
Grain-Priming with L-Arginine Improves the Growth Performance of Wheat (Triticum aestivum L.) Plants under Drought Stress

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Growth Parameters
2.1.1. Plant Height, Root Length, Tillers Number, Leaves Number, and Flag Leaf Area
2.1.2. Fresh and Dry Weights of Shoot and Root
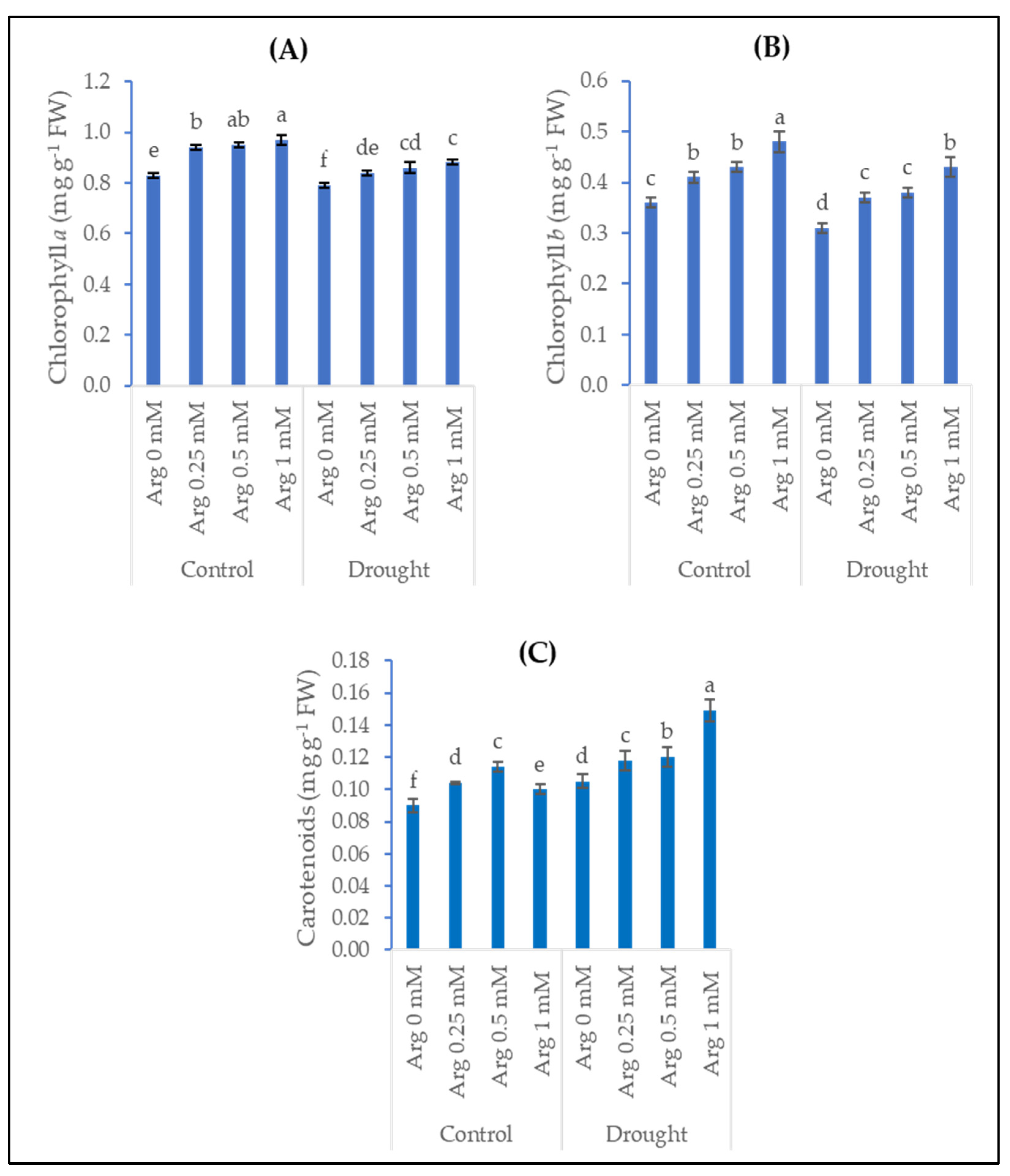
2.2. Photosynthetic Pigments
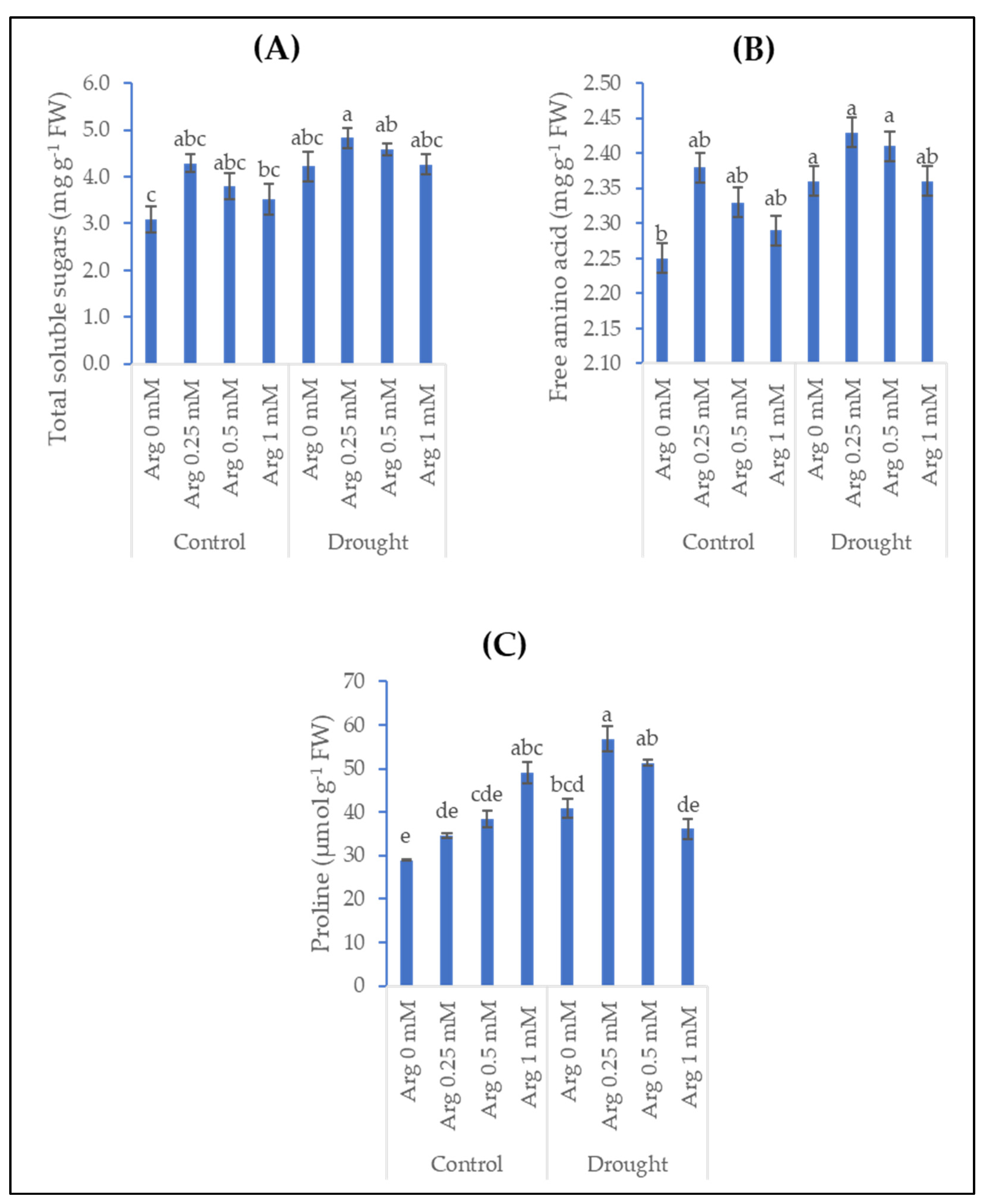
2.3. Osmo-Protectants (Soluble Sugars, Free Amino Acids, Free Proline)
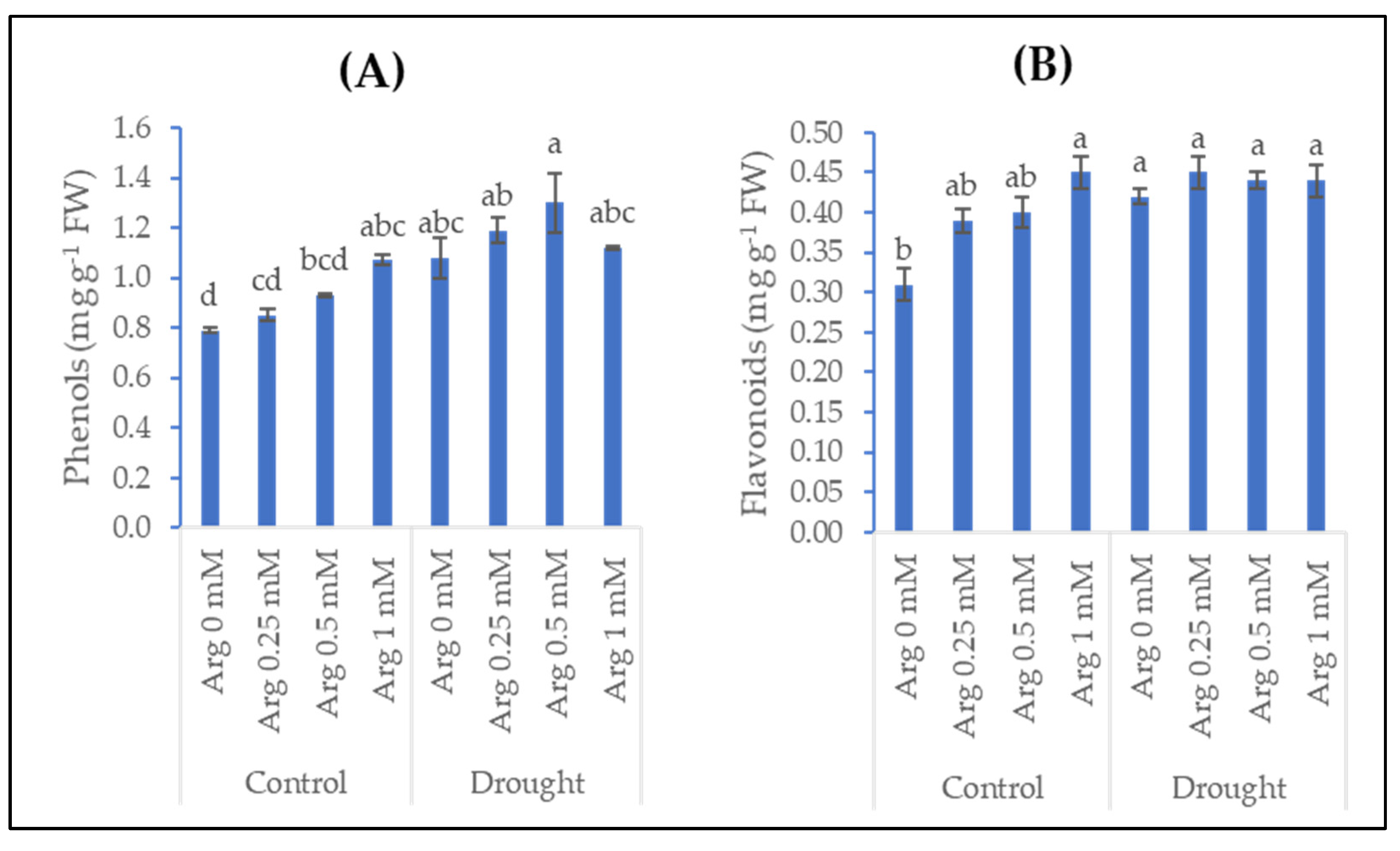
2.4. Phenols and Flavonoids
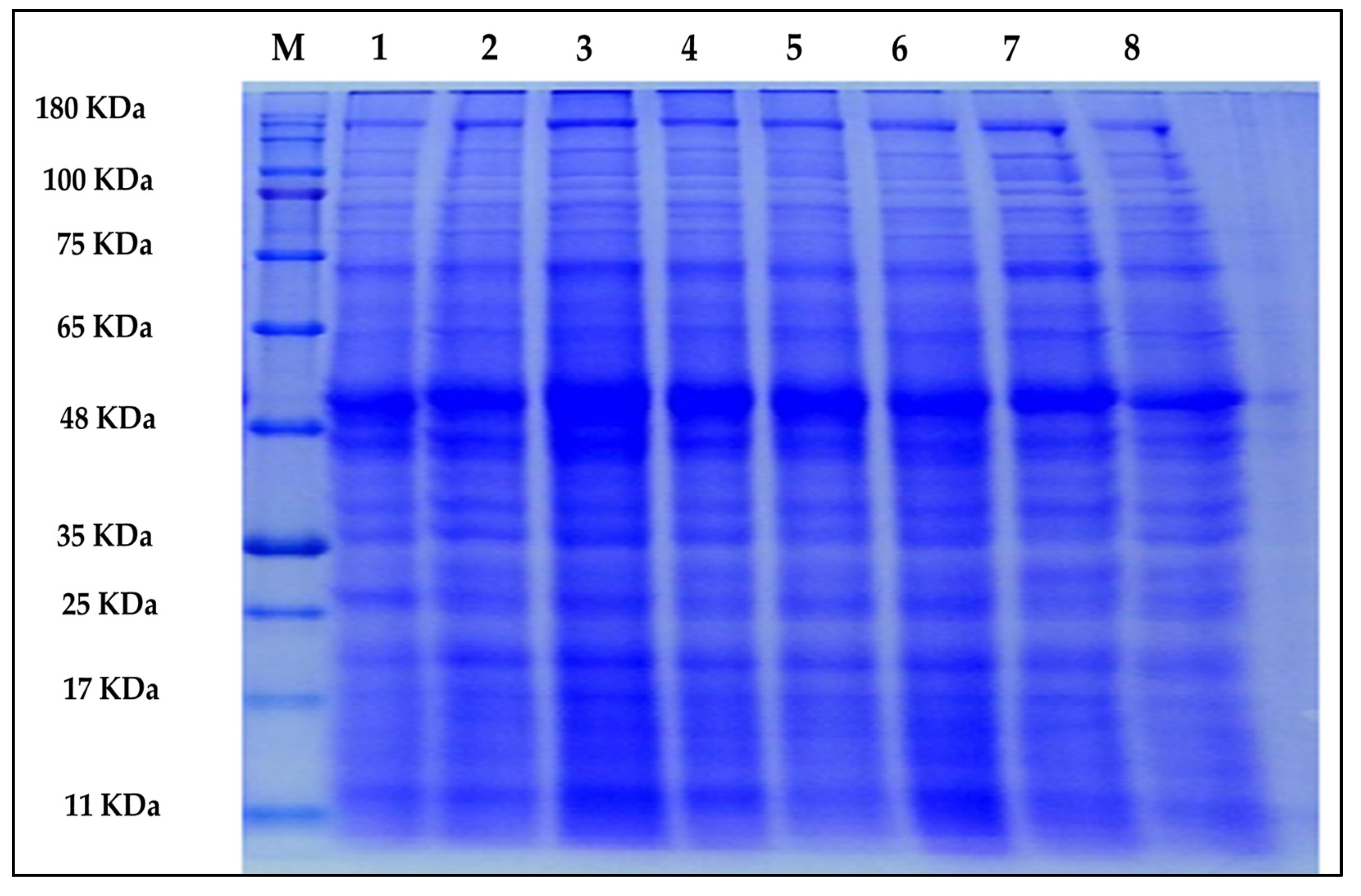
2.5. Protein Profile
3. Discussion
4. Materials and Methods
4.1. Experiment Design
4.2. Morphological Growth Traits
4.3. Photosynthetic Pigments
4.4. Total Soluble Sugars
4.5. Total Free Amino Acids
4.6. Free Proline
4.7. Total Phenolics
4.8. Total Flavonoids
4.9. Protein Profile
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iqbal, S.; Wang, X.; Mubeen, I.; Kamran, M.; Kanwal, I.; Díaz, G.A.; Abbas, A.; Parveen, A.; Atiq, M.N.; Alshaya, H.; et al. Phytohormones trigger drought tolerance in crop plants: Outlook and future perspectives. Front. Plant Sci. 2022, 12, 799318. [Google Scholar] [CrossRef] [PubMed]
- Ewais, E.A.; Ismail, M.A.; Amin, M.A.; Badawy, A.A. Efficiency of salicylic acid and glycine on sugar beet plants grown under heavy metals pollution. Egypt. J. Biotechnol. 2015, 48, 112–126. [Google Scholar]
- Hussein, H.-A.A. Influence of radio-grain priming on growth, antioxidant capacity, and yield of barley plants. Biotechnol. Rep. 2022, 34, e00724. [Google Scholar] [CrossRef]
- Gebaly, S.; Ahmed, F.; Namich, A. Effect of Spraying Some Organic, Amino Acids and Potassium Citrate on Alleviation of Drought Stress in Cotton Plant. J. Plant Prod. 2013, 4, 1369–1381. [Google Scholar] [CrossRef]
- Hu, W.; Ren, T.; Meng, F.; Cong, R.; Li, X.; White, P.J.; Lu, J. Leaf photosynthetic capacity is regulated by the interaction of nitrogen and potassium through coordination of CO2 diffusion and carboxylation. Physiol. Plant. 2019, 167, 418–432. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Kasim, W.A.; Hamada, E.A.M.; El-din, N.G.S.; Eskander, S. Influence of seaweed extracts on the growth, some metabolic activities and yield of wheat grown under drought stress. Int. J. Agron. Agric. Res. 2015, 7, 173–189. [Google Scholar]
