
Marker-Assisted Improvement of Bread Wheat Variety HD2967 for Leaf and Stripe Rust Resistance



Abstract
:1. Introduction
2. Results
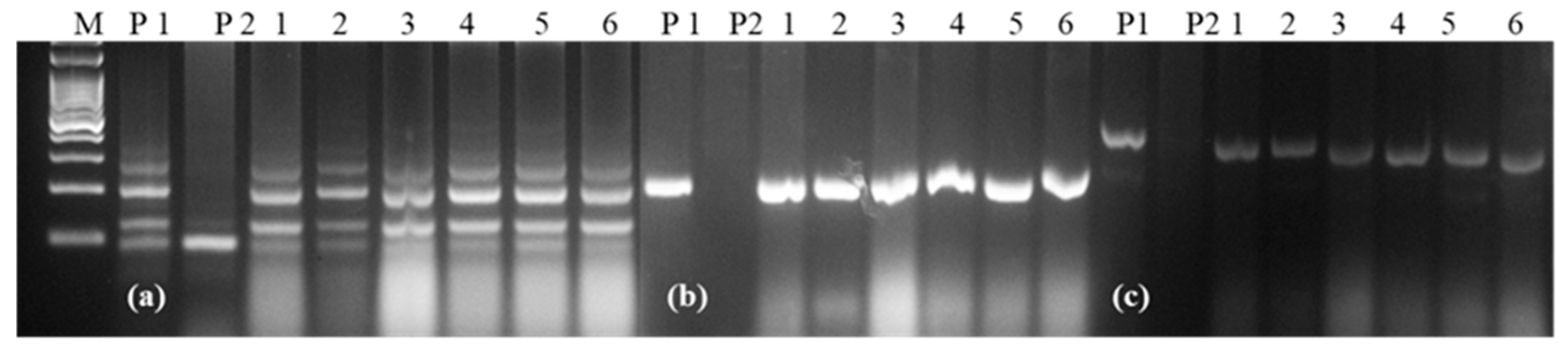
2.1. Development of NILs Carrying Leaf Rust Resistance Gene LrTrk and Leaf and Stripe Rust Resistance Gene LrTrk/YrTrk
2.2. Evaluation of HD2967 NILs for Yield-Related Traits
3. Discussion
4. Materials and Method
4.1. Plant Materials and Backcross Breeding Scheme
4.2. Marker Analysis
4.3. Screening of NILs for Rust Resistance
4.4. Evaluation of HD2967 + LrTrk NILs for Agro-Morphological Traits
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Marker | S.No. | Marker | S.No. | Marker | S.No. | Marker | S.No. | Marker |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Xbarc10 | 48 | Xbarc20 | 95 | Xbarc95 | 142 | Xcfd13 | 189 | Xcfd88 |
| 2 | Xbarc101 | 49 | Xbarc200 | 96 | Xbarc98 | 143 | Xcfd143 | 190 | Xgdm101 |
| 3 | Xbarc108 | 50 | Xbarc206 | 97 | Xcfa2019 | 144 | Xcfd15 | 191 | Xgdm109 |
| 4 | Xbarc109 | 51 | Xbarc21 | 98 | Xcfa2026 | 145 | Xcfd156 | 192 | Xgdm113 |
| 5 | Xbarc117 | 52 | Xbarc212 | 99 | Xcfa2028 | 146 | Xcfd16 | 193 | Xgdm116 |
| 6 | Xbarc119 | 53 | Xbarc229 | 100 | Xcfa2037 | 147 | Xcfd168 | 194 | Xgdm136 |
| 7 | Xbarc121 | 54 | Xbarc23 | 101 | Xcfa2040 | 148 | Xcfd170 | 195 | Xgdm14 |
| 8 | Xbarc123 | 55 | Xbarc232 | 102 | Xcfa2043 | 149 | Xcfd190 | 196 | Xgdm146 |
| 9 | Xbarc124 | 56 | Xbarc24 | 103 | Xcfa2056 | 150 | Xcfd193 | 197 | Xgdm28 |
| 10 | Xbarc127 | 57 | Xbarc240 | 104 | Xcfa2070 | 151 | Xcfd2 | 198 | Xgdm33 |
| 11 | Xbarc128 | 58 | Xbarc25 | 105 | Xcfa2076 | 152 | Xcfd2.1 | 199 | Xgdm36 |
| 12 | Xbarc13 | 59 | Xbarc267 | 106 | Xcfa2091 | 153 | Xcfd2.2 | 200 | Xgdm63 |
| 13 | Xbarc134 | 60 | Xbarc28 | 107 | Xcfa2104 | 154 | Xcfd20 | 201 | Xgpw2246 |
| 14 | Xbarc137 | 61 | Xbarc3 | 108 | Xcfa2106 | 155 | Xcfd219 | 202 | Xgpw3010 |
| 15 | Xbarc138 | 62 | Xbarc32 | 109 | Xcfa2110 | 156 | Xcfd22 | 203 | Xgpw3069 |
| 16 | Xbarc140 | 63 | Xbarc37 | 110 | Xcfa2114 | 157 | Xcfd24 | 204 | Xgpw3261 |
| 17 | Xbarc141 | 64 | Xbarc4 | 111 | Xcfa2121 | 158 | Xcfd242 | 205 | Xgpw5193 |
| 18 | Xbarc142 | 65 | Xbarc40 | 112 | Xcfa2123 | 159 | Xcfd25 | 206 | Xgpw7052 |
