
Water Deficit Improves Reproductive Fitness in Nicotiana benthamiana Plants Infected by Cucumber mosaic virus

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
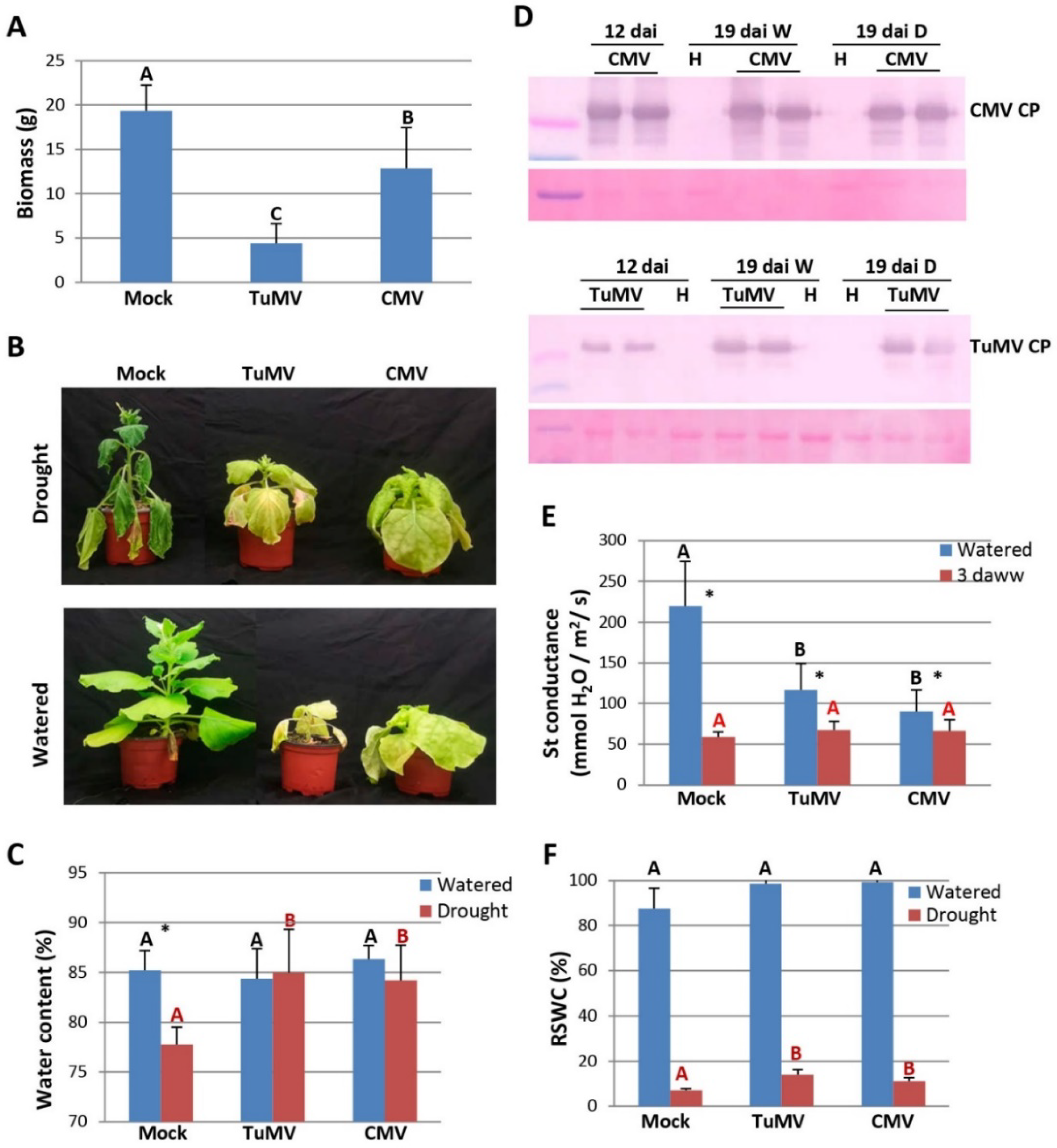
2.1. Tolerance to Drought in Virus-Infected N. Benthamiana Plants
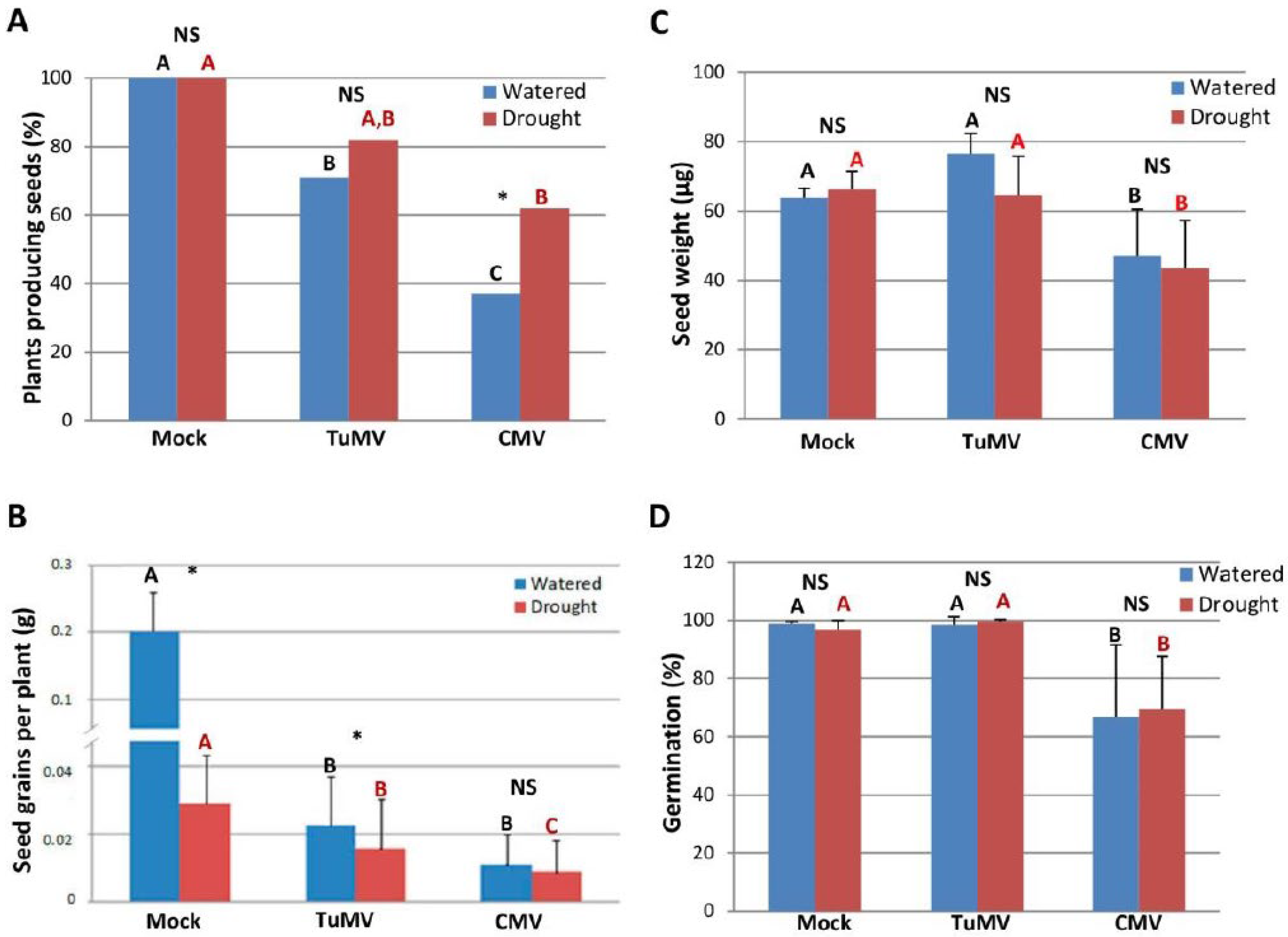
2.2. CMV-Infected N. Benthamiana Plants Improved Their Reproductive Fitness When Exposed to Drought
2.3. Tolerance to Drought in Virus-Infected Arabidopsis Plants
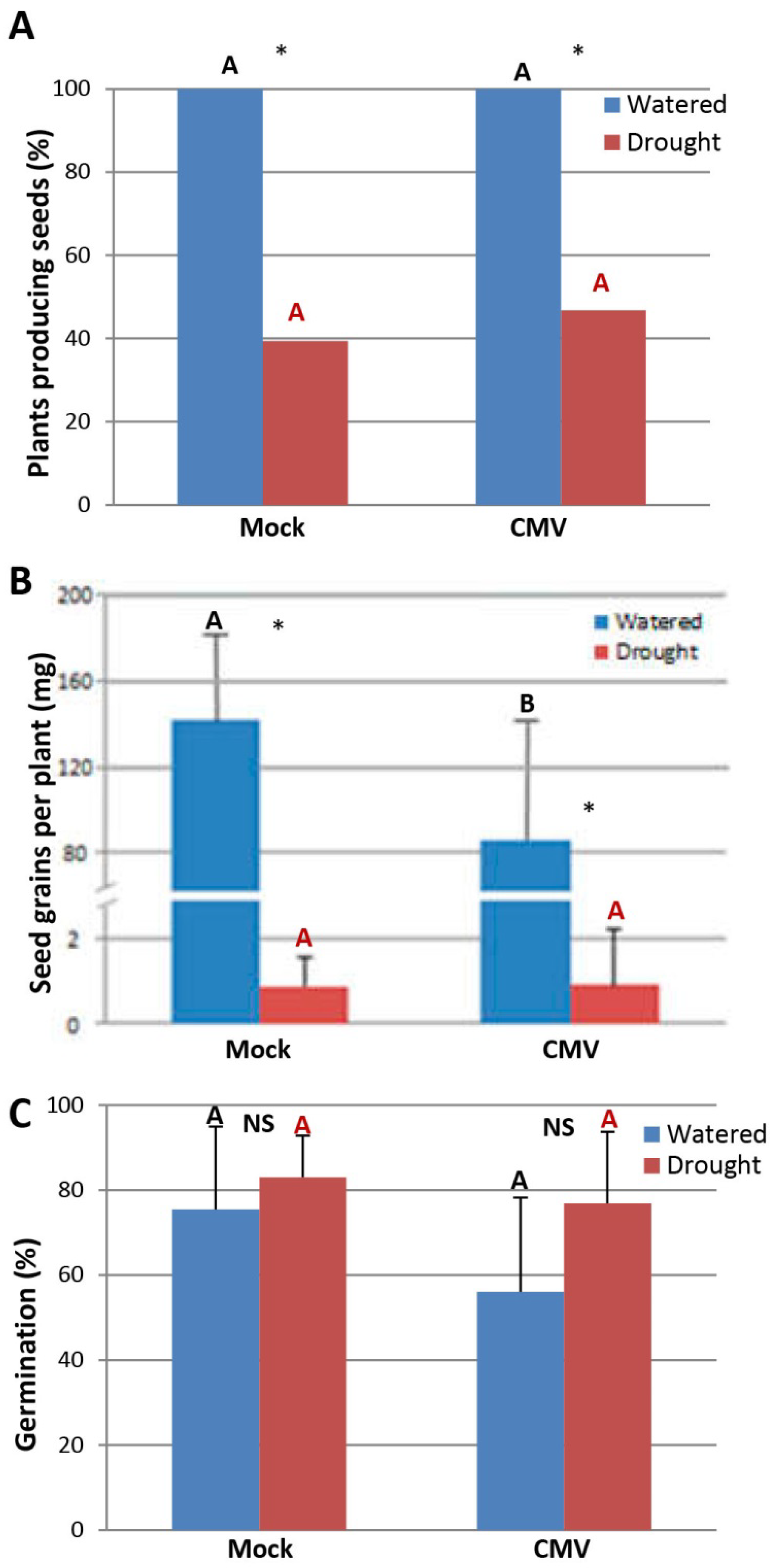
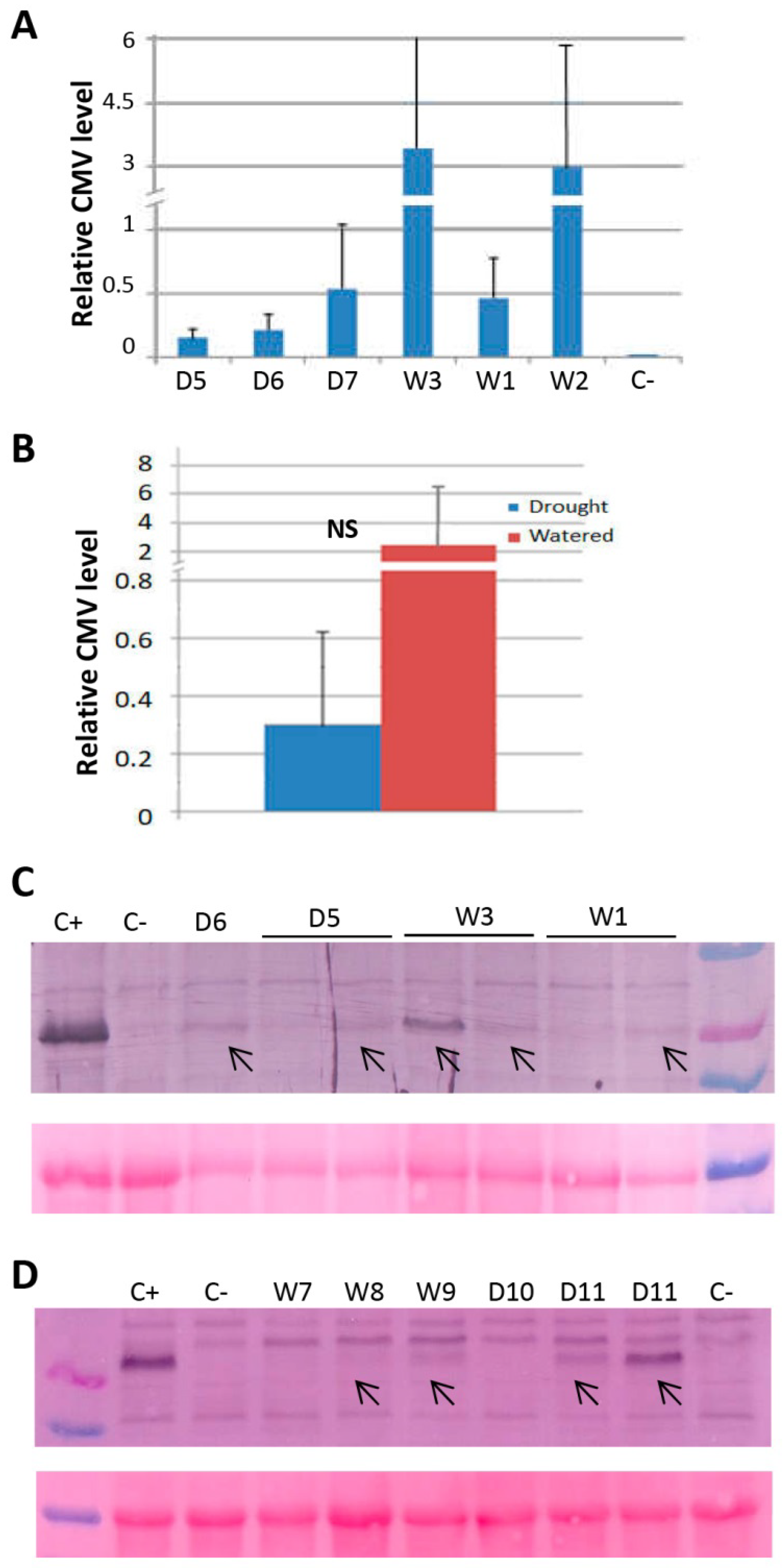
2.4. The Effect of Drought on the Transmission of CMV through N. Benthamiana and Arabidopsis Seeds
3. Discussion
4. Materials and Methods
4.1. Viruses
4.2. Plant Materials
4.3. Drought, Stomatal Conductance and Water Content Measurements
4.4. Reproductive Fitness Assays
4.5. Protein Gel Blot Analysis
4.6. Quantitative RT-PCR (qRT-PCR)
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prasch, C.M.; Sonnewald, U. Signaling events in plants: Stress factors in combination change the picture. Environ. Exp. Bot. 2015, 114, 4–14. [Google Scholar] [CrossRef]
- Ramegowda, V.; Senthil-Kumar, M. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant Physiol. 2015, 176, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rejeb, I.B.; Pastor, V.; Mauch-Mani, B. Plant Responses to Simultaneous Biotic and Abiotic Stress: Molecular Mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Márquez, L.M.; Redman, R.S.; Rodriguez, R.J.; Roossinck, M.J. A virus in a fungus in a plant—Three way symbiosis required for thermal tolerance. Science 2007, 315, 513–515. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Ahmad, M.S.A.; Ozturk, M.; Aksoy, A. Crop improvement through different means: Challenges and prospects. In Crop Production for Agricultural Improvement; Springer: Dordrecht, The Netherlands, 2012; pp. 1–15. [Google Scholar]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. (Eds.) Climate Change 2021. The Physical Science Basis. Summary for Policymakers; IPCC: Geneva, Switzerland, 2021. [Google Scholar]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef]