- Hussain, H.A.; Men, S.; Hussain, S.; Zhang, Q.; Ashraf, U.; Anjum, S.A.; Ali, I.; Wang, L. Maize tolerance against drought and chilling stresses varied with root morphology and antioxidative defense system. Plants 2020, 9, 720. [Google Scholar] [CrossRef]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef]
- Johnson, R.; Puthur, J.T. Seed priming as a cost effective technique for developing plants with cross tolerance to salinity stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef]
- Nejadalimoradi, H.; Nasibi, F.; Kalantari, K.M.; Zanganeh, R. Effect of seed priming with L-arginine and sodium nitroprusside on some physiological parameters and antioxidant enzymes of sunflower plants exposed to salt stress. Agric. Commun. 2014, 2, 23–30. [Google Scholar]
- Osman, M.S.; Badawy, A.A.; Osman, A.I.; Abdel Latef, A.A.H. Ameliorative impact of an extract of the halophyte Arthrocnemum macrostachyum on growth and biochemical parameters of soybean under salinity stress. J. Plant Growth Regul. 2021, 40, 1245–1256. [Google Scholar] [CrossRef]
- Hernández-Hernández, H.; Quiterio-Gutiérrez, T.; Cadenas-Pliego, G.; Ortega-Ortiz, H.; Hernández-Fuentes, A.D.; de la Fuente, M.; Valdés-Reyna, J.; Juárez-Maldonado, A. Impact of Selenium and Copper Nanoparticles on Yield, Antioxidant System, and Fruit Quality of Tomato Plants. Plants 2019, 8, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, M.A.; Ismail, M.A.; Badawy, A.A.; Awad, M.A.; Hamza, M.F.; Awad, M.F.; Fouda, A. The Potency of fungal-fabricated selenium nanoparticles to improve the growth performance of Helianthus annuus L. and control of cutworm Agrotis ipsilon. Catalysts 2021, 11, 1551. [Google Scholar] [CrossRef]
- Badawy, A.A.; Abdelfattah, N.A.H.; Salem, S.S.; Awad, M.F.; Fouda, A. Efficacy assessment of biosynthesized copper oxide nanoparticles (CuO-NPs) on stored grain insects and their impacts on morphological and physiological traits of wheat (Triticum aestivum L.) plant. Biology 2021, 10, 233. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Hamid, Z.H.; Amer, M.; Wahab, A. Effect of arginine on growth and yield of tomato plant (Lycopersicon esculentum) under drought stress. Plant Arch. 2019, 19, 4441–4444. [Google Scholar]
