
Can Microbial Consortium Applications Affect Yield and Quality of Conventionally Managed Processing Tomato?

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Yield and Its Components
2.2. Chemical and Technological Traits
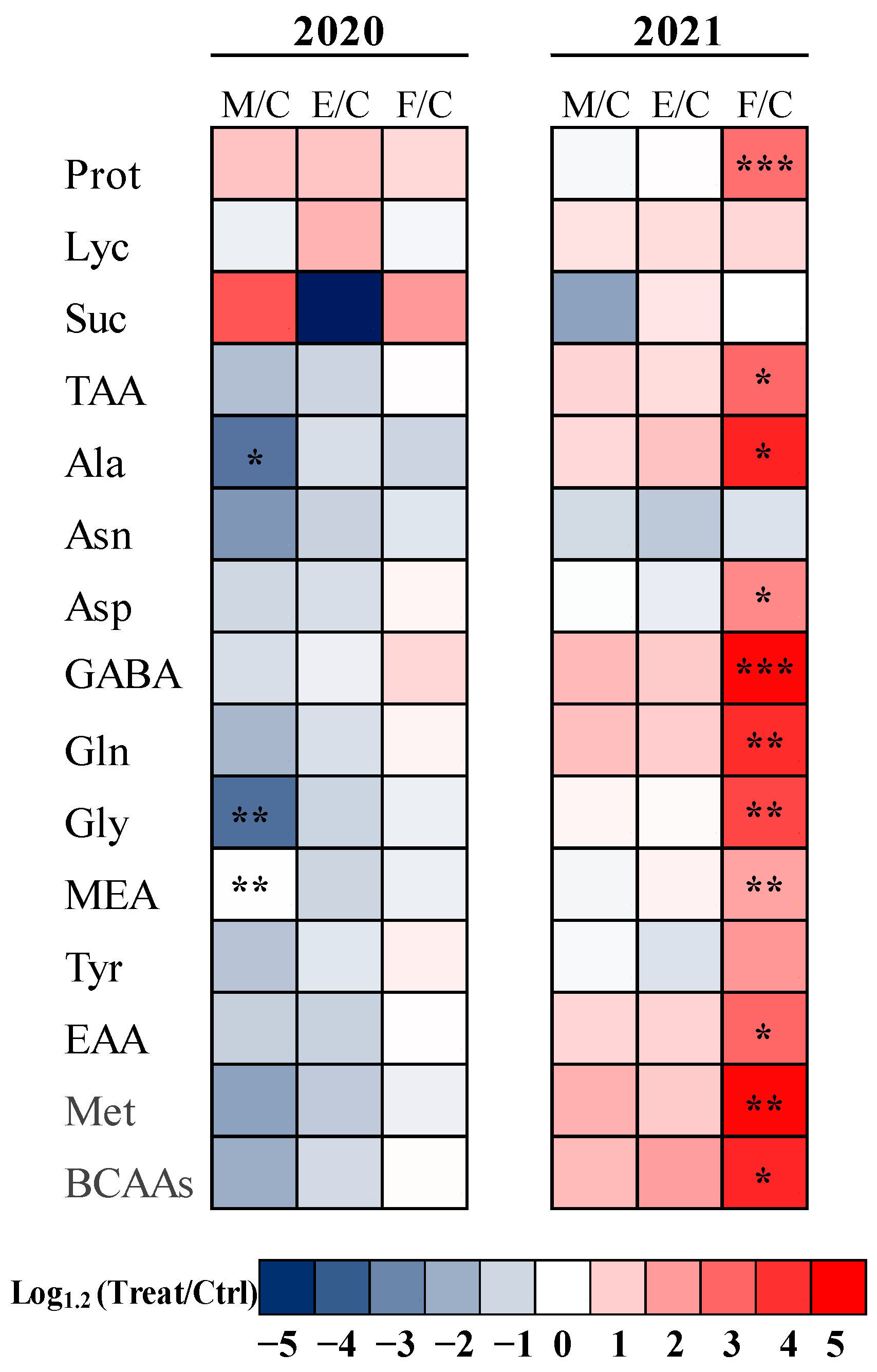
2.3. Primary Metabolite Content
3. Discussion
3.1. Effect of Different Microbial Consortia on Yield and Technological Quality of Tomato Fruits
3.2. Effect of Different Microbial Consortia on Biochemical Properties of Tomato Fruits
4. Materials and Methods
4.1. Location
4.2. Meteorological Data
4.3. Experimental Design and Microbial-Based Treatments
4.4. Yield and Merceological Assessment
4.5. Technological Characteristics
4.6. Fruit Metabolic Profiling
4.7. Starch and Soluble Sugars Analysis
4.8. Polyphenols and Lycopene Analysis
4.9. Soluble Proteins and Free Amino Acid Analysis
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gerszberg, A.; Hnatuszko-Konka, K.; Kowalczyk, T.; Kononowicz, A.K. Tomato (Solanum lycopersicum L.) in the Service of Biotechnology. Plant Cell Tissue Organ Cult. 2015, 120, 881–902. [Google Scholar] [CrossRef] [Green Version]
- FAO. World Food and Agriculture—Statistical Yearbook 2020; FAO: Rome, Italy, 2020; ISBN 978-92-5-133394-5. [Google Scholar]
- Collins, E.J.; Bowyer, C.; Tsouza, A.; Chopra, M. Tomatoes: An Extensive Review of the Associated Health Impacts of Tomatoes and Factors That Can Affect Their Cultivation. Biology 2022, 11, 239. [Google Scholar] [CrossRef] [PubMed]
- Lima, G.P.P.; Gómez, H.A.G.; Seabra Junior, S.; Maraschin, M.; Tecchio, M.A.; Borges, C.V. Functional and Nutraceutical Compounds of Tomatoes as Affected by Agronomic Practices, Postharvest Management, and Processing Methods: A Mini Review. Front. Nutr. 2022, 9, 584. [Google Scholar] [CrossRef] [PubMed]
- Nasir, M.U.; Hussain, S.; Jabbar, S. Tomato Processing, Lycopene and Health Benefits: A Review. Sci. Lett. 2015, 3, 1–5. [Google Scholar]
- Agarwal, S.; Rao, A.V. Tomato Lycopene and Its Role in Human Health and Chronic Diseases. Can. Med. Assoc. J. 2000, 163, 739. [Google Scholar]
- Gramazio, P.; Takayama, M.; Ezura, H. Challenges and Prospects of New Plant Breeding Techniques for GABA Improvement in Crops: Tomato as an Example. Front. Plant Sci 2020, 11, 577980. [Google Scholar] [CrossRef]
- Distefano, M.; Arena, E.; Mauro, R.P.; Brighina, S.; Leonardi, C.; Fallico, B.; Giuffrida, F. Effects of Genotype, Storage Temperature and Time on Quality and Compositional Traits of Cherry Tomato. Foods 2020, 9, 1729. [Google Scholar] [CrossRef]
- Dumas, Y.; Dadomo, M.; di Lucca, G.; Grolier, P. Effects of Environmental Factors and Agricultural Techniques on Antioxidantcontent of Tomatoes. J. Sci. Food Agric. 2003, 83, 369–382. [Google Scholar] [CrossRef]
- Parisi, M.; Burato, A.; Pentangelo, A.; Ronga, D. Towards the Optimal Mineral N Fertilization for Improving Peeled Tomato Quality Grown in Southern Italy. Horticulturae 2022, 8, 697. [Google Scholar] [CrossRef]
- Godlewska, K.; Ronga, D.; Michalak, I. Plant Extracts—Importance in Sustainable Agriculture. Ital. J. Agron. 2021, 16. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Boopathi, T.; Manivannan, P. Comprehensive Assessment of Ameliorative Effects of AMF in Alleviating Abiotic Stress in Tomato Plants. J. Fungi 2021, 7, 303. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial Inoculants: Reviewing the Past, Discussing the Present and Previewing an Outstanding Future for the Use of Beneficial Bacteria in Agriculture. AMB Express 2019, 9, 205. [Google Scholar] [CrossRef]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Pseudomonads Improve Yield, Quality and Nutritional Value of Tomato: A Field Study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef]
