
Foliar Diseases and the Associated Fungi in Rice Cultivated in Kenya

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Frequency and Agronomic Characteristics of Popular Rice Cultivars
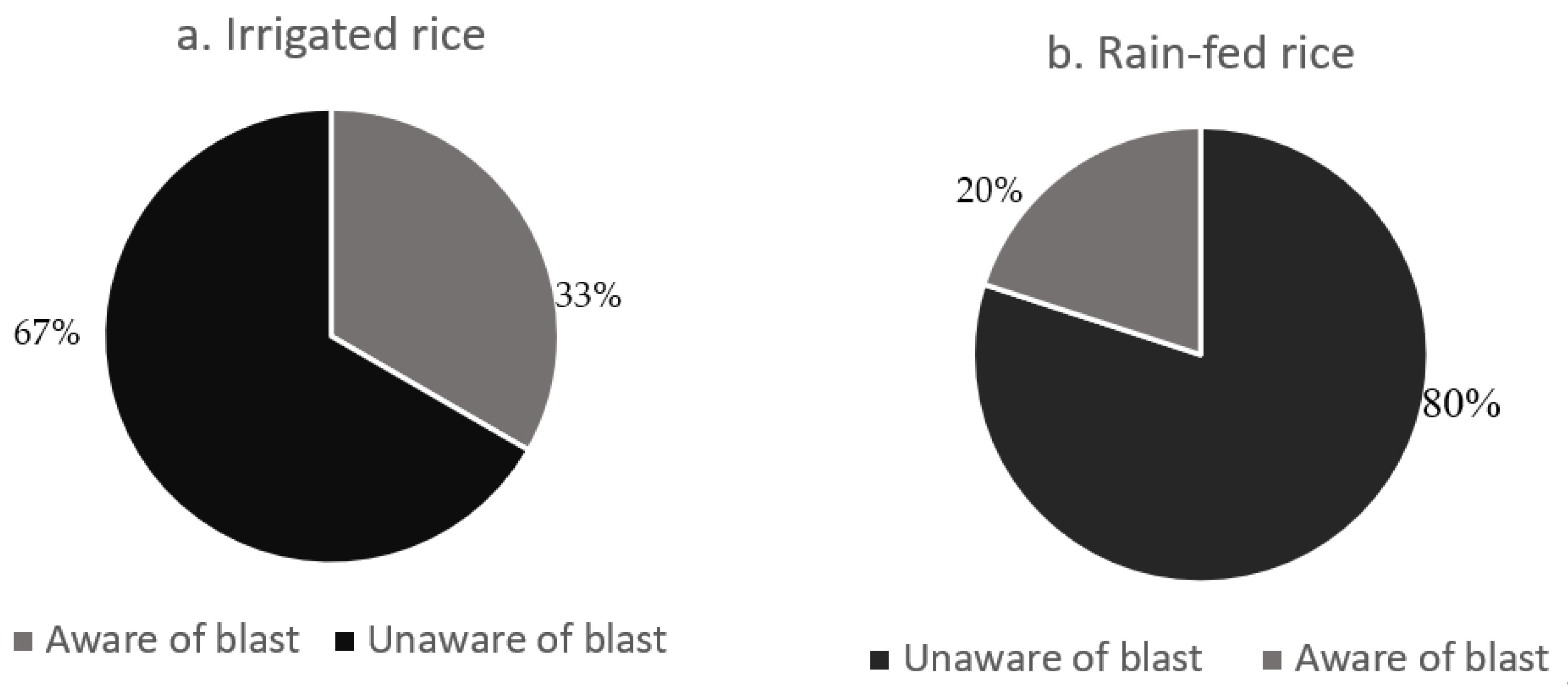
2.2. Incidence of Blast and Brown Spot Diseases and a Confirmation of Infection by the Causal Fungi
2.3. Relationship between Disease Components and Agronomic Traits of Rice
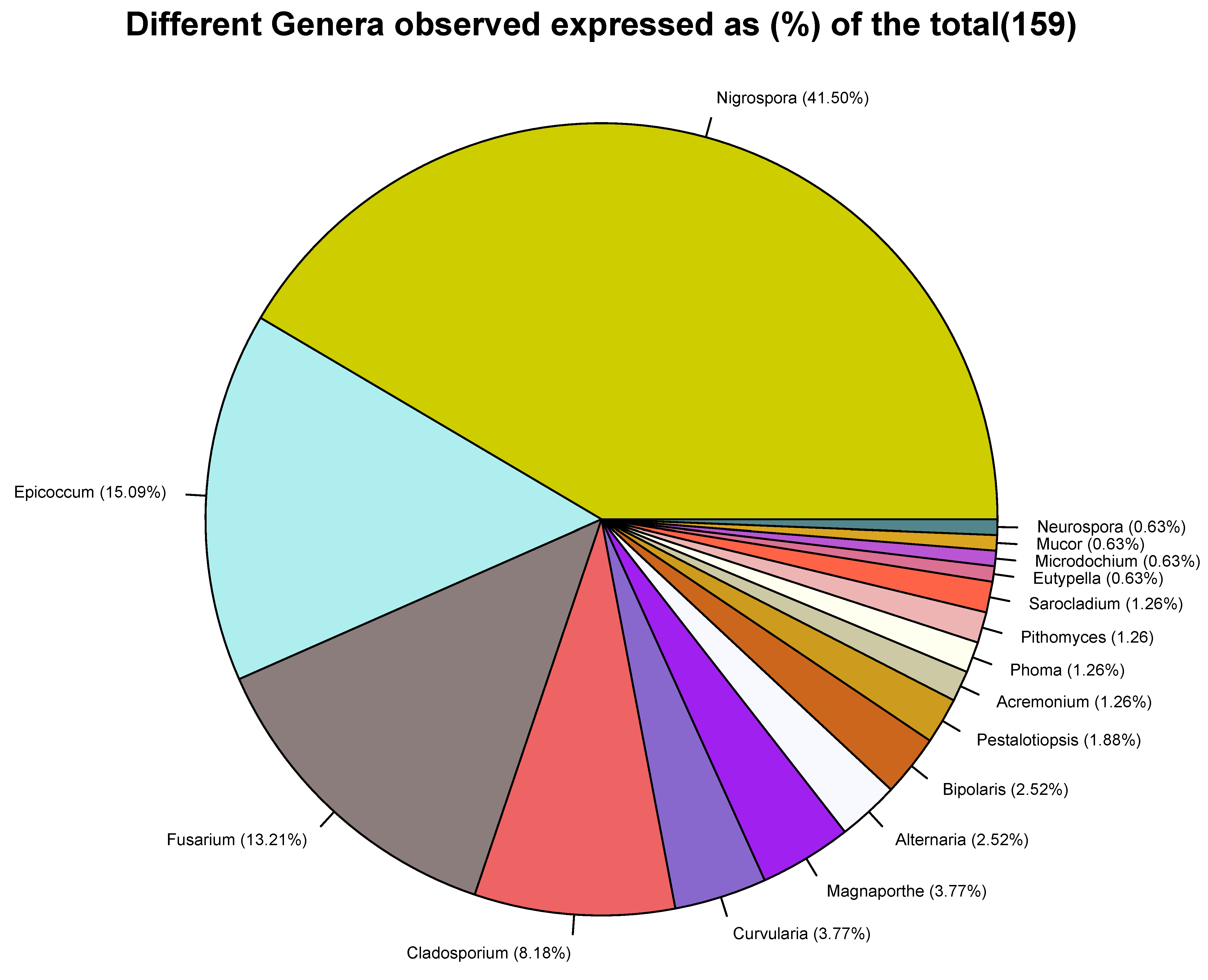
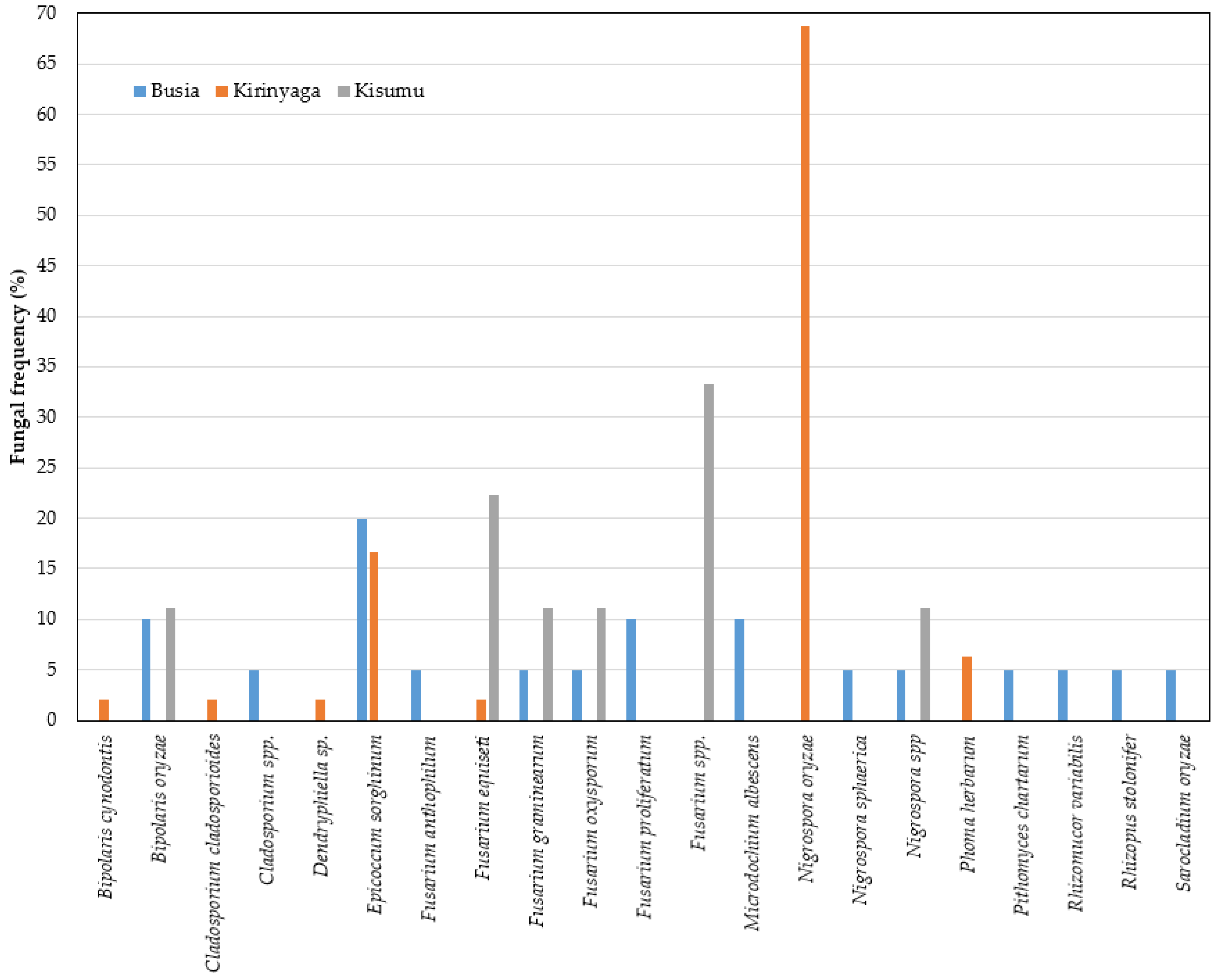
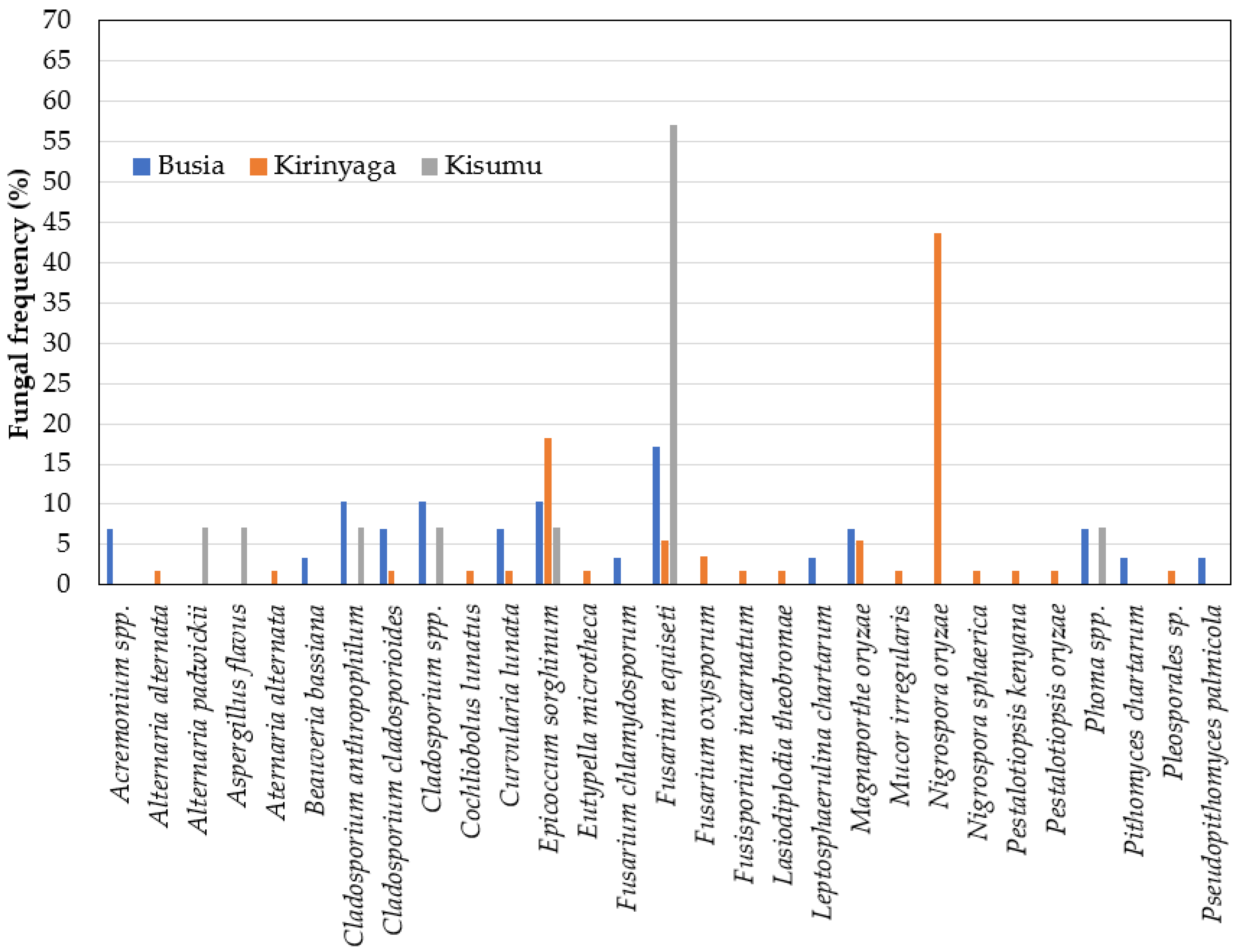
2.4. Fungal Profiles in Leaf and Neck Tissues of Rice
2.5. Relationship between Fungal Species and Rice Hosts
3. Discussion
4. Materials and Methods
4.1. Characteristics of the Study Sites
4.2. Study Design and Sampling
4.3. Sample Handling and Preparation
4.4. Molecular Detection of Magnaporthe oryzae and Cochliobolus miyabeanus in Rice
4.5. Isolation and Identification of Diverse Fungi in Rice
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mutiga, S.K.; Rotich, F.; Were, V.M.; Kimani, J.; Mwongera, D.T.; Mgonja, E.; Onaga, G.; Konaté, K.; Razanaboahirana, C.; Bigirimana, J.; et