- El-Bassiouny, H.M.; Mostafa, H.A.; El-Khawas, S.A.; Hassanein, R.A.; Khalil, S.I.; Abd El-Monem, A.A. Physiological Responses of Wheat Plant to Foliar Treatments with Arginine or Putrescine. Aust. J. Basic Appl. Sci. 2008, 2, 1390–1403. [Google Scholar]
- Nasibi, F.; Yaghoobi, M.M.; Kalantari, K.M. Effect of exogenous arginine on alleviation of oxidative damage in tomato plant underwater stress. J. Plant Interact. 2011, 6, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matysiak, K.; Kierzek, R.; Siatkowski, I.; Kowalska, J.; Krawczyk, R.; Miziniak, W. Effect of exogenous application of amino acids L-Arginine and glycine on maize under temperature stress. Agronomy 2020, 10, 769. [Google Scholar] [CrossRef]
- Nassar, A.H.; El-Tarabily, K.A.; Sivasithamparam, K. Growth promotion of bean (Phaseolus vulgaris L.) by a polyamine-producing isolate of Streptomyces griseoluteus. Plant Growth Regul. 2003, 40, 97–106. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.; Jha, P.N. The plant-growth-promoting bacterium Klebsiella sp. SBP-8 confers induced systemic tolerance in wheat (Triticum aestivum) under salt stress. J. Plant Physiol. 2015, 184, 57–67. [Google Scholar] [CrossRef]
- Jing, J.; Guo, S.; Li, Y.; Li, W. The alleviating effect of exogenous polyamines on heat stress susceptibility of different heat resistant wheat (Triticum aestivum L.) varieties. Sci. Rep. 2020, 10, 7467. [Google Scholar] [CrossRef]
- Abdullah, A.S.; Aziz, M.M.; Siddique, K.H.M.; Flower, K.C. Film antitranspirants increase yield in drought stressed wheat plants by maintaining high grain number. Agric. Water Manag. 2015, 159, 11–18. [Google Scholar] [CrossRef]
- Hammad, S.A.R.; Ali, O.A.M. Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Ann. Agric. Sci. 2014, 59, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Asif, I.; Qiang, D.; Xiangru, W.; Huiping, G.; Hengheng, Z.; Xiling, Z.; Meizhen, S. High nitrogen enhance drought tolerance in cotton through antioxidant enzymatic activities, nitrogen metabolism and osmotic adjustment. Plants 2020, 9, 178. [Google Scholar]