| 19 | Xbarc145 | 66 | Xbarc417 | 113 | Xcfa2129 | 160 | Xcfd251 | 207 | Xgpw7070 |
| 20 | Xbarc146 | 67 | Xbarc45 | 114 | Xcfa2134 | 161 | Xcfd257 | 208 | Xgpw7072 |
| 21 | Xbarc147 | 68 | Xbarc48 | 115 | Xcfa2141 | 162 | Xcfd267 | 209 | Xgwm10 |
| 22 | Xbarc148 | 69 | Xbarc49 | 116 | Xcfa2147 | 163 | Xcfd28 | 210 | Xgwm107 |
| 23 | Xbarc151 | 70 | Xbarc5 | 117 | Xcfa2149 | 164 | Xcfd283 | 211 | Xgwm108 |
| 24 | Xbarc154 | 71 | Xbarc55 | 118 | Xcfa2155 | 165 | Xcfd30 | 212 | Xgwm11 |
| 25 | Xbarc158 | 72 | Xbarc56 | 119 | Xcfa2163 | 166 | Xcfd31 | 213 | Xgwm112 |
| 26 | Xbarc159 | 73 | Xbarc59 | 120 | Xcfa2164 | 167 | Xcfd36 | 214 | Xgwm113 |
| 27 | Xbarc163 | 74 | Xbarc60 | 121 | Xcfa2170 | 168 | Xcfd39 | 215 | Xgwm114 |
| 28 | Xbarc164 | 75 | Xbarc67 | 122 | Xcfa2174 | 169 | Xcfd4 | 216 | Xgwm120 |
| 29 | Xbarc165 | 76 | Xbarc68 | 123 | Xcfa2179 | 170 | Xcfd48 | 217 | Xgwm122 |
| 30 | Xbarc167 | 77 | Xbarc69 | 124 | Xcfa2183 | 171 | Xcfd5 | 218 | Xgwm124 |
| 31 | Xbarc17 | 78 | Xbarc7 | 125 | Xcfa2185 | 172 | Xcfd50 | 219 | Xgwm126 |
| 32 | Xbarc170 | 79 | Xbarc72 | 126 | Xcfa2187 | 173 | Xcfd54 | 220 | Xgwm129 |
| 33 | Xbarc173 | 80 | Xbarc73 | 127 | Xcfa2190 | 174 | Xcfd59 | 221 | Xgwm130 |
| 34 | Xbarc174 | 81 | Xbarc75 | 128 | Xcfa2191 | 175 | Xcfd6 | 222 | Xgwm131 |
| 35 | Xbarc176 | 82 | Xbarc76 | 129 | Xcfa2193 | 176 | Xcfd60 | 223 | Xgwm132 |
| 36 | Xbarc178 | 83 | Xbarc77 | 130 | Xcfa2219 | 177 | Xcfd62 | 224 | Xgwm133 |
| 37 | Xbarc18 | 84 | Xbarc78 | 131 | Xcfa2226 | 178 | Xcfd65 | 225 | Xgwm135 |
| 38 | Xbarc180 | 85 | Xbarc8 | 132 | Xcfa2234 | 179 | Xcfd7 | 226 | Xgwm136 |
| 39 | Xbarc181 | 86 | Xbarc80 | 133 | Xcfa2240 | 180 | Xcfd70 | 227 | Xgwm140 |
| 40 | Xbarc182 | 87 | Xbarc81 | 134 | Xcfa2250 | 181 | Xcfd71 | 228 | Xgwm146 |
| 41 | Xbarc183 | 88 | Xbarc83 | 135 | Xcfa2256 | 182 | Xcfd73 | 229 | Xgwm148 |
| 42 | Xbarc186 | 89 | Xbarc84 | 136 | Xcfa2257 | 183 | Xcfd74 | 230 | Xgwm149 |
| 43 | Xbarc187 | 90 | Xbarc85 | 137 | Xcfa2262 | 184 | Xcfd79 | 231 | Xgwm153 |
| 44 | Xbarc188 | 91 | Xbarc87 | 138 | Xcfa2278 | 185 | Xcfd80 | 232 | Xgwm154 |
| 45 | Xbarc195 | 92 | Xbarc89 | 139 | Xcfa2293 | 186 | Xcfd81 | 233 | Xgwm155 |
| 46 | Xbarc197 | 93 | Xbarc92 | 140 | Xcfd1 | 187 | Xcfd82 | 234 | Xgwm156 |