al. Integrated strategies for durable rice blast resistance in sub-Saharan Africa. Plant Dis. 2021, 105, 2749–2770. [Google Scholar] [CrossRef] [PubMed]
- MOALFC. National Rice Development Strategy-2 (2019–2030); Ministry of Agriculture, Livestock, Fisheries and Cooperatives: Nairobi, Kenya, 2020; p. 116.
- Onyango, A.O. Promotion of rice production: A likely step to making kenya food secure. An assessment of current production and potential. Dev. Ctry. Stud. 2014, 4. Available online: https://www.iiste.org/Journals/index.php/DCS/article/view/16048/16175 (accessed on 22 November 2021).
- Muema, F.M.; Home, P.G.; Raude, J.M. Application of Benchmarking and Principal Component Analysis in Measuring Performance of Public Irrigation Schemes in Kenya. Agriculture 2018, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Laha, G.S.; Singh, R.; Ladhalakshmi, D.; Sunder, S.; Prasad, M.S.; Dagar, C.S.; Babu, V.R. Importance and Management of Rice Diseases: A Global Perspective. In Rice Production Worldwide; Chauhan, B.S., Jabran, K., Mahajan, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 303–360. [Google Scholar] [CrossRef]
- Godfray, H.C.; Mason-D’Croz, D.; Robinson, S. Food system consequences of a fungal disease epidemic in a major crop. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihoro, J.; Bosco, N.J.; Murage, H.; Ateka, E.; Makihara, D. Investigating the impact of rice blast disease on the livelihood of the local farmers in greater Mwea region of Kenya. Springerplus 2013, 2, 308. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.Y.; Fang, H.; Wang, Y.Y.; Fan, J.X.; Yang, S.S.; Mew, T.W.; Mundt, C.C. Panicle blast and canopy moisture in rice cultivar mixtures. Phytopathology 2005, 95, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Séré, Y.; Fargette, D.; Abo, M.E.; Wydra, K.; Bimerew, M.; Onasanya, A.; Akator, S.K. Managing the major diseases of rice in Africa. In Realizing Africa’s Rice Promise; Wopereis, M.C.S., Johnson, D.E., Ahmadi, N., Tollens, E., Jalloh, A., Eds.; CABI: Wallingford, UK, 2013; pp. 213–228. Available online: http://www.cabi.org/bookshop/book/9781845938123 (accessed on 10 November 2021).