- Pasković, I.; Soldo, B.; Goreta Ban, S.; Radić, T.; Lukić, M.; Urlić, B.; Mimica, M.; Brkić Bubola, K.; Colla, G.; Rouphael, Y.; et al. Fruit Quality and Volatile Compound Composition of Processing Tomato as Affected by Fertilisation Practices and Arbuscular Mycorrhizal Fungi Application. Food Chem. 2021, 359, 129961. [Google Scholar] [CrossRef]
- Cardarelli, M.; Coppa, E.; Rouphael, Y.; Mariotti, R.; Bonini, P.; Colla, G. Combined Applications of Endophytic Fungi and Vegetal Extracts Improve Crop Productivity and Economic Profitability in Processing Tomato. Italus Hortus 2020, 27, 96–105. [Google Scholar] [CrossRef]
- Saia, S.; Aissa, E.; Luziatelli, F.; Ruzzi, M.; Colla, G.; Ficca, A.G.; Cardarelli, M.; Rouphael, Y. Growth-Promoting Bacteria and Arbuscular Mycorrhizal Fungi Differentially Benefit Tomato and Corn Depending upon the Supplied Form of Phosphorus. Mycorrhiza 2020, 30, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; de Pascale, S.; Colla, G. Implications of Microbial and Non-Microbial Biostimulatory Action on the Quality of Leafy and Fruit Vegetables. Acta Hortic. 2020, 1268, 13–17. [Google Scholar] [CrossRef]
- Cammarano, D.; Ronga, D.; di Mola, I.; Mori, M.; Parisi, M. Impact of Climate Change on Water and Nitrogen Use Efficiencies of Processing Tomato Cultivated in Italy. Agric. Water Manag. 2020, 241, 106336. [Google Scholar] [CrossRef]
- Cammarano, D.; Jamshidi, S.; Hoogenboom, G.; Ruane, A.C.; Niyogi, D.; Ronga, D. Processing Tomato Production Is Expected to Decrease by 2050 Due to the Projected Increase in Temperature. Nat. Food 2022, 3, 437–444. [Google Scholar] [CrossRef]
- Caradonia, F.; Ronga, D.; Catellani, M.; Giaretta Azevedo, C.V.; Terrazas, R.A.; Robertson-Albertyn, S.; Francia, E.; Bulgarelli, D. Nitrogen Fertilizers Shape the Composition and Predicted Functions of the Microbiota of Field-Grown Tomato Plants. Phytobiomes J. 2019, 3, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Singh, A.K.; Singh, P.P.; Kumar, A. Interaction of Plant Growth Promoting Bacteria with Tomato under Abiotic Stress: A Review. Agric. Ecosyst. Environ. 2018, 267, 129–140. [Google Scholar] [CrossRef]
- Carillo, P.; Kyratzis, A.; Kyriacou, M.C.; Dell’Aversana, E.; Fusco, G.M.; Corrado, G.; Rouphael, Y. Biostimulatory Action of Arbuscular Mycorrhizal Fungi Enhances Productivity, Functional and Sensory Quality in ‘Piennolo Del Vesuvio’ Cherry Tomato Landraces. Agronomy 2020, 10, 911. [Google Scholar] [CrossRef]
- Kalozoumis, P.; Savvas, D.; Aliferis, K.; Ntatsi, G.; Marakis, G.; Simou, E.; Tampakaki, A.; Karapanos, I. Impact of Plant Growth-Promoting Rhizobacteria Inoculation and Grafting on Tolerance of Tomato to Combined Water and Nutrient Stress Assessed via Metabolomics Analysis. Front. Plant Sci. 2021, 12, 814. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Read, D. Mycorrhizal Symbiosis; Elsevier Ltd.: New York, NY, USA, 2008; ISBN 9780123705266. [Google Scholar]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–Plant–Pathogen Interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Lifshitz, R.; Zablotowicz, R.M. Free-Living Bacterial Inocula for Enhancing Crop Productivity. Trends Biotechnol. 1989, 7, 39–44. [Google Scholar] [CrossRef]