- Saikia, J.; Sarma, R.K.; Dhandia, R.; Yadav, A.; Bharali, R.; Gupta, V.K.; Saikia, R. Alleviation of drought stress in pulse crops with ACC deaminase producing rhizobacteria isolated from acidic soil of Northeast India. Sci. Rep. 2018, 8, 3560. [Google Scholar] [CrossRef]
- Faraji, J.; Sepehri, A. Ameliorative effects of TiO2 nanoparticles and sodium nitroprusside on seed germination and seedling growth of wheat under PEG-stimulated drought stress. J. Seed Sci. 2019, 41, 309–317. [Google Scholar] [CrossRef]
- Shalaby, M.A.F.; Ahmed, M.A.; Khater, M.A. Physiological responses of some barley cultivars to foliar treatments with arginine under water stress conditions. Middle East J. Agric. Res. 2018, 7, 1102–1123. [Google Scholar]
- Ahmed, M.A.; Shalaby, M.A.F.; El-Housini, E.A.; Khater, M.A. Alleviation of drought stress on fenugreek (Trigonella foenum-graecum L.) plants by foliar application of polyamines compounds. Middle East J. Appl. Sci. 2018, 8, 883–894. [Google Scholar]
- Hanafy, R.S. Using Moringa olifera leaf extract as a bio-fertilizer for drought stress mitigation of Glycine max L. plants. Egypt. J. Bot. 2017, 57, 281–292. [Google Scholar] [CrossRef]
- Hellal, F.; Abdel-Hady, M.; Khatab, I.; El-Sayed, S.; Abdelly, C. Yield characterization of Mediterranean barley under drought stress condition. AIMS Agric. Food 2019, 4, 518–533. [Google Scholar] [CrossRef]
- Bogati, K.; Walczak, M. The impact of drought stress on soil microbial community, enzyme activities and plants. Agronomy 2022, 12, 189. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Ramadan, A.A.; Abd Elhamid, E.M.; Sadak, M.S. Comparative study for the effect of arginine and sodium nitroprusside on sunflower plants grown under salinity stress conditions. Bull. Natl. Res. Cent. 2019, 43, 118. [Google Scholar] [CrossRef]
- García, J.E.; Maroniche, G.; Creus, C.; Suárez-rodríguez, R.; Ramirez-trujillo, J.A.; Groppa, M.D. In vitro PGPR properties and osmotic tolerance of different Azospirillum native strains and their effects on growth of maize under drought stress. Microbiol. Res. 2017, 202, 21–29. [Google Scholar] [CrossRef]
- Ahmed, A.G.; Zaki, N.M.; Tawfik, M.H.M.M.M.; Hassanein, M.S. Growth and yield response of two chickpea cultivars (Cicer arietinum L.) to skipping one irrigation. Middle East J. Agri. Res 2013, 2, 146–151. [Google Scholar]