- Muimba-Kankolongo, A. (Ed.) Chapter 8—Cereal Production. In Food Crop Production by Smallholder Farmers in Southern Africa; Academic Press: Cambridge, MA, USA, 2018; pp. 73–121. [Google Scholar] [CrossRef]
- Savary, S.; Teng, P.S.; Willocquet, L.; Forrest, W.; Nutter, J. Quantification and Modeling of Crop Losses: A Review of Purposes. Annu. Rev. Phytopathol. 2006, 44, 89–112. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Elazegui, F.A.; Castilla, N.P.; Teng, P.S. Rice Pest Constraints in Tropical Asia: Quantification of Yield Losses Due to Rice Pests in a Range of Production Situations. Plant Dis. 2000, 84, 357–369. [Google Scholar] [CrossRef] [Green Version]
- Barnwal, M.K.; Kotasthane, A.; Magculia, N.; Mukherjee, P.K.; Savary, S.; Sharma, A.K.; Singh, H.B.; Singh, U.S.; Sparks, A.H.; Variar, M.; et al. A review on crop losses, epidemiology and disease management of rice brown spot to identify research priorities and knowledge gaps. Eur. J. Plant Pathol. 2013, 136, 443–457. [Google Scholar] [CrossRef] [Green Version]
- Constantine, K.L.; Kansiime, M.K.; Mugambi, I.; Nunda, W.; Chacha, D.; Rware, H.; Makale, F.; Mulema, J.; Lamontagne-Godwin, J.; Williams, F.; et al. Why don’t smallholder farmers in Kenya use more biopesticides? Pest Manag. Sci. 2020, 76, 3615–3625. [Google Scholar] [CrossRef]
- Naik, B.S.; Shashikala, J.; Krishnamurthy, Y.L. Study on the diversity of endophytic communities from rice (Oryza sativa L.) and their antagonistic activities in vitro. Microbiol. Res. 2009, 164, 290–296. [Google Scholar] [CrossRef]
- Lapmak, K.; Lumyong, S.; Wangspa, R.; Sardsud, U. Diversity of filamentous fungi on brown rice from Pattalung Province, Thailand. J. Agric. Technol. 2009, 5, 129–142. [Google Scholar]
- Yuan, Z.L.; Zhang, C.L.; Lin, F.C.; Kubicek, C.P. Identity, diversity, and molecular phylogeny of the endophytic mycobiota in the roots of rare wild rice (Oryza granulate) from a nature reserve in Yunnan, China. Appl. Environ. Microbiol. 2010, 76, 1642–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leewijit, T.; Pongnak, W.; Soytong, K.; Poeaim, S. Isolation of Soil and Endophytic Fungi from Rice (Oryza sativa L.). Int. J. Agric. Technol. 2016, 12, 2191–2202. [Google Scholar]
- Gaddeyya, G.; Niharika, P.S.; Bharathi, P.; Kumar, P.K.R. Isolation and identification of soil mycoflora in different crop fieldsat Salur Mandal. Adv. Appl. Sci. Res. 2012, 3, 2020–2026. [Google Scholar]
- Seephueak, P.; Preecha, C.; Seephueak, W. The diversity of fungi associated with rice (Oryza sativa L.) from Nakhon Si Thammarat, Thailand. Int. J. Agric. Technol. 2019, 15, 485–500. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Badotti, F.; de Oliveira, F.S.; Garcia, C.F.; Vaz, A.B.M.; Fonseca, P.L.C.; Nahum, L.A.; Oliveira, G.; Góes-Neto, A. Effectiveness of ITS and sub-regions as DNA barcode markers for the identification of Basidiomycota (Fungi). BMC Microbiol. 2017, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Khan, M.S.; Gao, J.; Munir, I.; Zhang, M.; Liu, Y.; Moe, T.S.; Xue, J.; Zhang, X. Characterization of Endophytic Fungi, Acremonium sp., from Lilium davidii and Analysis of Its Antifungal and Plant Growth-Promoting Effects. BioMed Res. Int. 2021, 2021, 9930210. [Google Scholar] [CrossRef]
- Gutiérrez, S.A.; Carmona, M.A.; Reis, E.M. Methods for Detection of Alternaria padwickii in Rice Seeds. J. Phytopathol. 2010, 158, 523–526. [Google Scholar] [CrossRef]
- Gonçalves, A.; Gkrillas, A.; Dorne, J.L.; Dall’Asta, C.; Palumbo, R.; Lima, N.; Battilani, P.; Venâncio, A.; Giorni, P. Pre- and Postharvest Strategies to Minimize Mycotoxin Contamination in the Rice Food Chain. Compr. Rev. Food Sci. Food Saf. 2019, 18, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Khashaba, E.H.K. Inoculation and colonization of isolated entomopathogenic fungi Beauveria bassiana in rice plants, Oryza sativa L. through seed immersion method. Egypt. J. Biol. Pest Control 2021, 31, 92. [Google Scholar] [CrossRef]
- Ahn, I.P.; Kim, S.; Kang, S.; Suh, S.C.; Lee, Y.H. Rice Defense Mechanisms Against Cochliobolus miyabeanus and Magnaporthe grisea Are Distinct. Phytopathology 2005, 95, 1248–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.M.; Huang, S.W.; Wang, L.; Hou, E.Q.; Xiao, D.F. First Report of Leaf Blight of Rice Caused by Cochliobolus lunatus in China. Plant Dis. 2014, 98, 686. [Google Scholar] [CrossRef] [PubMed]
- Louis, B.; Waikhom, S.D.; Roy, P.; Bhardwaj, P.K.; Sharma, C.K.; Singh, M.W.; Talukdar, N.C. Host-Range Dynamics of Cochliobolus lunatus: From a Biocontrol Agent to a Severe Environmental Threat. BioMed Res. Int. 2014, 2014, 378372. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.M.; Zhao, Y.; Zhang, Y.L.; Wang, L.; Hou, Y.X.; Huang, S.W. First Report of Leaf Spot Disease on Rice Caused by Epicoccum sorghinum in China. Plant Dis. 2020, 104, 2735. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, X.; Zhang, R.; Liu, J.; Ali, E.; Ji, P.; Pan, H.; Zhang, K. Occurrence of Seedling Blight Caused by Fusarium tricinctum on Rice in China. Plant Dis. 2019, 103, 1789. [Google Scholar] [CrossRef]
- Eğerci, Y.; Kınay-Teksür, P.; Uysal-Morca, A. First report of Bakanae disease caused by Fusarium proliferatum on rice in Turkey. J. Plant Dis. Prot. 2021, 128, 577–582. [Google Scholar] [CrossRef]
- Wang, L.; Ge, S.L.; Zhao, K.H.; Huang, S.W. First Report of Fusarium incarnatum Causing Spikelet Rot on Rice in China. Plant Dis. 2021, 105, 3306. [Google Scholar] [CrossRef]
- Claudius-Cole, A. Lasiodiplodia theobromae in the Root Rot Disease Complex of Rice. Rice Res. Open Access 2018, 6, 197. [Google Scholar] [CrossRef]