- Antoun, H.; Prévost, D. Ecology of Plant Growth Promoting Rhizobacteria. In PGPR: Biocontrol and Biofertilization; Springer: Dordrecht, The Netherlands, 2006; pp. 1–38. [Google Scholar] [CrossRef]
- Li, J.; van Gerrewey, T.; Geelen, D. A Meta-Analysis of Biostimulant Yield Effectiveness in Field Trials. Front. Plant Sci. 2022, 13, 1106. [Google Scholar] [CrossRef]
- Fusco, G.M.; Nicastro, R.; Rouphael, Y.; Carillo, P. The Effects of the Microbial Biostimulants Approved by EU Regulation 2019/1009 on Yield and Quality of Vegetable Crops. Foods 2022, 11, 2656. [Google Scholar] [CrossRef]
- Caradonia, F.; Ronga, D.; Flore, A.; Barbieri, R.; Moulin, L.; Terzi, V.; Francia, E. Biostimulants and Cherry Rootstock Increased Tomato Fruit Yield and Quality in Sustainable Farming Systems. Ital. J. Agron. 2020, 15, 121–131. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Haruta, S.; Yamamoto, K. Model Microbial Consortia as Tools for Understanding Complex Microbial Communities. Curr. Genom. 2018, 19, 723–733. [Google Scholar] [CrossRef]
- Ram, R.M.; Singh, H.B. Microbial Consortium in Biological Control: An Explicit Example of Teamwork below Ground. J. Eco-Friendly Agric. 2018, 13, 1–12. [Google Scholar]
- Bhatia, S.K.; Bhatia, R.K.; Choi, Y.K.; Kan, E.; Kim, Y.G.; Yang, Y.H. Biotechnological Potential of Microbial Consortia and Future Perspectives. Crit. Rev. Biotechnol. 2018, 38, 1209–1229. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Toward a Sustainable Agriculture through Plant Biostimulants: From Experimental Data to Practical Applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Ronga, D.; Caradonia, F.; Francia, E.; Morcia, C.; Rizza, F.; Badeck, F.W.; Ghizzoni, R.; Terzi, V. Interaction of Tomato Genotypes and Arbuscular Mycorrhizal Fungi under Reduced Irrigation. Horticulturae 2019, 5, 79. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Casa, R.; lo Cascio, B.; Saccardo, F.; Temperini, O.; Leoni, C. Responses of Processing Tomato to Water Regime and Fertilization in Central Italy. Acta Hortic. 1999, 487, 531–535. [Google Scholar] [CrossRef]
- Di Cesare, L.F.; Migliori, C.; Ferrari, V.; Parisi, M.; Campanelli, G.; Candido, V.; Perrone, D. Effects of Irrigation-Fertilization and Irrigation-Mycorrhization on the Alimentary and Nutraceutical Properties of Tomatoes. In Irrigation Systems and Practices in Challenging Environments; InTech: Rijeka, Croatia, 2012; pp. 207–232. ISBN 978-953-51-0420-9. [Google Scholar]
- Patanè, C.; Saita, A. Biomass, Fruit Yield, Water Productivity and Quality Response of Processing Tomato to Plant Density and Deficit Irrigation under a Semi-Arid Mediterranean Climate. Crop Pasture Sci. 2015, 66, 224–234. [Google Scholar] [CrossRef]
- Patanè, C.; Tringali, S.; Sortino, O. Effects of Deficit Irrigation on Biomass, Yield, Water Productivity and Fruit Quality of Processing Tomato under Semi-Arid Mediterranean Climate Conditions. Sci. Hortic. 2011, 129, 590–596. [Google Scholar] [CrossRef]
- Tahiri, A.I.; Meddich, A.; Raklami, A.; Alahmad, A.; Bechtaoui, N.; Anli, M.; Göttfert, M.; Heulin, T.; Achouak, W.; Oufdou, K. Assessing the Potential Role of Compost, PGPR, and AMF in Improving Tomato Plant Growth, Yield, Fruit Quality, and Water Stress Tolerance. J. Soil Sci. Plant Nutr. 2021, 22, 743–764. [Google Scholar] [CrossRef]