- El-sayed, S.; Ramadan, A.A.E.; Hellal, F. Drought stress mitigation by application of algae extract on peanut grown under sandy soil conditions. Asian J. Plant Sci. 2020, 19, 230–239. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Exogenous nitric oxide donor and arginine provide protection against short-term drought stress in wheat seedlings. Physiol. Mol. Biol. Plants 2018, 24, 993–1004. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.; Farooq, M.; Xue, L.; Ali, S. Fulvic acid application improves the maize performance under well-watered and drought conditions. J. Agron. Crop Sci. 2011, 197, 409–417. [Google Scholar] [CrossRef]
- Marcińska, I.; Czyczyło-Mysza, I.; Skrzypek, E.; Filek, M.; Grzesiak, S.; Grzesiak, M.T.; Janowiak, F.; Hura, T.; Dziurka, M.; Dziurka, K.; et al. Impact of osmotic stress on physiological and biochemical characteristics in drought-susceptible and drought-resistant wheat genotypes. Acta Physiol. Plant. 2013, 35, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Nasibi, F.; Heidari, T.; Asrar, Z.; Mansoori, H. Effect of arginine pre-treatment on nickel accumulation and alleviation of the oxidative stress in Hyoscyamus niger. J. Soil Sci. Plant Nutr. 2013, 13, 680–689. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Liang, Z.; Shao, H.; Du, F. Effect of water deficits on the activity of anti-oxidative enzymes and osmoregulation among three different genotypes of Radix Astragali at seeding stage. Colloids Surf. B Biointerfaces 2006, 49, 60–65. [Google Scholar] [CrossRef]
- Badawy, A.A.; Alotaibi, M.O.; Abdelaziz, A.M.; Osman, M.S.; Khalil, A.M.A.; Saleh, A.M.; Mohammed, A.E.; Hashem, A.H. Enhancement of seawater stress tolerance in barley by the endophytic fungus Aspergillus ochraceus. Metabolites 2021, 11, 428. [Google Scholar] [CrossRef]
- Iqbal, S. Physiology of Wheat (Triticum aestivum L.) Accessions and the Role of Phytohormones under Water Stress; Quaid-I-Azam University: Islamabad, Pakistan, 2009. [Google Scholar]
- Abdel Latef, A.A.H.; Omer, A.M.; Badawy, A.A.; Osman, M.S.; Ragaey, M.M. Strategy of salt tolerance and interactive impact of Azotobacter chroococcum and/or Alcaligenes faecalis inoculation on canola (Brassica napus L.) plants grown in saline soil. Plants 2021, 10, 110. [Google Scholar] [CrossRef]