- Couch, B.C.; Fudal, I.; Lebrun, M.H.; Tharreau, D.; Valent, B.; van Kim, P.; Nottéghem, J.L.; Kohn, L.M. Origins of host-specific populations of the blast pathogen Magnaporthe oryzae in crop domestication with subsequent expansion of pandemic clones on rice and weeds of rice. Genetics 2005, 170, 613–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, H.; Black, R. Rice Leaf Scald: Pathogen Biology and Diversity. In Major Fungal Diseases of Rice; Springer: New York, NY, USA, 2001; pp. 307–319. [Google Scholar]
- Liu, Y.L.; Tang, J.R.; Li, Y.; Zhou, H.K. First Report of Leaf Spots caused by Nigrospora oryzae on Wild Rice in China. Plant Dis. 2021, 105, 3293. [Google Scholar] [CrossRef] [PubMed]
- Lau, K.H.; Sheridan, J.E. Mycoflora of rice (Oryza sativa L.) seed in New Zealand. N. Z. J. Agric. Res. 1975, 18, 237–250. [Google Scholar] [CrossRef]
- Bigirimana, V.d.P.; Hua, G.K.H.; Nyamangyoku, O.I.; Höfte, M. Rice Sheath Rot: An Emerging Ubiquitous Destructive Disease Complex. Front. Plant Sci. 2015, 6, 1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalley, L.; Tsiboe, F.; Durand-Morat, A.; Shew, A.; Thoma, G. Economic and Environmental Impact of Rice Blast Pathogen (Magnaporthe oryzae) Alleviation in the United States. PLoS ONE 2016, 11, e0167295. [Google Scholar] [CrossRef] [PubMed]
- Yamane, Y. Rice Distribution in Kenya: The Conditions of Rice Traders in the Rice Cultivation Area around Lake Victoria. Agric. Res. Technol. Open Access J. 2019, 19, 556099. [Google Scholar] [CrossRef]
- Watanabe, M.; Sumita, Y.; Azechi, I.; Ito, K.; Noda, K. Production Costs and Benefits of Japonica Rice in Mwea, Kenya. Agriculture 2021, 11, 629. [Google Scholar] [CrossRef]
- Mwongera, D. Determining the Population Structure and Avirulence Gene Repertoire of the Rice Blast Fungus Magnaporthe oryzae in Kenya by Comparative Genome Analysis; University of Exeter: Exeter, UK, 2018; Available online: https://ore.exeter.ac.uk/repository/handle/10871/33087 (accessed on 12 February 2022).
- KEPHIS. National Crop Variety List—Kenya; Kenya Plant Health Inspectorate Service: Nairobi, Kenya, 2020; p. 169. Available online: https://kephis.org/images/pdf-files (accessed on 27 August 2021).
- Seidel, D.; Herrera, L. Survival of Cochliobolus miyabeanus in the soil (author’s transl). Zent. Bakteriol Nat. 1979, 134, 40–42. [Google Scholar]
- Mwendo, M.M.; Ochwo-Ssemakula, M.; Mwale, S.E.; Lamo, J.; Gibson, P.; Edema, R. Inheritance of resistance to brown spot disease in upland rice in Uganda. J. Plant Breed. Crop Sci. 2017, 9, 37–44. [Google Scholar]
- Sapre, S.; Gontia-Mishra, I.; Thakur, V.V.; Sikdar, S.; Tiwari, S. Chapter 20—Molecular techniques used in plant disease diagnosis. In Food Security and Plant Disease Management; Kumar, A., Droby, S., Eds.; Woodhead Publishing: Sawston, UK, 2021; pp. 405–421. [Google Scholar] [CrossRef]
- Khemmuk, W.; Shivas, R.G.; Henry, R.J.; Geering, A.D.W. Fungi associated with foliar diseases of wild and cultivated rice (Oryza spp.) in northern Queensland. Australas. Plant Pathol. 2016, 45, 297–308. [Google Scholar] [CrossRef]
- Baldi, P.; La Porta, N. Molecular Approaches for Low-Cost Point-of-Care Pathogen Detection in Agriculture and Forestry. Front. Plant Sci. 2020, 11, 570862. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, K.; Zhao, Y.; Zhang, Y.; Fu, Q.; Shiwen, H. Nigrospora oryzae Causing Panicle Branch Rot Disease on Oryza sativa (rice). Plant Dis. 2021, 105, 2724. [Google Scholar] [CrossRef] [PubMed]
- Bhunjun, C.S.; Phillips, A.J.L.; Jayawardena, R.S.; Promputtha, I.; Hyde, K.D. Importance of Molecular Data to Identify Fungal Plant Pathogens and Guidelines for Pathogenicity Testing Based on Koch’s Postulates. Pathogens 2021, 10, 1096. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; Gordon, T.R. Cryptic fungal infections: The hidden agenda of plant pathogens. Front. Plant Sci. 2014, 5, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef] [Green Version]
- Tamošiūnė, I.; Stanienė, G.; Haimi, P.; Stanys, V.; Rugienius, R.; Baniulis, D. Endophytic Bacillus and Pseudomonas spp. Modulate Apple Shoot Growth, Cellular Redox Balance, and Protein Expression under in Vitro Conditions. Front. Plant Sci. 2018, 9, 889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atugala, D.; Deshappriya, N. Effect of endophytic fungi on plant growth and blast disease incidence of two traditional rice varieties. J. Natl. Sci. Found. Sri Lanka 2015, 43, 173. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted Interactions between Endophytes and Plant: Developments and Prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Martinelli, F.; Scalenghe, R.; Davino, S.; Panno, S.; Scuderi, G.; Ruisi, P.; Villa, P.; Stroppiana, D.; Boschetti, M.; Goulart, L.R.; et al. Advanced methods of plant disease detection. A review. Agron. Sustain. Dev. 2015, 35, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Hibbett, D.S.; Ohman, A.; Glotzer, D.; Nuhn, M.; Kirk, P.; Nilsson, R.H. Progress in molecular and morphological taxon discovery in Fungi and options for formal classification of environmental sequences. Fungal Biol. Rev. 2011, 25, 38–47. [Google Scholar] [CrossRef]
- Silvestri, S.; Macharia, M.; Uzayisenga, B. Analysing the potential of plant clinics to boost crop protection in Rwanda through adoption of IPM: The case of maize and maize stem borers. Food Secur. 2019, 11, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Otieno, W.; Ochilo, W.; Migiro, L.; Jenner, W.; Kuhlmann, U. Tools for pest and disease management by stakeholders: A case study on Plantwise. In The Sustainable Intensification of Smallholder Farming Systems; Klauser, D., Robinson, M., Eds.; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2020. [Google Scholar] [CrossRef]
- Sorre, A.M. Effects of climate change on rural livelihoods in Busia county, Kenya. Int. J. Agric. Earth Sci. 2017, 3. Available online: https://www.iiardjournals.org/get/IJAES/VOL.%203%20NO.%208%202017 (accessed on 15 February 2022).