- Turhan, A. Interactive Effects of Boron Stress and Mycorrhizal (AMF) Treatments on Tomato Growth, Yield, Leaf Chlorophyll and Boron Accumulation, and Fruit Characteristics. Arch. Agron. Soil Sci. 2021, 67, 1974–1985. [Google Scholar] [CrossRef]
- Ronga, D.; Parisi, M.; Pentangelo, A.; Mori, M.; Mola, I. di Effects of Nitrogen Management on Biomass Production and Dry Matter Distribution of Processing Tomato Cropped in Southern Italy. Agronomy 2019, 9, 855. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; dell’Aversana, E.; D’Amelia, L.; Colla, G.; Caruso, G.; de Pascale, S.; Rouphael, Y. Sensory and Functional Quality Characterization of Protected Designation of Origin ‘Piennolo Del Vesuvio’ Cherry Tomato Landraces from Campania-Italy. Food Chem. 2019, 292, 166–175. [Google Scholar] [CrossRef]
- Thies, F.; Mills, L.M.; Moir, S.; Masson, L.F. Cardiovascular Benefits of Lycopene: Fantasy or Reality? Proc. Nutr. Soc. 2017, 76, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Frusciante, L.; Carli, P.; Ercolano, M.R.; Pernice, R.; di Matteo, A.; Fogliano, V.; Pellegrini, N. Antioxidant Nutritional Quality of Tomato. Mol. Nutr. Food Res. 2007, 51, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Sipos, L.; Orbán, C.; Bálint, I.; Csambalik, L.; Divéky-Ertsey, A.; Gere, A. Colour Parameters as Indicators of Lycopene and Antioxidant Activity Traits of Cherry Tomatoes (Solanum lycopersicum L.). Eur. Food Res. Technol. 2017, 243, 1533–1543. [Google Scholar] [CrossRef]
- Giovannetti, M.; Avio, L.; Barale, R.; Ceccarelli, N.; Cristofani, R.; Iezzi, A.; Mignolli, F.; Picciarelli, P.; Pinto, B.; Reali, D.; et al. Nutraceutical Value and Safety of Tomato Fruits Produced by Mycorrhizal Plants. Br. J. Nutr. 2012, 107, 242–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nzanza, B.; Marais, D.; Soundy, P. Yield and Nutrient Content of Tomato (Solanum lycopersicum L.) as Influenced by Trichoderma Harzianum and Glomus Mosseae Inoculation. Sci. Hortic. 2012, 144, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Soteriou, G.A.; Kyriacou, M.C.; Giordano, M.; Raimondi, G.; Napolitano, F.; di Stasio, E.; di Mola, I.; Mori, M.; Rouphael, Y. Regulated Salinity Eustress in a Floating Hydroponic Module of Sequentially Harvested Lettuce Modulates Phytochemical Constitution, Plant Resilience, and Post-Harvest Nutraceutical Quality. Agronomy 2021, 11, 1040. [Google Scholar] [CrossRef]
- Thomas, A.; Beena, R.; Laksmi, G.; Soni, K.B.; Alex, S.; Viji, M.M. Changes in Sucrose Metabolic Enzymes to Water Stress in Contrasting Rice Genotypes. Plant Stress 2022, 5, 100088. [Google Scholar] [CrossRef]
- Kowalska, I.; Konieczny, A.; Gąstoł, M.; Sady, W.; Hanus-Fajerska, E. Effect of Mycorrhiza and Phosphorus Content in Nutrient Solution on the Yield and Nutritional Status of Tomato Plants Grown on Rockwool or Coconut Coir. Agric. Food Sci. 2015, 24, 39–51. [Google Scholar] [CrossRef]
- Carillo, P.; Woo, S.L.; Comite, E.; El-nakhel, C.; Rouphael, Y.; Fusco, G.M.; Borzacchiello, A.; Lanzuise, S.; Vinale, F. Application of Trichoderma Harzianum, 6-Pentyl-α-Pyrone and Plant Biopolymer Formulations Modulate Plant Metabolism and Fruit Quality of Plum Tomatoes. Plants 2020, 9, 771. [Google Scholar] [CrossRef]