- Hammad, S.A.R.; El-Gamal, S.M. Response of pepper plants grown under water stress condition to biofertilizers (Halex 2) and mineral nitrogen. Minufiya J. Agric. Res 2004, 29, 1–27. [Google Scholar]
- Feiz, F.S.; Hakimi, L.; Mousavi, A.; Jahromi, M.G. The effects of glycine betaine and L-arginine on biochemical properties of pot marigold (Calendula officinalis L.) under water stress. Iran. J. Plant Physiol. 2019, 9, 2795–2805. [Google Scholar]
- Liu, J.H.; Kitashiba, H.; Wang, J.; Ban, Y.; Moriguchi, T. Polyamines and their ability to provide environmental stress tolerance to plants. Plant Biotechnol. 2007, 24, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Qados, A. Effect of arginine on growth, nutrient composition, yield and nutritional value of mung bean plants grown under salinity stress. Nature 2010, 8, 30–42. [Google Scholar]
- Hussein, H.-A.A.; Mekki, B.B.; Abd El-Sadek, M.E.; El Lateef, E.E. Effect of L-Ornithine application on improving drought tolerance in sugar beet plants. Heliyon 2019, 5, e02631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Liu, Q.; Wang, N.; Dai, J.; Lu, Q.; Jia, X.; Lin, L.; Yu, F.; Zuo, Y. Foliar arginine application improves tomato plant growth, yield, and fruit quality via nitrogen accumulation. Plant Growth Regul. 2021, 95, 421–428. [Google Scholar] [CrossRef]
- Metzner, H.; Rau, H.; Senger, H. Untersuchungen zur Synchronisierbarkeit einzelner Pigmentmangel-Mutanten von Chlorella. Planta 1965, 65, 186–194. [Google Scholar] [CrossRef]
- Cerning, B.J. A note on sugar determination by the anthrone method. Cereal Chem. 1975, 52, 857–860. [Google Scholar]
- Rosen, H. A modified ninhydrin colorimetric analysis for amino acids. Arch. Biochem. Biophys. 1957, 67, 10–15. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Savitree, M.; Isara, P.; Nittaya, S.L.; Worapan, S. Radical scavenging activity and total phenolic content of medicinal plants used in primary health care. J. Pharm. Sci. 2004, 9, 32–35. [Google Scholar]