| 47 | Xbarc198 | 94 | Xbarc94 | 141 | Xcfd11 | 188 | Xcfd86 | 235 | Xgwm159 |
| 236 | Xgwm16 | 284 | Xgwm312 | 332 | Xgwm471 | 380 | Xgwm636 | 428 | Xwmc166 |
| 237 | Xgwm160 | 285 | Xgwm314 | 333 | Xgwm473 | 381 | Xgwm637 | 429 | Xwmc168 |
| 238 | Xgwm162 | 286 | Xgwm319 | 334 | Xgwm480 | 382 | Xgwm639 | 430 | Xwmc169 |
| 239 | Xgwm164 | 287 | Xgwm32 | 335 | Xgwm493 | 383 | Xgwm644 | 431 | Xwmc17 |
| 240 | Xgwm165 | 288 | Xgwm328 | 336 | Xgwm494 | 384 | Xgwm66 | 432 | Xwmc173 |
| 241 | Xgwm169 | 289 | Xgwm33 | 337 | Xgwm495 | 385 | Xgwm664 | 433 | Xwmc175 |
| 242 | Xgwm179 | 290 | Xgwm332 | 338 | Xgwm497 | 386 | Xgwm666 | 434 | Xwmc177 |
| 243 | Xgwm18 | 291 | Xgwm333 | 339 | Xgwm498 | 387 | Xgwm666.1 | 435 | Xwmc179 |
| 244 | Xgwm181 | 292 | Xgwm334 | 340 | Xgwm499 | 388 | Xgwm666.2 | 436 | Xwmc181 |
| 245 | Xgwm182 | 293 | Xgwm335 | 341 | Xgwm5 | 389 | Xgwm67 | 437 | Xwmc182 |
| 246 | Xgwm186 | 294 | Xgwm339 | 342 | Xgwm501 | 390 | Xgwm674 | 438 | Xwmc183 |
| 247 | Xgwm191 | 295 | Xgwm340 | 343 | Xgwm508 | 391 | Xgwm68 | 439 | Xwmc201 |
| 248 | Xgwm192 | 296 | Xgwm344 | 344 | Xgwm512 | 392 | Xgwm70 | 440 | Xwmc206 |
| 249 | Xgwm193 | 297 | Xgwm350 | 345 | Xgwm513 | 393 | Xgwm72 | 441 | Xwmc213 |
| 250 | Xgwm2 | 298 | Xgwm356 | 346 | Xgwm515 | 394 | Xgwm77 | 442 | Xwmc215 |
| 251 | Xgwm205 | 299 | Xgwm357 | 347 | Xgwm518 | 395 | Xgwm88 | 443 | Xwmc216 |
| 252 | Xgwm210 | 300 | Xgwm359 | 348 | Xgwm526 | 396 | Xgwm88.1 | 444 | Xwmc218 |
| 253 | Xgwm213 | 301 | Xgwm361 | 349 | Xgwm537 | 397 | Xgwm88.2 | 445 | Xwmc219 |
| 254 | Xgwm219 | 302 | Xgwm368 | 350 | Xgwm538 | 398 | Xgwm95 | 446 | Xwmc230 |
| 255 | Xgwm233 | 303 | Xgwm369 | 351 | Xgwm540 | 399 | Xgwm99 | 447 | Xwmc231 |
| 256 | Xgwm234 | 304 | Xgwm371 | 352 | Xgwm544 | 400 | Xwmc1 | 448 | Xwmc232 |
| 257 | Xgwm247 | 305 | Xgwm372 | 353 | Xgwm547 | 401 | Xwmc10 | 449 | Xwmc235 |
| 258 | Xgwm249 | 306 | Xgwm374 | 354 | Xgwm55 | 402 | Xwmc104 | 450 | Xwmc238 |
| 259 | Xgwm251 | 307 | Xgwm375 | 355 | Xgwm550 | 403 | Xwmc105 | 451 | Xwmc24 |
| 260 | Xgwm257 | 308 | Xgwm376 | 356 | Xgwm554 | 404 | Xwmc109 | 452 | Xwmc243 |
| 261 | Xgwm259 | 309 | Xgwm382 | 357 | Xgwm558 | 405 | Xwmc11 | 453 | Xwmc245 |
| 262 | Xgwm260 | 310 | Xgwm388 | 358 | Xgwm565 | 406 | Xwmc110 | 454 | Xwmc247 |