- Carillo, P. GABA Shunt in Durum Wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Ruiz, R.; Poirot, E.; Flores-Mosquera, M. GABA, a Non-Protein Amino Acid Ubiquitous in Food Matrices. Cogent Food Agric. 2018, 4, 1534323. [Google Scholar] [CrossRef]
- Ngo, D.H.; Vo, T.S. An Updated Review on Pharmaceutical Properties of Gamma-Aminobutyric Acid. Molecules 2019, 24, 2678. [Google Scholar] [CrossRef] [Green Version]
- Tiansawang, K.; Luangpituksa, P.; Varanyanond, W.; Hansawasdi, C. GABA (γ-Aminobutyric Acid) Production, Antioxidant Activity in Some Germinated Dietary Seeds and the Effect of Cooking on Their GABA Content. Food Sci. Technol. 2016, 36, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Parisi, D.; Woodrow, P.; Pontecorvo, G.; Massaro, G.; Annunziata, M.G.; Fuggi, A.; Sulpice, R. Salt-Induced Accumulation of Glycine Betaine Is Inhibited by High Light in Durum Wheat. Funct. Plant Biol. 2011, 38, 139–150. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Measurement of Antioxidant Activity in Food and Biological Systems. ACS Symp. Ser. 2007, 956, 36–66. [Google Scholar] [CrossRef]
- Salvioli, A.; Zouari, I.; Chalot, M.; Bonfante, P. The Arbuscular Mycorrhizal Status Has an Impact on the Transcriptome Profile and Amino Acid Composition of Tomato Fruit. BMC Plant Biol. 2012, 12, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, W.; Secor, J.; Schrader, L.E. Rapid Accumulation of Gamma-Aminobutyric Acid and Alanine in Soybean Leaves in Response to an Abrupt Transfer to Lower Temperature, Darkness, or Mechanical Manipulation. Plant Physiol. 1984, 75, 170–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Fuggi, A. Nitrate Reductase in Durum Wheat Seedlings as Affected by Nitrate Nutrition and Salinity. Funct. Plant Biol. 2005, 32, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.R.; Qiao, S.Y.; Lu, W.Q.; Li, D.F. Effects of Enzyme Supplementation on Performance, Nutrient Digestibility, Gastrointestinal Morphology, and Volatile Fatty Acid Profiles in the Hindgut of Broilers Fed Wheat-Based Diets. Poult. Sci. 2005, 84, 875–881. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Nitrogen Containing Compounds and Adaptation of Plants to Salinity Stress. Biol. Plant. 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Ishizaki, K.; Nunes-Nesi, A.; Larson, T.R.; Tohge, T.; Krahnert, I.; Witt, S.; Obata, T.; Schauer, N.; Graham, I.A.; et al. Identification of the 2-Hydroxyglutarate and Isovaleryl-CoA Dehydrogenases as Alternative Electron Donors Linking Lysine Catabolism to the Electron Transport Chain of Arabidopsis Mitochondria. Plant Cell 2010, 22, 1549–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, V.; Joung, J.G.; Fei, Z.; Jander, G. Interdependence of Threonine, Methionine and Isoleucine Metabolism in Plants: Accumulation and Transcriptional Regulation under Abiotic Stress. Amino Acids 2010, 39, 933–947. [Google Scholar] [CrossRef]
- Obata, T.; Fernie, A.R. The Use of Metabolomics to Dissect Plant Responses to Abiotic Stresses. Cell. Mol. Life Sci. 2012, 69, 3225. [Google Scholar] [CrossRef] [Green Version]
- Iwasa, M.; Kobayashi, Y.; Mifuji-Moroka, R.; Hara, N.; Miyachi, H.; Sugimoto, R.; Tanaka, H.; Fujita, N.; Gabazza, E.C.; Takei, Y. Branched-Chain Amino Acid Supplementation Reduces Oxidative Stress and Prolongs Survival in Rats with Advanced Liver Cirrhosis. PLoS ONE 2013, 8, e70309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korenaga, M.; Nishina, S.; Korenaga, K.; Tomiyama, Y.; Yoshioka, N.; Hara, Y.; Sasaki, Y.; Shimonaka, Y.; Hino, K. Branched-Chain Amino Acids Reduce Hepatic Iron Accumulation and Oxidative Stress in Hepatitis C Virus Polyprotein-Expressing Mice. Liver Int. 2015, 35, 1303–1314. [Google Scholar] [CrossRef] [Green Version]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014. [Google Scholar]