- MoALF. Climate Risk Profile for Kisumu County; Kenya County Climate Risk Profile Series; The Ministry of Agriculture: Nairobi, Kenya, 2017.
- Mulinge, W.M.; Mwaniki, A.; Murithi, F.M.; Croce, S. Analysis of Price Incentives and Disincentives for Rice in Kenya over the Period 2005–2013; FAO: Rome, Italy, 2019. [Google Scholar]
- IRRI. Standard Evaluation System (SES) for Rice, 5th ed.; International Rice Research Institute: Los Banos, Philippines, 2013; p. 57. [Google Scholar]
- Abebrese, S.O.; Yeboah, A.; Dogbe, W.; Dartey, P.K.A.; Akromah, R.; Gracen, V.E.; Offei, S.K.; Danquah, E.Y. Evaluation of Yield, Reaction to Diseases, and Grain Physical Attributes of Some Introduced Rice Hybrids in Ghana. Int. J. Agron. 2019, 2019, 3926765. [Google Scholar] [CrossRef] [Green Version]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmon, P.F.; Dunkle, L.D.; Latin, R. A Rapid PCR-Based Method for the Detection of Magnaporthe oryzae from Infected Perennial Ryegrass. Plant Dis. 2003, 87, 1072–1076. [Google Scholar] [CrossRef] [Green Version]
- Su’udi, M.; Park, J.-M.; Kang, W.-R.; Park, S.-R.; Hwang, D.-J.; Ahn, I.-P. Quantification of rice brown leaf spot through Taqman real-time PCR specific to the unigene encoding Cochliobolus miyabeanus SCYTALONE DEHYDRATASE1 involved in fungal melanin biosynthesis. J. Microbiol. 2012, 50, 947–954. [Google Scholar] [CrossRef]
- Namulawa, V.T.; Mutiga, S.; Musimbi, F.; Akello, S.; Ngángá, F.; Kago, L.; Kyallo, M.; Harvey, J.; Ghimire, S. Assessment of Fungal Contamination in Fish Feed from the Lake Victoria Basin, Uganda. Toxins 2020, 12, 233. [Google Scholar] [CrossRef] [Green Version]
- Pitt, J.I.; Hocking, A.D. Methods for isolation, enumeration and identification. In Fungi and Food Spoilage; Springer: Boston, MA, USA, 1997; pp. 21–57. [Google Scholar] [CrossRef]
- Dooso Oloo, R.; Okoth, S.; Wachira, P.; Mutiga, S.; Ochieng, P.; Kago, L.; Nganga, F.; Domelevo Entfellner, J.-B.; Ghimire, S. Genetic Profiling of Aspergillus Isolates with Varying Aflatoxin Production Potential from Different Maize-Growing Regions of Kenya. Toxins 2019, 11, 467. [Google Scholar] [CrossRef] [Green Version]
- Manter, D.K.; Vivanco, J.M. Use of the ITS primers, ITS1F and ITS4, to characterize fungal abundance and diversity in mixed-template samples by qPCR and length heterogeneity analysis. J. Microbiol. Methods 2007, 71, 7–14. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bozdogan, H. Model selection and Akaike’s Information Criterion (AIC): The general theory and its analytical extensions. Psychometrika 1987, 52, 345–370. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
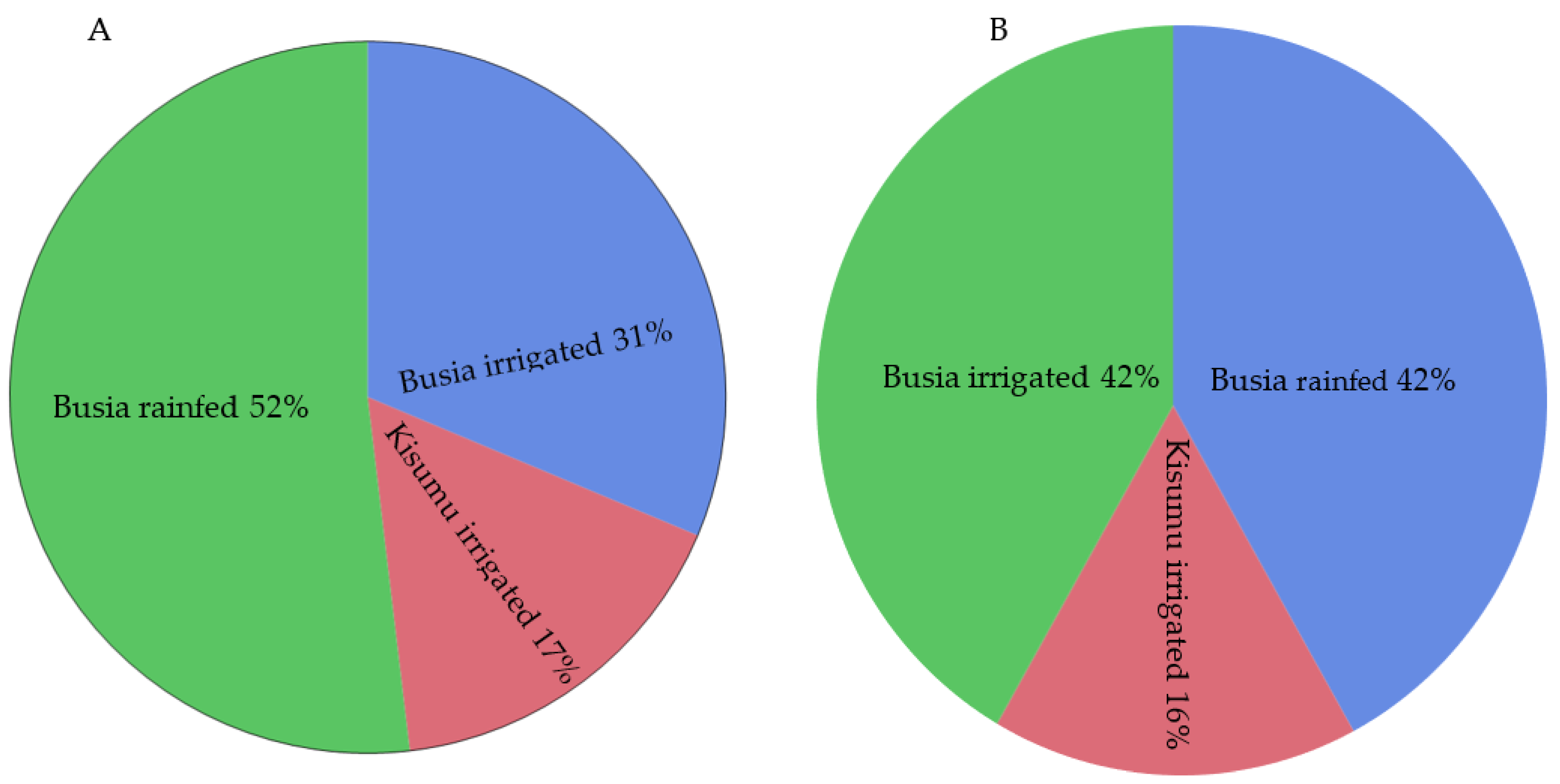
| Size of Farm (ha) | Farmers (n) | Rice Tissue Samples (n) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Leaves | Neck | ||||||||
| County | Site | Irrigation Type | Minimum | Mean | Maximum | Asymptomatic | ¥ Symptomatic | Symptomatic | |
| Busia | Bunyala Central, Busia | Canal | 0.04 | 0.34 | 0.65 | 20 | 20 | 32 | 7 |
| Ng’elechom, Busia | Canal | 0.40 | 0.607 | 0.81 | 3 | 3 | 6 | 3 | |
| Rain-fed | 0.10 | 1.127 | 4.05 | 12 | 12 | 24 | 8 | ||
| Kisumu | Ahero, Kisumu | Canal | 0.40 | 0.903 | 3.24 | 15 | 11 | 23 | 6 |
| West Kano, Kisumu | Canal | 0.40 | 0.549 | 0.81 | 13 | 10 | 18 | 2 | |
| Kirinyaga | Mwea East, Kirinyaga | Canal | 0.10 | 0.751 | 4.05 | 19 | 8 | 19 | 19 |