| 263 | Xgwm264 | 311 | Xgwm389 | 359 | Xgwm566 | 407 | Xwmc113 | 455 | Xwmc25 |
| 264 | Xgwm265 | 312 | Xgwm391 | 360 | Xgwm569 | 408 | Xwmc116 | 456 | Xwmc254 |
| 265 | Xgwm268 | 313 | Xgwm397 | 361 | Xgwm570 | 409 | Xwmc118 | 457 | Xwmc256 |
| 266 | Xgwm271 | 314 | Xgwm4 | 362 | Xgwm573 | 410 | Xwmc120 | 458 | Xwmc257 |
| 267 | Xgwm273 | 315 | Xgwm400 | 363 | Xgwm577 | 411 | Xwmc125 | 459 | Xwmc258 |
| 268 | Xgwm274 | 316 | Xgwm403 | 364 | Xgwm582 | 412 | Xwmc128 | 460 | Xwmc261 |
| 269 | Xgwm275 | 317 | Xgwm408 | 365 | Xgwm595 | 413 | Xwmc134 | 461 | Xwmc262 |
| 270 | Xgwm276 | 318 | Xgwm410 | 366 | Xgwm6 | 414 | Xwmc139 | 462 | Xwmc264 |
| 271 | Xgwm282 | 319 | Xgwm413 | 367 | Xgwm60 | 415 | Xwmc145 | 463 | Xwmc265 |
| 272 | Xgwm284 | 320 | Xgwm415 | 368 | Xgwm601 | 416 | Xwmc149 | 464 | Xwmc269 |
| 273 | Xgwm285 | 321 | Xgwm425 | 369 | Xgwm604 | 417 | Xwmc15 | 465 | Xwmc27 |
| 274 | Xgwm291 | 322 | Xgwm427 | 370 | Xgwm608 | 418 | Xwmc150 | 466 | Xwmc272 |
| 275 | Xgwm293 | 323 | Xgwm429 | 371 | Xgwm610 | 419 | Xwmc152 | 467 | Xwmc273 |
| 276 | Xgwm294 | 324 | Xgwm43 | 372 | Xgwm611 | 420 | Xwmc153 | 468 | Xwmc274 |
| 277 | Xgwm296 | 325 | Xgwm44 | 373 | Xgwm613 | 421 | Xwmc154 | 469 | Xwmc276 |
| 278 | Xgwm297 | 326 | Xgwm443 | 374 | Xgwm614 | 422 | Xwmc156 | 470 | Xwmc278 |
| 279 | Xgwm299 | 327 | Xgwm445 | 375 | Xgwm617 | 423 | Xwmc158 | 471 | Xwmc28 |
| 280 | Xgwm30 | 328 | Xgwm448 | 376 | Xgwm626 | 424 | Xwmc16 | 472 | Xwmc283 |
| 281 | Xgwm302 | 329 | Xgwm459 | 377 | Xgwm63 | 425 | Xwmc160 | 473 | Xwmc289 |
| 282 | Xgwm304 | 330 | Xgwm46 | 378 | Xgwm630 | 426 | Xwmc161 | 474 | Xwmc291 |
| 283 | Xgwm311 | 331 | Xgwm47 | 379 | Xgwm635 | 427 | Xwmc163 | 475 | Xwmc296 |
| 476 | Xwmc307 | 524 | Xwmc453 | 572 | Xwmc580 | 620 | Xwmc679 | 668 | Xwmc776 |
| 477 | Xwmc31 | 525 | Xwmc455 | 573 | Xwmc581 | 621 | Xwmc680 | 669 | Xwmc777 |
| 478 | Xwmc310 | 526 | Xwmc468 | 574 | Xwmc59 | 622 | Xwmc682 | 670 | Xwmc78 |
| 479 | Xwmc311 | 527 | Xwmc469 | 575 | Xwmc592 | 623 | Xwmc684 | 671 | Xwmc783 |
| 480 | Xwmc312 | 528 | Xwmc47 | 576 | Xwmc593 | 624 | Xwmc687 | 672 | Xwmc786 |
| 481 | Xwmc313 | 529 | Xwmc471 | 577 | Xwmc594 | 625 | Xwmc692 | 673 | Xwmc787 |