- Hargreaves, G.H.; Samani, Z.A. Reference Crop Evapotranspiration from Temperature. Appl. Eng. Agric. 1985, 1, 96–99. [Google Scholar] [CrossRef]
- Allen, R.; Pereira, L.; Raes, D.; Smith, M. FAO Irrigation and Drainage Paper No. 56; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; Volume 56, pp. 26–40. [Google Scholar]
- Doorenbos, J.; Pruitt, W.O. Crop Water Requirement. FAO Irrigation and Drainage Paper 24; FAO: Rome, Italy, 1977. [Google Scholar]
- Di Cesare, L.F.; Migliori, C.; Viscardi, D.; Parisi, M. Quality of Tomato Fertilized with Nitrogen and Phosphorous. Ital. J. Food Sci. 2010, 22, 186–191. [Google Scholar]
- Parisi, M.; di Dato, F.; Ricci, S.; Mennella, G.; Cardi, T.; Tripodi, P. A Multi-Trait Characterization of the ‘Friariello’ Landrace: A Mediterranean Resource for Sweet Pepper Breeding. Plant Genet. Resour. 2017, 15, 165–176. [Google Scholar] [CrossRef]
- Dell’Aversana, E.; Hessini, K.; Ferchichi, S.; Fusco, G.M.; Woodrow, P.; Ciarmiello, L.F.; Abdelly, C.; Carillo, P. Salinity Duration Differently Modulates Physiological Parameters and Metabolites Profile in Roots of Two Contrasting Barley Genotypes. Plants 2021, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Salbitani, G.; Carillo, P.; di Martino, C.; Bolinesi, F.; Mangoni, O.; Loreto, F.; Carfagna, S. Microalgae Cross-Fertilization: Short-Term Effects of Galdieria Phlegrea Extract on Growth, Photosynthesis and Enzyme Activity of Chlorella Sorokiniana Cells. J. Appl. Phycol. 2022, 34, 1957–1966. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Sadler, G.; Davis, J.; Dezman, D. Rapid Extraction of Lycopene and β-Carotene from Reconstituted Tomato Paste and Pink Grapefruit Homogenates. J. Food Sci. 1990, 55, 1460–1461. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Carillo, P.; Colla, G.; Graziani, G.; di Mola, I.; Mori, M.; Kyriacou, M.C.; Rouphael, Y.; Soteriou, G.A.; et al. Plant-Derived Biostimulants Differentially Modulate Primary and Secondary Metabolites and Improve the Yield Potential of Red and Green Lettuce Cultivars. Agronomy 2022, 12, 1361. [Google Scholar] [CrossRef]


{kind=link}
| RY | GY | MY | TY | RY/TY | GY/TY | MY/TY | FW | SsF | VrF | TDF | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (t ha−1) | (t ha−1) | (t ha−1) | (t ha−1) | (%) | (%) | (%) | (g) | (%) | (%) | (%) | |
| Treatment | |||||||||||
| MIC | 4.61 | 19.68 | 75.80 | 100.10 | 4.67 | 19.35 | 75.98 | 71.45 | 3.00 | 17.17 | 20.17 |
| EKO | 4.70 | 20.94 | 79.80 | 105.43 | 4.65 | 19.19 | 76.16 | 69.06 | 4.67 | 14.00 | 18.67 |
| FID | 5.45 | 24.18 | 82.05 | 111.67 | 4.90 | 21.34 | 73.75 | 71.17 | 4.17 | 15.00 | 19.17 |
| CTRL | 4.97 | 12.93 | 83.18 | 101.08 | 4.93 | 12.49 | 82.58 | 67.96 | 5.83 | 12.50 | 18.33 |
| Year | |||||||||||
| 2020 | 6.13 a | 13.55 b | 80.89 | 100.58 | 6.09 a | 13.02 b | 80.89 a | 68.19 b | 5.83 a | 26.50 a | 32.33 a |
| 2021 | 3.73 b | 25.31 a | 79.52 | 108.56 | 3.48 b | 23.17 a | 73.34 b | 71.63 a | 3.00 b | 2.83 b | 5.83 b |
| Treatment × Year | |||||||||||
| MIC 2020 | 6.26 | 10.85 | 79.42 | 96.54 | 6.45 | 11.15 | 82.40 | 69.54 | 3.00 | 32.33 | 35.33 |