- Pourmorad, F.; Hosseinimehr, S.J.; Shahabimajd, N. Antioxidant activity, phenol and flavonoid contents of some selected Iranian medicinal plants. Afr. J. Biotechnol. 2006, 5, 1142–1145. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Darwesh, O.M.; Elshahawy, I.E. Silver nanoparticles inactivate sclerotial formation in controlling white rot disease in onion and garlic caused by the soil borne fungus Stromatinia cepivora. Eur. J. Plant Pathol. 2021, 160, 917–934. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods; The Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Root Length (cm) | Tillers Number | Leaves Number | Flag Leaf Area (cm2) | |
|---|---|---|---|---|---|---|
| Irrigation | Arginine | |||||
| Normal | 0 mM | 64.0 ± 2.65 b | 7.3 ± 0.33 bc | 2.3 ± 0.33 ab | 5.0 ± 0.58 cd | 18.3 ± 0.28 ab |
| 0.25 mM | 64.3 ± 0.33 b | 8.0 ± 0.58 bc | 2.0 ± 0.33 ab | 8.3 ± 0.33 abc | 18.6 ± 0.9 ab | |
| 0.5 mM | 67.7 ± 0.67 ab | 9.0 ± 0.58 b | 3.3 ± 0.33 ab | 10.7 ± 1.45 a | 19.3 ± 1.43 ab | |
| 1 mM | 73.0 ± 0.58 a | 11.7 ± 0.33 a | 4.0 ± 0.58 a | 9.7 ± 0.67 a | 22.7 ± 2.28 a | |
| Drought | 0 mM | 53.3 ± 0.88 c | 6.1 ± 0.35 c | 1.7 ± 0.32 b | 3.7 ± 0.33 d | 11.8 ± 0.53 c |
| 0.25 mM | 68.3 ± 1.47 ab | 7.7 ± 0.33 bc | 2.3 ± 0.3 ab | 5.0 ± 0.58 cd | 16.8 ± 0.03 bc | |
| 0.5 mM | 68.9 ± 1.45 ab | 8.3 ± 0.33 b | 2.3 ± 0.33 ab | 6.0 ± 0.58 bcd | 16.5 ± 1.32 bc | |
| 1 mM | 63.3 ± 1.42 b | 9.0 ± 0.12 b | 3.3 ± 0.31 ab | 8.7 ± 0.33 ab | 14.9 ± 0.92 bc | |
| Treatments | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Root Fresh Weight (g) | Root Dry Weight (g) | |
|---|---|---|---|---|---|
| Irrigation | Arginine | ||||
| Normal | 0 mM | 4.36 ± 0.3 cd | 1.12 ± 0.09 d | 0.92 ± 0.09 abc | 0.34 ± 0.02 bcd |
| 0.25 mM | 6.26 ± 0.37 bc | 1.72 ± 0.15 c | 1.08 ± 0.09 ab | 0.4 ± 0.03 abc | |
| 0.5 mM | 8.72 ± 1.33 ab | 2.44 ± 0.12 b | 1.02 ± 0.15 abc | 0.42 ± 0.02 ab | |
| 1 mM | 9.26 ± 0.49 a | 3.05 ± 0.16 a | 1.15 ± 0.05 a | 0.45 ± 0.01 a | |
| Drought | 0 mM | 2.85 ± 0.25 d | 1.1 ± 0.08 d | 0.37 ± 0.04 d | 0.15 ± 0.02 f |
| 0.25 mM | 4.61 ± 0.27 cd | 1.16 ± 0.07 d | 0.73 ± 0.04 bcd | 0.22 ± 0.01 ef | |
| 0.5 mM | 5.8 ± 0.08 c | 1.46 ± 0.06 cd | 0.71 ± 0.03 cd | 0.28 ± 0.02 de | |
| 1 mM | 6 ± 0.26 c | 1.58 ± 0.04 cd | 0.78 ± 0.01 bc | 0.31 ± 0.01 cde | |
| Band No. | M.W. (KDa) | Treatments | |||||||
|---|---|---|---|---|---|---|---|---|---|
| L1 | L2 | L3 | L4 | L5 | L6 | L7 | L8 | ||
| 1 | 180 | + | + | + | + | + | + | + | + |