| Mwea West, Kirinyaga | Canal | 0.20 | 0.549 | 1.21 | 7 | 4 | 7 | 7 | |
| County | Rice cultivar | Favorable Trait | Grower (n) | Growers (%) |
|---|---|---|---|---|
| Busia | Abednego from Uganda | Long grain and high yield | 2 | 6 |
| KEH10004 (Arize 6444) | High tillering | 1 | 3 | |
| KEH10005 (Arize Tej Gold) | Less damage by pests and high grain yield | 3 | 7 | |
| NIBAM11 (Pishori) | Aroma and ready market | 3 | 12 | |
| NIBAM109 (BW196) | Tolerant to multiple stresses | 3 | 9 | |
| NIBAM108 (IR2793-80-1) | Resistant to multiple diseases | 5 | 14 | |
| NIBAM108 mixed with NIBAM109 | These were the only cultivars whose seed was available for them. | 1 | 3 | |
| NIBAM110 (ITA310) | High grain yield | 4 | 11 | |
| IR05N221 (Komboka) | Aroma and high grain yield | 3 | 9 | |
| Pakistan K23 from Uganda | Drought tolerant and high grain yield | 2 | 6 | |
| Palata from Uganda | Aroma and high grain yield | 1 | 3 | |
| Vietnam from Uganda | High grain yield | 6 | 17 | |
| subtotal | 35 | |||
| Kisumu | KEH10005 | - | 2 | 7 |
| NIBAM11 | - | 2 | 7 | |
| NIBAM108 | - | 9 | 32 | |
| NIBAM110 | - | 7 | 25 | |
| IR05N221 | - | 5 | 18 | |
| Nyaboda from Uganda | High grain yield | 2 | 7 | |
| SUPA 1 from Uganda | High grain yield | 1 | 4 | |
| subtotal | 28 | |||
| Kirinyaga | NIBAM11 | - | 24 | 92 |
| IR05N221 | - | 1 | 4 | |
| NIBAM109 | - | 1 | 4 | |
| subtotal | 26 | |||
| Total | 89 |
| Source | DF | Sum of Squares | Mean Square | F Ratio | Prob > F |
|---|---|---|---|---|---|
| Blast Incidence | |||||
| Cultivar | 9 | 7781.5505 | 864.617 | 2.2543 | 0.0525 |
| Production system ∞ | 1 | 343.6142 | 343.614 | 0.8959 | 0.3529 |
| Knowledge of blast | 1 | 6553.9186 | 6553.919 | 17.0883 | 0.0004 |
| County | 1 | 431.4867 | 431.487 | 1.125 | 0.299 |
| Model | 12 | 20095.878 | 1674.66 | 4.3664 | 0.0009 |
| Error | 25 | 9588.333 | 383.53 | ||
| C. Total | 37 | 29,684.211 | |||
| Blast Severity | |||||
| Cultivar | 9 | 68.864343 | 7.65159 | 2.81 | 0.022 |
| Production system | 1 | 2.669446 | 2.66945 | 0.9803 | 0.3324 |
| Knowledge of blast | 1 | 42.862593 | 42.86259 | 15.7408 | 0.0006 |
| County | 1 | 14.643451 | 14.64345 | 5.3776 | 0.0296 |
| Model | 12 | 212.12039 | 17.6767 | 6.4916 | <0.0001 |
| Error | 23 | 62.62961 | 2.723 | ||
| C. Total | 35 | 274.75 | |||
| Brown Spot Incidence | |||||
| Cultivar | 9 | 14,758.087 | 1639.787 | 3.1472 | 0.0106 |
| Production system | 1 | 1377.274 | 1377.274 | 2.6434 | 0.116 |
| Knowledge of blast | 1 | 5511.005 | 5511.005 | 10.5771 | 0.0032 |
| County | 1 | 6527.422 | 6527.422 | 12.5279 | 0.0015 |
| Model | 12 | 40,119.827 | 3343.32 | 6.4167 | <0.0001 |
| Error | 26 | 13,546.839 | 521.03 | ||
| C. Total | 38 | 53,666.667 | |||
| Cultivar | Growers (n) | Incidence (% ± SE) | Growers (n) | Severity (0–9) ± SE | § Infection Rate (%) |
|---|---|---|---|---|---|
| Blast | |||||
| NIBAM11 (Pishori) | 10 | 0 ± 9.4 C | 10 | 0 B | 50 |
| Pakistan K23 | 3 | 0.4 ± 13.2 BC | 3 | 1.5 ± 1.1 AB | 33 |
| Palata from Uganda | 1 | 0.4 ± 13.2 ABC | 1 | 1.5 ± 1.8 AB | 0 |
| Abednego from Uganda | 2 | 5.8 ± 23.4 ABC | 2 | 2.2 ± 2 AB | 50 |
| NIBAM109 (BW196) | 5 | 8 ± 8 ABC | 5 | 0.8 ± 0.8 B | 80 |
| IR05N221 (Komboka) | 10 | 16.9 ± 7.7 ABC | 10 | 2.8 ± 0.8 AB | 40 |
| Nyaboda from Uganda | 2 | 19.6 ± 15.5 ABC | 2 | 2.5 ± 1.3 AB | 50 |