| EKO 2020 | 6.23 | 19.25 | 78.14 | 103.62 | 6.21 | 17.31 | 76.48 | 67.82 | 7.00 | 26.00 | 33.00 |
| FID 2020 | 6.29 | 15.39 | 82.02 | 103.70 | 6.01 | 14.90 | 79.09 | 70.55 | 5.67 | 25.00 | 30.67 |
| CTRL 2020 | 5.74 | 8.72 | 83.98 | 98.45 | 5.70 | 8.70 | 85.60 | 64.86 | 7.67 | 22.67 | 30.33 |
| MIC 2021 | 2.96 | 28.52 | 72.18 | 103.65 | 2.89 | 27.56 | 69.56 | 73.36 | 3.00 | 2.00 | 5.00 |
| EKO 2021 | 3.16 | 22.62 | 81.46 | 107.24 | 3.09 | 21.07 | 75.84 | 70.29 | 2.33 | 2.00 | 4.33 |
| FID 2021 | 4.60 | 32.96 | 82.08 | 119.64 | 3.80 | 27.78 | 68.42 | 71.79 | 2.67 | 5.00 | 7.67 |
| CTRL 2021 | 4.19 | 17.15 | 82.37 | 103.70 | 4.15 | 16.29 | 79.56 | 71.06 | 4.00 | 2.33 | 6.33 |
| Significance | |||||||||||
| Treatment | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Year | * | ** | ns | ns | ** | ** | * | * | * | *** | *** |
| Treatment × Year | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Mean | 4.93 | 19.43 | 80.21 | 104.57 | 4.79 | 18.10 | 77.12 | 69.91 | 4.42 | 14.67 | 19.08 |
| Glc | Fru | Suc | Tsu | Sta | PP | Lyc | pH | TtA | SSC | DM | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (µg g−1) | (µg g−1) | (µg g−1) | (µg g−1) | (µg g−1) | (µg mg−1) | (mg g−1) | (g% Citric Acid) | (°Brix) | (g) | ||
| Treatment | |||||||||||
| MIC | 12.63 | 9.06 | 0.73 a | 22.42 | 6.27 | 0.29 | 6.77 ab | 4.44 | 0.48 | 5.35 | 6.28 |
| EKO | 12.11 | 9.52 | 0.27 b | 21.90 | 6.52 | 0.31 | 7.99 a | 4.52 | 0.44 | 5.33 | 6.33 |
| FID | 12.05 | 9.47 | 0.64 a | 22.15 | 5.98 | 0.30 | 7.04 ab | 4.48 | 0.47 | 5.38 | 6.32 |
| CTRL | 12.08 | 9.88 | 0.48 ab | 22.44 | 6.31 | 0.30 | 6.56 b | 4.50 | 0.45 | 5.17 | 6.19 |
| Year | |||||||||||
| 2020 | 11.58 | 8.20 b | 0.82 a | 20.60 b | 4.69 b | 0.29 | 6.11 b | 4.41 b | 0.51 a | 5.41 a | 6.53 a |
| 2021 | 12.86 | 10.76 a | 0.23 b | 23.86 a | 7.85 a | 0.31 | 8.07 a | 4.56 a | 0.41 b | 5.21 b | 6.03 b |
| Treat. × Year | |||||||||||
| MIC 2020 | 12.24 | 7.80 | 1.30 a | 21.34 | 4.60 | 0.26 | 5.40 | 4.39 | 0.54 | 5.47 a–c | 6.58 |
| EKO 2020 | 12.07 | 8.24 | 0.26 c | 20.57 | 5.06 | 0.30 | 7.67 | 4.42 | 0.47 | 5.53 ab | 6.74 |
| FID 2020 | 10.61 | 7.87 | 1.03 ab | 19.50 | 4.41 | 0.30 | 5.59 | 4.38 | 0.52 | 5.60 a | 6.56 |
| CTRL 2020 | 11.39 | 8.89 | 0.71 b | 20.99 | 4.71 | 0.30 | 5.80 | 4.44 | 0.49 | 5.03 d | 6.24 |
| MIC 2021 | 13.03 | 10.33 | 0.16 c | 23.51 | 7.95 | 0.31 | 8.15 | 4.49 | 0.42 | 5.23 a–d | 5.98 |
| EKO 2021 | 12.16 | 10.80 | 0.27 c | 23.23 | 7.99 | 0.33 | 8.30 | 4.61 | 0.40 | 5.13 cd | 5.92 |
| FID 2021 | 13.50 | 11.07 | 0.24 c | 24.81 | 7.55 | 0.30 | 8.50 | 4.58 | 0.41 | 5.17 b–d | 6.07 |
| CTRL 2021 | 12.77 | 10.87 | 0.24 c | 23.88 | 7.91 | 0.30 | 7.32 | 4.56 | 0.41 | 5.30 a–d | 6.14 |
| Significance | |||||||||||
| Treatment | ns | ns | *** | ns | ns | ns | * | ns | ns | ns | ns |
| Year | ns | *** | *** | * | *** | ns | *** | ** | *** | * | *** |
| Treat. × Year | ns | ns | *** | ns | ns | ns | ns | ns | ns | * | ns |
| Mean | 12.22 | 9.48 | 0.53 | 22.23 | 6.27 | 0.30 | 7.09 | 4.48 | 0.46 | 5.31 | 6.28 |
| Prot | Ala | Asn | Asp | GABA | Gln | Glu | Gly | MEA | Orn | Pro | Ser | Tyr | TAA | EAA | BCAAs | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | (µmol g−1) | |
| Treatment | ||||||||||||||||
| MIC | 3.60 b | 1.82 b | 4.36 b | 5.08 b | 11.16 b | 14.33 b | 14.14 | 0.23 c | 0.45 b | 0.16 | 0.80 | 1.06 | 0.71 b | 58.28 b | 2.29 b | 0.94 b |
