Google Scholar] [CrossRef]
- Gharaghani, A.; Ghasemi Soloklui, A.A.; Oraguzie, N.; Zare, D. Pollen Source Influences Fruit Quality, Aril Properties, and Seed Characteristics in Pomegranate. Int. J. Fruit Sci. 2017, 17, 333–348. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, H.; Wang, Y.; Zhang, X.; Zhao, X.; Yuan, Z. Genome-Wide Identification and Expression Analysis of MIKC-Type MADS-Box Gene Family in Punica Granatum L. Agronomy 2020, 10, 1197. [Google Scholar] [CrossRef]
- Solomakhin, A.A.; Blanke, M.M. Mechanical Flower Thinning Improves the Fruit Quality of Apples. J. Sci. Food Agric. 2010, 90, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Link, H. Significance of Flower and Fruit Thinning on Fruit Quality. Plant Growth Regul. 2000, 31, 17–26. [Google Scholar] [CrossRef]
- Byers, R.E.; Costa, G.; Vizzotto, G. Flower and Fruit Thinning of Peach and Other Prunus. Hortic. Rev. (Am. Soc. Hortic. Sci.) 2010, 28, 351–392. [Google Scholar] [CrossRef]
- Nafees, M.; Jafar Jaskani, M.; Ahmad, I.; Ashraf, I.; Maqsood, A.; Ahmar, S.; Azam, M.; Hussain, S.; Hanif, A.; Chen, J.-T. Biochemical Analysis of Organic Acids and Soluble Sugars in Wild and Cultivated Pomegranate Germplasm Based in Pakistan. Plants 2020, 9, 493. [Google Scholar] [CrossRef]
- Ali, T.; Nafees, M.; Maqsood, A.; Naqvi, S.A.; Shahzad, U.; Haider, M.S.; Aslam, M.N.; Shafqat, W.; Hameed, M.; Khan, I.A. Contribution of Root Anatomical Characteristics in Fruit Profile of Pomegranate Genotypes to Expand Production Area in Pakistan. Agronomy 2020, 10, 810. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters (cm) | Flower Types | |||||
|---|---|---|---|---|---|---|
| Male | Intermediate | Hermaphrodite | ||||
| Range | Median | Range | Median | Range | Median | |
| Complete flower length | 2.72–3.94 | 3.58 | 3.24–4.82 | 4.18 | 4.44–5.88 | 5.12 |
| Length from the base of flowers to sepal notch | 1.24–2.02 | 1.78 | 1.42–2.39 | 2.06 | 2.25–3.06 | 2.83 |
| Length from the base of flowers to the tip of sepals | 2.04–2.94 | 2.64 | 2.56–3.54 | 3.08 | 2.77–4.26 | 3.86 |
| Length of stigma+ style+ stylopodium | 0.38–0.72 | 0.59 | 0.66–1.04 | 0.9 | 1.13–2.12 | 1.7 |
| Length of stigma+ style | 0.19–0.43 | 0.31 | 0.36–0.75 | 0.5 | 0.76–1.39 | 1.18 |
| Ovary width | 0.39–0.89 | 0.7 | 0.73–1.01 | 0.91 | 1.08–1.33 | 1.22 |
| Cross Combination (Female × Male) | Fruit Set (%) | Cross Combination (Female × Male) | Fruit Set (%) |
|---|---|---|---|
| Kandhari White × Chakwal | 95.6 | BW3 × Tarnab Ghulabi | 58.3 |
| Tarnab Ghulabi × Tarnab 2 | 92.8 | Sandhora × Sandhora Khatta | 56.6 |
| Ternab 2 × Red | 91.8 | Takht-i-Babri × Green Khushab | 56.3 |
| Ternab 2 × Kandhari White | 89.3 | Red × BW3 | 51.2 |
| Ternab 2 × Sava | 87.2 | Sandhora Khatta × BW3 | 47.8 |
| Kandhari White × Tarnab Ghulabi | 85.7 | Red × Takht-i-Babri | 46.6 |
| Ternab 2 × Takht-i-Babri | 85.4 | Tarnab Ghulabi × BW3 | 45.7 |