| KEH10005 (Arize Tej Gold) | 6 | 19.6 ± 15.5 ABC | 6 | 2.5 ± 1.3 AB | 67 |
| NIBAM108 (IR2793-80-1) | 14 | 21.9 ± 8.9 AB | 14 | 3.2 ± 0.8 A | 36 |
| NIBAM110 (ITA310) | 10 | 33.4 ± 10.1 AB | 10 | 3.9 ± 0.9 A | 80 |
| Vietnam | 7 | 35.4 ± 12 A | 7 | 2.5 ± 1.0 AB | 71 |
| Brown Spot | |||||
| NIBAM11 | 10 | 16.7 ± 10.8 AB | 70 | ||
| Pakistan K23 | 3 | 21.4 ± 15.3 AB | 67 | ||
| Palata | 1 | 61.4 ± 24.1 AB | 100 | ||
| Abednego | 2 | 32.5 ± 27.3 AB | 100 | ||
| NIBAM109 (BW196) | 5 | 34 ± 14 AB | 60 | ||
| IR05N221 (Komboka) | 10 | 60.3 ± 9 A | 70 | ||
| Nyaboda | 2 | 28.6 ± 17.9 AB | 100 | ||
| KEH10005 (Arize Tej Gold) | 6 | 28.6 ± 17.9 AB | 33 | ||
| NIBAM108 (IR2793-80-1) | 14 | 61.0 ± 10.3 A | 64 | ||
| NIBAM110 (ITA310) | 10 | 7.8 ± 11.8 B | 80 | ||
| Vietnam | 7 | 58.2 ± 12.1 AB | 57 | ||
| Fungal Species | Where Isolated in Rice | Effect on Rice | Reference |
|---|---|---|---|
| Acremonium spp. | neck | Potentially protective | [24] |
| Alternaria padwickii | neck | Seed rot | [25] |
| Aspergillus flavus | neck | Seed rot | [26] |
| Beauveria bassiana | neck | Potentially protective | [27] |
| Cochliobolus miyabeanus | leaf | Brown spot | [28] |
| Cladosporium anthropophilum | neck | No reported disease (NRD) | |
| Cladosporium cladosporioides | neck | NRD | |
| Cladosporium spp. | leaf and neck | NRD | |
| Cochliobolus lunatus | neck | Leaf spot, seedling blight and sheath rot | [29,30] |
| Epicoccum sorghinum | leaf and neck | Leaf spot | [31] |
| Eutypella microtheca | neck | NRD | |
| Fusarium anthophilum | leaf | NRD | |
| Fusarium chlamydosporum | neck | NRD | |
| Fusarium equiseti | leaf and neck | NRD | |
| Fusarium graminearum | leaf | Head blight | |
| Fusarium oxysporum | leaf | Seedling blight | [32] |
| Fusarium proliferatum | leaf | Bakanae disease | [33] |
| Fusarium incarnatum | neck | Spikelet rot | [34] |
| Lasiodiplodia theobromae | neck | Root rot complex disease | [35] |
| Magnaporthe oryzae | neck | Blast | [36] |
| Microdochium albescens | leaf | Leaf scald | [37] |
| Mucor irregularis | neck | NRD | |
| Nigrospora sphaerica | leaf | NRD | |
| Nigrospora oryzae | leaf | Leaf spot | [38] |
| Pestalotiopsis kenyana | neck | NRD | |
| Pestalotiopsis oryzae | neck | NRD | |
| Phoma herbarum | neck | Endophyte | |
| Leptosphaerulina chartarum | leaf and neck | Glume blotch | [39] |
| Pseudopithomyces palmicola | neck | NRD | |
| Rhizopus stolonifer | leaf | NRD | |
| Sarocladium oryzae | leaf | Sheath rot | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nganga, E.M.; Kyallo, M.; Orwa, P.; Rotich, F.; Gichuhi, E.; Kimani, J.M.; Mwongera, D.; Waweru, B.; Sikuku, P.; Musyimi, D.M.; et al. Foliar Diseases and the Associated Fungi in Rice Cultivated in Kenya. Plants 2022, 11, 1264. https://doi.org/10.3390/plants11091264
Nganga EM, Kyallo M, Orwa P, Rotich F, Gichuhi E, Kimani JM, Mwongera D, Waweru B, Sikuku P, Musyimi DM, et al. Foliar Diseases and the Associated Fungi in Rice Cultivated in Kenya. Plants. 2022; 11(9):1264. https://doi.org/10.3390/plants11091264
Chicago/Turabian StyleNganga, Everlyne M., Martina Kyallo, Philemon Orwa, Felix Rotich, Emily Gichuhi, John M. Kimani, David Mwongera, Bernice Waweru, Phoebe Sikuku, David M. Musyimi, and et al. 2022. "Foliar Diseases and the Associated Fungi in Rice Cultivated in Kenya" Plants 11, no. 9: 1264. https://doi.org/10.3390/plants11091264