| 482 | Xwmc317 | 530 | Xwmc473 | 578 | Xwmc596 | 626 | Xwmc693 | 674 | Xwmc79 |
| 483 | Xwmc323 | 531 | Xwmc474 | 579 | Xwmc597 | 627 | Xwmc694 | 675 | Xwmc790 |
| 484 | Xwmc326 | 532 | Xwmc475 | 580 | Xwmc598 | 628 | Xwmc695 | 676 | Xwmc792 |
| 485 | Xwmc329 | 533 | Xwmc476 | 581 | Xwmc602 | 629 | Xwmc696 | 677 | Xwmc794 |
| 486 | Xwmc332 | 534 | Xwmc477 | 582 | Xwmc603 | 630 | Xwmc698 | 678 | Xwmc795 |
| 487 | Xwmc335 | 535 | Xwmc479 | 583 | Xwmc606 | 631 | Xwmc70 | 679 | Xwmc798 |
| 488 | Xwmc336 | 536 | Xwmc48 | 584 | Xwmc607 | 632 | Xwmc702 | 680 | Xwmc805 |
| 489 | Xwmc344 | 537 | Xwmc486 | 585 | Xwmc611 | 633 | Xwmc705 | 681 | Xwmc807 |
| 490 | Xwmc349 | 538 | Xwmc487 | 586 | Xwmc612 | 634 | Xwmc707 | 682 | Xwmc808 |
| 491 | Xwmc35 | 539 | Xwmc488 | 587 | Xwmc613 | 635 | Xwmc710 | 683 | Xwmc809 |
| 492 | Xwmc356 | 540 | Xwmc489 | 588 | Xwmc615 | 636 | Xwmc713 | 684 | Xwmc810 |
| 493 | Xwmc361 | 541 | Xwmc49 | 589 | Xwmc616 | 637 | Xwmc716 | 685 | Xwmc813 |
| 494 | Xwmc364 | 542 | Xwmc491 | 590 | Xwmc617 | 638 | Xwmc718 | 686 | Xwmc815 |
| 495 | Xwmc366 | 543 | Xwmc492 | 591 | Xwmc619 | 639 | Xwmc719 | 687 | Xwmc817 |
| 496 | Xwmc376 | 544 | Xwmc494 | 592 | Xwmc623 | 640 | Xwmc722 | 688 | Xwmc818 |
| 497 | Xwmc382 | 545 | Xwmc497 | 593 | Xwmc625 | 641 | Xwmc723 | 689 | Xwmc819 |
| 498 | Xwmc386 | 546 | Xwmc498 | 594 | Xwmc626 | 642 | Xwmc726 | 690 | Xwmc826 |
| 499 | Xwmc388 | 547 | Xwmc500 | 595 | Xwmc627 | 643 | Xwmc727 | 691 | Xwmc827 |
| 500 | Xwmc396 | 548 | Xwmc505 | 596 | Xwmc63 | 644 | Xwmc728 | 692 | Xwmc83 |
| 501 | Xwmc397 | 549 | Xwmc508 | 597 | Xwmc630 | 645 | Xwmc73 | 693 | Xwmc830 |
| 502 | Xwmc398 | 550 | Xwmc51 | 598 | Xwmc631 | 646 | Xwmc734 | 694 | Xwmc85 |
| 503 | Xwmc405 | 551 | Xwmc511 | 599 | Xwmc632 | 647 | Xwmc737 | 695 | Xwmc89 |
| 504 | Xwmc406 | 552 | Xwmc513 | 600 | Xwmc633 | 648 | Xwmc740 | 696 | Xwmc9 |
| 505 | Xwmc407 | 553 | Xwmc516 | 601 | Xwmc640 | 649 | Xwmc744 | 697 | Xwmc93 |
| 506 | Xwmc413 | 554 | Xwmc517 | 602 | Xwmc644 | 650 | Xwmc745 | 698 | Xwmc95 |
| 507 | Xwmc415 | 555 | Xwmc52 | 603 | Xwmc646 | 651 | Xwmc748 | 699 | Xwmc96 |
| 508 | Xwmc416 | 556 | Xwmc522 | 604 | Xwmc65 | 652 | Xwmc75 | 700 | Xwmc99 |
| 509 | Xwmc417 | 557 | Xwmc524 | 605 | Xwmc650 | 653 | Xwmc751 | ||