- Zhang, H.; Sonnewald, U. Differences and commonalities of plant responses to single and combined stresses. Plant J. 2017, 90, 839–855. [Google Scholar] [CrossRef] [Green Version]
- Szczepaniec, A.; Finke, D. Plant-Vector-Pathogen Interactions in the Context of Drought Stress. Front. Ecol. Evol. 2019, 7, 262. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, E.; Cutrona, C.; del Toro, F.J.; Vallarino, J.G.; Osorio, S.; Pérez-Bueno, M.L.; Barón, M.; Chung, B.N.; Canto, T.; Tenllado, F. Virulence determines beneficial trade-offs in the response of virus-infected plants to drought via induction of salicylic acid. Plant Cell Environ. 2017, 40, 2909–2930. [Google Scholar] [CrossRef]
- Corrales-Gutierrez, M.; Medina-Puche, L.; Yu, Y.; Wang, L.; Ding, X.; Luna, A.P.; Bejarano, E.R.; Castillo, A.G.; Lozano-Duran, R. The C4 protein from the geminivirus Tomato yellow leaf curl virus confers drought tolerance in Arabidopsis through an ABA-independent mechanism. Plant Biotechnol. J. 2019, 18, 1121–1123. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.S.; Bosque-Pérez, N.A.; Foote, N.E.; Magney, T.; Eigenbrode, S.D. Environmentally dependent host–pathogen and vector–pathogen interactions in the Barley yellow dwarf virus pathosystem. J. Appl. Ecol. 2015, 52, 1392–1401. [Google Scholar] [CrossRef]
- Fernández-Calvino, L.; Osorio, S.; Hernández, M.L.; Hamada, I.B.; del Toro, F.J.; Donaire, L.; Yu, A.; Bustos, R.; Fernie, A.R.; Martinez-Rivas, J.M.; et al. Virus-induced alterations in primary metabolism modulate susceptibility to Tobacco rattle virus in Arabidopsis. Plant Physiol. 