| Kandhari White × Kandhari Red | 84.3 | Sandhora × Ternab 2 | 45.7 |
| Kandhari White × Sava | 84.1 | Qabli × Takht-i-Babri | 45.5 |
| Kandhari White × Qabli | 82.5 | BW3 × Red | 43.8 |
| Kandhari Red × Qabli | 82.4 | Takht-i-Babri × Kandhari White | 41.3 |
| Sandhora Khatta × Sava | 82.2 | Takht-i-Babri × Sava | 41.2 |
| Tarnab Ghulabi × Sava | 81.7 | Takht-i-Babri × Kandhari Red | 36.4 |
| Ternab 2 × Sandhora | 79.8 | Sava × Sandhora | 34.6 |
| Kandhari White × Ternab 2 | 77.4 | Red × Kandhari Red | 32.1 |
| Kandhari White × Ternab Ghulabi | 75.6 | BW3 × Kandhari White | 32.1 |
| Green Khushab × Takht-i-Babri | 74.5 | Takht-i-Babri × Sandhora | 31.3 |
| Qabli × Kandhari White | 67.5 | BW3 × Kandhari Red | 29.4 |
| Qabli × Khatta Khushab | 64.3 | Red × Kandhari White | 28.7 |
| Qabli × Kandhari Red | 59.5 | BW3 × Ternab 2 | 23.6 |
| Genotype Name | TSS (Brix) | TA (g/L) | Fruit Rind Color | Rind Thickness (mm) | Aril Color | WPI Seed Hardiness (%) | Seed Area (mm2) | Fruit Weight (g) |
|---|---|---|---|---|---|---|---|---|
| Tarnab Ghulabi | 14.2 | 0.91 | Pinkish red | 4.47 | Red | 3.99 | 7.89 | ±289 |
| Kanhati | 13.2 | 0.15 | Yellowish green | 4.09 | Pinkish | 6.11 | 10.87 | ±70 |
| Tarnab 2 | N/A | |||||||
| Qabli | 16.8 | 0.44 | Reddish-green | 3.34 | Pinkish-white | 4.89 | 9.87 | ±220 |
| Green Khushab | 13.2 | 0.15 | Pinkish-green | 3.99 | Pinkish-white | 3.22 | 9.17 | ±170 |
| Sandhora | 12.9 | 0.11 | Pinkish-green | 3.77 | Pinkish-red | 6.78 | 9.91 | ±221 |
| Sandhora Khatta | 11.2 | 0.25 | Pinkish-yellow | 3.66 | Pinkish-white | 4.78 | 8.96 | ±184 |
| Sava | 15.9 | 0.09 | Yellowish | 3.22 | White-pink | 3.89 | 11.65 | ±278 |
| Takht-i-Babri | 13 | 0.23 | Reddish | 2.48 | Pinkish-white | 13.65 | 8.97 | ±46 |
| Chakwal | 13.8 | 0.15 | Reddish | 2.01 | white | 15.67 | 9.22 | ±40 |
| Kandhari red | 14.9 | 0.17 | Red | 2.87 | Red | 3.59 | 10.78 | ±240 |
| Kandhari White | 15.1 | 1.4 | Pinkish-red | 2.99 | Red | 5.33 | 8.98 | ±220 |
| Dasi | 15.7 | 0.12 | Reddish-pink | 2.24 | Pinkish-white | 5.78 | 9.87 | ±118 |
| Red | 14.5 | 0.15 | 3.77 | 4.67 | 9.78 | ±167 | ||
| BW3 | N/A | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikram, S.; Shafqat, W.; Rehman, S.U.; Qureshi, M.A.; Din, S.u.; Ikram, S.; Nafees, M.; Jaskani, M.J. Floral Characterization of Pomegranate Genotypes to Improve Hybridization Efficiency. Plants 2023, 12, 165. https://doi.org/10.3390/plants12010165
Ikram S, Shafqat W, Rehman SU, Qureshi MA, Din Su, Ikram S, Nafees M, Jaskani MJ. Floral Characterization of Pomegranate Genotypes to Improve Hybridization Efficiency. Plants. 2023; 12(1):165. https://doi.org/10.3390/plants12010165
Chicago/Turabian StyleIkram, Sufian, Waqar Shafqat, Sami Ur Rehman, Muhammad Ahsan Qureshi, Safeer ud Din, Salman Ikram, Muhammad Nafees, and Muhammad Jafar Jaskani. 2023. "Floral Characterization of Pomegranate Genotypes to Improve Hybridization Efficiency" Plants 12, no. 1: 165. https://doi.org/10.3390/plants12010165