| 510 | Xwmc418 | 558 | Xwmc525 | 606 | Xwmc651 | 654 | Xwmc752 | ||
| 511 | Xwmc419 | 559 | Xwmc526 | 607 | Xwmc652 | 655 | Xwmc753 | ||
| 512 | Xwmc420 | 560 | Xwmc527 | 608 | Xwmc653 | 656 | Xwmc754 | ||
| 513 | Xwmc422 | 561 | Xwmc532 | 609 | Xwmc654 | 657 | Xwmc756 | ||
| 514 | Xwmc426 | 562 | Xwmc533 | 610 | Xwmc657 | 658 | Xwmc757 | ||
| 515 | Xwmc428 | 563 | Xwmc537 | 611 | Xwmc658 | 659 | Xwmc758 | ||
| 516 | Xwmc43 | 564 | Xwmc539 | 612 | Xwmc661 | 660 | Xwmc759 | ||
| 517 | Xwmc430 | 565 | Xwmc540 | 613 | Xwmc662 | 661 | Xwmc76 | ||
| 518 | Xwmc434 | 566 | Xwmc544 | 614 | Xwmc664 | 662 | Xwmc760 | ||
| 519 | Xwmc435 | 567 | Xwmc546 | 615 | Xwmc667 | 663 | Xwmc762 | ||
| 520 | Xwmc44 | 568 | Xwmc553 | 616 | Xwmc672 | 664 | Xwmc764 | ||
| 521 | Xwmc441 | 569 | Xwmc557 | 617 | Xwmc673 | 665 | Xwmc766 | ||
| 522 | Xwmc445 | 570 | Xwmc559 | 618 | Xwmc674 | 666 | Xwmc770 | ||
| 523 | Xwmc446 | 571 | Xwmc577 | 619 | Xwmc675 | 667 | Xwmc773 |
| S.No. | Markers | S.No. | Markers | S.No. | Markers | S.No. | Markers |
|---|---|---|---|---|---|---|---|
| 1 | Xbarc10 | 22 | Xcfa2170 | 43 | Xgwm155 | 64 | Xgwm573 |
| 2 | Xbarc128 | 23 | Xcfa2187 | 44 | Xgwm165 | 65 | Xgwm6 |
| 3 | Xbarc148 | 24 | Xcfa2193 | 45 | Xgwm186 | 66 | Xgwm60 |
| 4 | XBarc163 | 25 | Xcfa2262 | 46 | Xgwm191 | 67 | Xgwm613 |
| 5 | Xbarc197 | 26 | Xcfd13 | 47 | Xgwm192 | 68 | Xgwm63 |
| 6 | Xbarc212 | 27 | Xcfd193 | 48 | Xgwm2 | 69 | Xgwm635 |
| 7 | Xbarc229 | 28 | Xcfd20 | 49 | Xgwm234 | 70 | Xgwm66 |
| 8 | Xbarc23 | 29 | Xcfd242 | 50 | Xgwm249 | 71 | Xwmc11 |
| 9 | Xbarc232 | 30 | Xcfd39 | 51 | Xgwm251 | 72 | Xwmc247 |
| 10 | Xbarc417 | 31 | Xcfd48 | 52 | Xgwm294 | 73 | Xwmc291 |
| 11 | Xbarc69 | 32 | Xcfd6 | 53 | Xgwm304 | 74 | Xwmc311 |
| 12 | Xbarc73 | 33 | Xcfd71 | 54 | Xgwm328 | 75 | Xwmc317 |
| 13 | Xbarc83 | 34 | Xcfd88 | 55 | Xgwm332 | 76 | Xwmc417 |
| 14 | Xbarc98 | 35 | Xgdm63 | 56 | Xgwm334 | 77 | Xwmc420 |
| 15 | Xcfa2040 | 36 | Xgwm11 | 57 | Xgwm350 | 78 | Xwmc44 |
| 16 | Xcfa2076 | 37 | Xgwm126 | 58 | Xgwm382 | 79 | Xwmc473 |
| 17 | Xcfa2114 | 38 | Xgwm131 | 59 | Xgwm403 | 80 | Xwmc500 |
| 18 | Xcfa2121 | 39 | Xgwm148 | 60 | Xgwm46 | 81 | Xwmc748 |
| 19 | Xcfa2141 | 40 | Xgwm149 | 61 | Xgwm493 | 82 | Xwmc76 |