2014, 166, 1821–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, J.H.; Mccann, L.; Naish, M.; Dixon, H.; Murphy, A.M.; Stancombe, M.A.; Carr, J.P. A viral RNA silencing suppressor interferes with abscisic acid-mediated signalling and induces drought tolerance in Arabidopsis thaliana. Mol. Plant Pathol. 2013, 14, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef]
- Bueso, E.; Serrano, R.; Pallás, V.; Sánchez-Navarro, J.A. Seed tolerance to deterioration in Arabidopsis is affected by virus infection. Plant Physiol. Biochem. 2017, 116, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Achuo, E.A.; Prinsen, E.; Hofle, M. Influence of drought; salt stress and abscisic acid on the resistance of tomato to Botrytis cinerea and Oidium neolycopersici. Plant Pathol. 2006, 55, 178–186. [Google Scholar] [CrossRef]
- Ramegowda, V.; Senthil-Kumar, M.; Ishiga, Y.; Kaundal, A.; Udayakumar, M.; Mysore, K.S. Drought stress acclimation impacts tolerance to Sclerotina sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana. Int. J. Mol. Sci. 2013, 14, 9497–9513. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Ren, Q.; Sun, Y.; Ye, L.; Cao, H.; Ge, F. Lower incidence and severity of Tomato virus in elevated CO2 was accompanied by modulated plant induced defense in tomato. Plant Biol. 2012, 14, 905–913. [Google Scholar] [CrossRef]
- Ye, L.F.; Fu, X.; Ge, F. Elevated CO2 alleviates damage from Potato virus Y infection in tobacco plants. Plant Sci. 2010, 179, 219–224. [Google Scholar] [CrossRef]
- Zhang, S.; Li, X.; Sun, Z.; Shao, S.; Hu, L.; Ye, M.; Zhou, Y.; Xia, X.; Yu, J.; Shi, K. Antagonism between phytohormone signalling underlies the variation in disease susceptibility of tomato plants under elevated CO2. J. Exp. Bot. 2015, 66, 1951–1963. [Google Scholar] [CrossRef] [Green Version]