| 2 | 125 | + | + | + | + | + | + | + | + |
| 3 | 122 | - | - | - | - | - | + | + | + |
| 4 | 100 | + | + | + | + | + | + | + | + |
| 5 | 80 | + | + | + | + | + | + | + | + |
| 6 | 68 | + | + | + | + | + | + | + | + |
| 7 | 66 | + | + | + | + | + | + | + | + |
| 8 | 65 | + | + | + | + | + | + | + | + |
| 9 | 64 | - | - | - | - | - | - | - | + |
| 10 | 63 | - | + | + | + | + | + | + | + |
| 11 | 61 | + | + | + | + | + | + | + | + |
| 12 | 60 | - | + | + | + | + | + | + | + |
| 13 | 54 | - | - | - | - | + | - | - | - |
| 14 | 52 | + | + | + | + | + | + | + | + |
| 15 | 48 | + | + | + | + | + | + | + | + |
| 16 | 37 | + | + | + | + | + | + | + | + |
| 17 | 35 | - | - | - | - | - | + | + | + |
| 18 | 34 | + | + | + | + | + | + | + | + |
| 19 | 33 | - | + | + | + | + | + | + | + |
| 20 | 31 | + | + | + | + | + | + | + | + |
| 21 | 29 | + | + | + | + | + | + | + | + |
| 22 | 26 | + | + | + | + | + | + | + | + |
| 23 | 23 | + | + | + | + | + | + | + | + |
| 24 | 21 | + | + | + | + | + | + | + | + |
| 25 | 20 | - | + | + | + | + | + | + | + |
| 26 | 18 | + | + | + | + | + | + | + | + |
| 27 | 17 | + | + | + | + | + | + | + | + |
| 28 | 16 | + | + | + | + | + | + | + | + |
| 29 | 15 | + | + | + | + | - | + | + | + |
| 30 | 14 | + | + | + | + | - | + | + | + |
| 31 | 12 | + | + | + | + | + | + | + | + |
| Total bands | 23 | 27 | 27 | 27 | 26 | 29 | 29 | 30 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, H.-A.A.; Alshammari, S.O.; Kenawy, S.K.M.; Elkady, F.M.; Badawy, A.A. Grain-Priming with L-Arginine Improves the Growth Performance of Wheat (Triticum aestivum L.) Plants under Drought Stress. Plants 2022, 11, 1219. https://doi.org/10.3390/plants11091219
Hussein H-AA, Alshammari SO, Kenawy SKM, Elkady FM, Badawy AA. Grain-Priming with L-Arginine Improves the Growth Performance of Wheat (Triticum aestivum L.) Plants under Drought Stress. Plants. 2022; 11(9):1219. https://doi.org/10.3390/plants11091219
Chicago/Turabian StyleHussein, Hebat-Allah A., Shifaa O. Alshammari, Sahar K. M. Kenawy, Fatma M. Elkady, and Ali A. Badawy. 2022. "Grain-Priming with L-Arginine Improves the Growth Performance of Wheat (Triticum aestivum L.) Plants under Drought Stress" Plants 11, no. 9: 1219. https://doi.org/10.3390/plants11091219