| 20 | Xcfa2155 | 41 | Xgwm153 | 62 | Xgwm495 | 83 | Xwmc807 |
| 21 | Xcfa2163 | 42 | Xgwm154 | 63 | Xgwm513 |
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Kolmer, J.A. Tracking wheat rust on a continental scale. Curr. Opin. Plant Biol. 2005, 8, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Tomar, S.M.S.; Singh, S.K.; Sivasamy, M.; Vinod. Wheat rusts in India: Resistance breeding and gene deployment—A review. Indian J. Genet. 2014, 74, 129–156. [Google Scholar] [CrossRef]
- Saari, E.E.; Prescott, J.M. World distribution in relation to economic losses. In Diseases, Distribution, Epidemiology, and Control; Academic Press: Cambridge, MA, USA, 1985; pp. 259–298. [Google Scholar]
- Bolton, M.D.; Kolmer, J.A.; Garvin, D.F. Wheat leaf rust caused by Puccinia triticina. Mol. Plant Pathol. 2008, 9, 563–575. [Google Scholar] [CrossRef]
- Huerta-Espino, J.; Singh, R.P.; German, S.; McCallum, B.D.; Park, R.F.; Chen, W.Q.; Bhardwaj, S.C.; Goyeau, H. Global status of wheat leaf rust caused by Puccinia triticina. Euphytica 2011, 179, 143–160. [Google Scholar] [CrossRef]
- Mishra, A.N.; Shirsekar, G.S.; Yadav, S.; Kaushal, K.; Dubey, V.G.; Sai Prasad, S.V. Sources of resistance to Indian pathotypes of Puccinia graminis tritici and P. triticina in durum wheat. Plant Breed. 2015, 134, 508–513. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Chen, W.Q.; Kang, Z.S.; Ma, Z.H.; Xu, S.C.; Jin, S.L.; Jiang, Y.Y. Integrated management of wheat stripe rust caused by Puccinia striiformis f. sp. tritici in China. Sci. Agric. Sin. 2013, 46, 4254–4262. [Google Scholar]
- Roelfs, A.P. Rust Diseases of Wheat: Concepts and Methods of Disease Management; Cimmyt: El Batan, Mexico, 1992. [Google Scholar]
- Sharma, R.K.; Singh, P.K.; Vinod; Joshi, A.K.; Bhardwaj, S.C.; Bains, N.S.; Singh, S. Protecting South Asian Wheat Production from Stem Rust (Ug99) Epidemic. J. Phytopathol. 2013, 161, 299–307. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Hodson, D.P.; Jin, Y.; Lagudah, E.S.; Ayliffe, M.A.; Bhavani, S.; Rouse, M.N.; Pretorius, Z.A.; Szabo, L.J.; Huerta-Espino, J.; et al. Emergence and Spread of New Races of Wheat Stem Rust Fungus: Continued Threat to Food Security and Prospects of Genetic Control. Phytopathology 2015, 105, 872–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S. Deadly new wheat disease threatens Europe’s crops. Nature 2017, 542, 145–146. [Google Scholar] [CrossRef]