- Bilgin, D.D.; Aldea, M.; O’Neill, B.F.; Benitez, M.; Li, M.; Clough, S.J.; De Lucia, E.H. Elevated ozone alters soybean-virus interaction. Mol. Plant-Microbe Interact. 2008, 21, 1297–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hull, R. Disease symptoms and host range. In Matthew’s Plant Virology, 4th ed.; Elsevier: San Diego, CA, USA, 2002; pp. 47–74. [Google Scholar]
- Bergès, S.E.; Vile, D.; Vazquez-Rovere, C.; Blanc, S.; Yvon, M.; Bédiée, A.; Rolland, G.; Dauzat, M.; van Munster, M. Interactions between Drought and Plant Genotype Change Epidemiological Traits of Cauliflower mosaic virus. Front. Plant Sci. 2018, 9, 703. [Google Scholar] [CrossRef]
- Bergès, S.E.; Vasseur, F.; Bediée, A.; Rolland, G.; Masclef, D.; Dauzat, M.; van Munster, M.; Vile, D. Natural variation of Arabidopsis thaliana responses to Cauliflower mosaic virus infection upon water deficit. PLoS Pathog. 2020, 16, e1008557. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.P. Exploring how viruses enhance plants’ resilience to drought and the limits to this form of viral payback. Plant Cell Environ. 2017, 40, 2906–2908. [Google Scholar] [CrossRef] [PubMed]
- Molina-Montenegro, M.A.; Salgado-Luarte, C.; Oses, R.; Torres-Díaz, C. Is Physiological Performance a Good Predictor for Fitness? Insights from an Invasive Plant Species. PLoS ONE 2013, 8, e76432. [Google Scholar] [CrossRef] [Green Version]
- Pagán, I.; Fraile, A.; Fernández-Fueyo, E.; Montes, N.; Alonso-Blanco, C.; García-Arenal, F. Arabidopsis thaliana as a model for the study of plant–virus co-evolution. Philos. Trans. R. Soc. Lond. B 2010, 365, 1983–1995. [Google Scholar] [CrossRef] [Green Version]