| EKO | 3.69 b | 2.52 ab | 4.97 ab | 4.92 b | 11.15 b | 14.99 b | 13.63 | 0.29 bc | 0.44 b | 0.14 | 0.66 | 1.26 | 0.72 b | 59.70 b | 2.59 b | 1.09 b |
| FID | 5.30 a | 3.06 a | 5.50 ab | 7.24 a | 18.31 a | 22.65 a | 16.18 | 0.41 a | 0.55 a | 0.15 | 0.71 | 1.47 | 1.02 a | 82.71 a | 3.42 a | 1.48 a |
| CTRL | 3.45 b | 2.63 ab | 6.20 a | 5.52 b | 10.37 b | 14.60 b | 14.78 | 0.32 b | 0.46 ab | 0.15 | 0.73 | 1.23 | 0.81 ab | 61.93 b | 2.58 b | 1.02 b |
| Year | ||||||||||||||||
| 2020 | 2.14 b | 3.15 a | 6.66 a | 4.65 b | 10.22 b | 13.71 b | 12.87 b | 0.34 a | 0.39 b | 0.18 a | 0.98 a | 1.51 a | 0.78 | 59.63 a | 2.72 | 1.12 |
| 2021 | 5.88 a | 1.86 b | 3.86 b | 6.73 a | 15.28 a | 19.57 a | 16.49 a | 0.28 b | 0.56 a | 0.12 b | 0.47 b | 1.00 b | 0.86 | 71.67 b | 2.72 | 1.14 |
| Treat. × Year | ||||||||||||||||
| MIC 2020 | 2.30 c | 2.10 b–d | 4.99 | 4.16 b | 8.90 b | 10.89 b | 11.05 | 0.22 b | 0.41 bc | 0.19 a | 1.17 a | 1.16 | 0.64 | 49.60 b | 2.12 b | 0.88 b |
| EKO 2020 | 2.30 c | 3.38 ab | 6.50 | 4.28 b | 9.66 b | 13.10 b | 10.27 | 0.34 ab | 0.34 c | 0.15 bc | 0.87 b | 1.50 | 0.75 | 54.90 b | 2.51 b | 1.07 b |
| FID 2020 | 2.13 c | 3.23 a–c | 7.14 | 5.18 b | 11.98 b | 15.73 b | 15.01 | 0.39 a | 0.39 bc | 0.18 ab | 0.90 b | 1.72 | 0.88 | 67.38 b | 3.14 ab | 1.28 ab |
| CTRL 2020 | 1.85 c | 3.92 a | 8.00 | 4.98 b | 10.33 b | 15.12 b | 15.17 | 0.42 a | 0.41 bc | 0.19 a | 0.99 ab | 1.68 | 0.83 | 66.67 b | 3.11 ab | 1.27 ab |
| MIC 2021 | 4.90 b | 1.54 d | 3.72 | 6.01 b | 13.43 b | 17.77 b | 17.24 | 0.23 b | 0.49 bc | 0.12 c | 0.43 c | 0.95 | 0.78 | 66.95 b | 2.45 b | 1.01 b |
| EKO 2021 | 5.09 b | 1.67 cd | 3.43 | 5.56 b | 12.64 b | 16.88 b | 16.98 | 0.23 b | 0.53 b | 0.13 c | 0.45 c | 1.03 | 0.70 | 64.51 b | 2.66 b | 1.11 b |
| FID 2021 | 8.47 a | 2.89 a–d | 3.86 | 9.30 a | 24.64 a | 29.56 a | 17.35 | 0.43 a | 0.71 a | 0.12 c | 0.53 c | 1.22 | 1.16 | 98.04 a | 3.69 a | 1.68 a |
| CTRL 2021 | 5.05 b | 1.34 d | 4.41 | 6.06 b | 10.41 b | 14.07 b | 14.39 | 0.22 b | 0.51 b | 0.12 c | 0.46 c | 0.79 | 0.80 | 57.19 b | 2.05 b | 0.78 b |
| Significance | ||||||||||||||||
| Treatment | *** | * | * | *** | *** | *** | ns | *** | * | ns | ns | ns | ** | ** | * | * |
| Year | *** | *** | *** | *** | *** | *** | ** | * | *** | *** | *** | *** | ns | ** | ns | ns |
| Treat. × Year | *** | * | ns | * | *** | ** | ns | ** | ** | * | * | ns | ns | ** | * | * |
| Mean | 4.01 | 2.51 | 5.26 | 5.69 | 12.75 | 16.64 | 14.68 | 0.31 | 0.47 | 0.15 | 0.72 | 1.26 | 0.82 | 65.65 | 2.72 | 1.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fusco, G.M.; Burato, A.; Pentangelo, A.; Cardarelli, M.; Nicastro, R.; Carillo, P.; Parisi, M. Can Microbial Consortium Applications Affect Yield and Quality of Conventionally Managed Processing Tomato? Plants 2023, 12, 14. https://doi.org/10.3390/plants12010014
Fusco GM, Burato A, Pentangelo A, Cardarelli M, Nicastro R, Carillo P, Parisi M. Can Microbial Consortium Applications Affect Yield and Quality of Conventionally Managed Processing Tomato? Plants. 2023; 12(1):14. https://doi.org/10.3390/plants12010014
Chicago/Turabian StyleFusco, Giovanna Marta, Andrea Burato, Alfonso Pentangelo, Mariateresa Cardarelli, Rosalinda Nicastro, Petronia Carillo, and Mario Parisi. 2023. "Can Microbial Consortium Applications Affect Yield and Quality of Conventionally Managed Processing Tomato?" Plants 12, no. 1: 14. https://doi.org/10.3390/plants12010014