- Schiavinato, M.; Marcet-Houben, M.; Dohm, J.C.; Gabaldón, T.; Himmelbauer, H. Parental origin of the allotetraploid tobacco Nicotiana benthamiana. Plant J. 2020, 102, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Montes, N.; Pagán, I. Light intensity modulates the efficiency of virus seed transmission through modifications of plant tolerance. Plants 2019, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- González, R.; Butković, A.; Elena, S.F. From foes to friends: Viral infections expand the limits of host phenotypic plasticity. Adv. Virus Res. 2020, 106, 85–121. [Google Scholar]
- Murray, R.R.; Emblow, M.S.M.; Hetherington, A.M.; Foster, G.D. Plant virus infections control stomatal development. Sci. Rep. 2016, 6, 34507. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, F.; Manrique, P.; Mansilla, C.; Lunello, P.; Wang, X.; Rodrigo, G.; López-González, S.; Jenner, C.; González-Melendi, P.; Elena, S.F.; et al. Viral strain-specific differential alterations in Arabidopsis developmental patterns. Mol. Plant-Microbe Interact. 2015, 28, 1304–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobos, A.; Montes, N.; López-Herranz, M.; Gil-Valle, M.; Pagán, I. Within-host multiplication and speed of colonization as infection traits associated with plant virus vertical transmission. J. Virol. 2019, 93, e01078-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alazem, M.; Lin, N.S. Antiviral Roles of Abscisic Acid in Plants. Front. Plant Sci. 2017, 8, 1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasin, F.; Shan, H.; García, B.; Muller, M.; San León, D.; Ludman, M.; Fresno, D.H.; Fátyol, K.; Munné-Bosch, S.; Rodrigo, G.; et al. Abscisic Acid Connects Phytohormone Signaling with RNA Metabolic Pathways and Promotes an Antiviral Response that Is Evaded by a Self-Controlled RNA Virus. Plant Commun. 2020, 1, 100099. [Google Scholar] [CrossRef]
- Carr, J.P.; Gruden, K. An update on salicylic acid biosynthesis, its induction and potential exploitation by plant viruses. Curr. Opin. Virol. 2020, 42, 8–17. [Google Scholar]
- Alazem, M.; Lin, K.Y.; Lin, N.S. The abscisic acid pathway has multifaceted effects on the accumulation of Bamboo mosaic virus. Mol. Plant Microbe Interact. 2014, 27, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Carr, J.P.; Murphy, A.M.; Tungadi, T.; Yoon, J.Y. Plant defense signals: Players and pawns in plant-virus-vector interactions. Plant Sci. 2019, 279, 87–95. [Google Scholar] [CrossRef]
- Montes, N.; Vijayan, V.; Pagán, I. Trade-offs between host tolerances to different pathogens in plant-virus interactions. Virus Evol. 2020, 6, veaa019. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, E.; Lozano-Duran, R. Plant viruses as probes to engineer tolerance to abiotic stress in crops. Stress Biol. 2022, 2. [Google Scholar] [CrossRef]
- Sastry, K.S. Seed-Borne Plant Virus Diseases; Springer: New Delhi, India, 2013. [Google Scholar]
- Roossinck, M.J.; Palukaitis, P. Rapid induction and severity of symptoms in zucchini squash (Cucurbita pepo) map to RNA 1 of Cucumber mosaic virus. Mol. Plant-Microbe Interact. 1990, 3, 188–192. [Google Scholar] [CrossRef]
- Dunoyer, P.; Thomas, C.; Harrison, S.; Revers, F.; Maule, A. A cysteine-rich plant protein potentiates Potyvirus movement through an interaction with the virus genome-linked protein VPg. J. Virol. 2004, 78, 2301–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, F.; Martínez-Herrera, D.; Aguilar, I.; Ponz, F. Infectivity of turnip mosaic potyvirus cDNA clones and transcripts on the systemic host Arabidopsis thaliana and local lesion hosts. Virus Res. 1998, 55, 207–219. [Google Scholar] [CrossRef]
- Tenllado, F.; Díaz-Ruíz, J.R. Double-Stranded RNA-Mediated Interference with Plant Virus Infection. J. Virol. 2001, 75, 12288–12297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasin, F.; Simón-Mateo, C.; García, J.A. The Hypervariable Amino-Terminus of P1 Protease Modulates Potyviral Replication and Host Defense Responses. PLoS Pathog. 2014, 10, e1003985. [Google Scholar] [CrossRef]
- Miura, K.; Okamoto, H.; Okuma, E.; Shiba, H.; Kamada, H.; Hasegawa, P.M.; Murata, Y. SIZ1 deficiency causes reduced stomatal aperture and enhanced drought tolerance via controlling salicylic acid-induced accumulation of reactive oxygen species in Arabidopsis. Plant J. 2013, 73, 91–104. [Google Scholar] [CrossRef]
- Del Toro, F.J.; Aguilar, E.; Hernández-Walias, F.J.; Tenllado, F.; Chung, B.N.; Canto, T. High temperature, high ambient CO2 affect the interactions between three positive-sense RNA viruses and a compatible host differentially, but not their silencing suppression efficiencies. PLoS ONE 2015, 10, e0136062. [Google Scholar] [CrossRef] [Green Version]
- García-Marcos, A.; Pacheco, R.; Manzano, A.; Aguilar, E.; Tenllado, F. Oxylipin biosynthesis genes positively regulate programmed cell death during compatible infections with the synergistic pair Potato virus X-Potato virus Y and Tomato spotted wilt virus. J. Virol. 2013, 87, 5769–5783. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, M.; Ojeda, B.; Hernández-Walias, F.J.; Sanz-García, E.; Canto, T.; Tenllado, F. Water Deficit Improves Reproductive Fitness in Nicotiana benthamiana Plants Infected by Cucumber mosaic virus. Plants 2022, 11, 1240. https://doi.org/10.3390/plants11091240
Moreno M, Ojeda B, Hernández-Walias FJ, Sanz-García E, Canto T, Tenllado F. Water Deficit Improves Reproductive Fitness in Nicotiana benthamiana Plants Infected by Cucumber mosaic virus. Plants. 2022; 11(9):1240. https://doi.org/10.3390/plants11091240
Chicago/Turabian StyleMoreno, Marina, Belén Ojeda, Francisco J. Hernández-Walias, Eugenio Sanz-García, Tomás Canto, and Francisco Tenllado. 2022. "Water Deficit Improves Reproductive Fitness in Nicotiana benthamiana Plants Infected by Cucumber mosaic virus" Plants 11, no. 9: 1240. https://doi.org/10.3390/plants11091240