- Kolmer, J.A. Genetics of resistance to wheat leaf rust. Annu. Rev. Phytopathol. 1996, 34, 435–455. [Google Scholar] [CrossRef]
- Keller, B.; Krattinger, S.G.; Yahiaoui, N.; Brunner, S.; Kaur, N.; Cloutre, C.; Cloutier, S.; Ling, H.Q.; Travella, S. Molecular analysis of fungal disease resistance in wheat. In Proceedings of the 11th International Wheat Genetics Symposium, Brisbane, QLD, Australia, 24–29 August 2008; Appels, R., Eastwood, R., Lagudah, E., Langridge, P., Mackay, M., Eds.; Sydney University Press: Sydney, NSW, Australia, 2008; Volume 1, Lynne McIntyre and Peter Sharp. [Google Scholar]
- Naik, B.K.; Vinod; Sharma, J.B.; Sivasamy, M.; Prabhu, K.V.; Tomar, R.S.; Tomar, S.M.S. Molecular mapping and validation of the microsatellite markers linked to the Secale cereal derived leaf rust resistance gene Lr45 in wheat. Mol. Breed. 2015, 35, 61. [Google Scholar] [CrossRef]
- Rani, K.; Raghu, B.R.; Jha, S.K.; Agarwal, P.; Mallick, N.; Niranjana, M.; Sharma, J.B.; Singh, A.K.; Sharma, N.K.; Rajkumar, S.; et al. A novel leaf rust resistance gene introgressed from Aegilops markgrafii maps on chromosome arm 2AS of wheat. Theor. Appl. Genet. 2020, 133, 2685–2694. [Google Scholar] [CrossRef] [PubMed]
- Dinkar, V.; Jha, S.K.; Mallick, N.; Niranjana, M.; Agarwal, P.; Sharma, J.B.; Vinod. Molecular mapping of a new recessive wheat leaf rust resistance gene originating from Triticum spelta. Sci. Rep. 2020, 10, 22113. [Google Scholar] [CrossRef] [PubMed]
- Bhawar, K.B.; Vinod; Sharma, J.B.; Singh, A.K.; Sivasamy, M.; Singh, M.; Prabhu, K.V.; Tomar, S.M.S.; Sharma, T.R.; Singh, B. Molecular marker assisted pyramiding of leaf rust resistance genes Lr19 and Lr28 in wheat variety HD2687. Indian J. Genet. 2011, 71, 304–311. [Google Scholar]
- Mallick, N.; Vinod.; Sharma, J.B.; Tomar, R.S.; Sivasamy, M.; Prabhu, K.V. Marker-assisted backcross breeding to combine multiple rust resistance in wheat. Plant Breed. 2015, 134, 172–177. [Google Scholar] [CrossRef]
- Singh, A.K.; Sharma, J.B.; Vinod; Singh, P.K.; Singh, A.; Mallick, N. Genetics and mapping of a new leaf rust resistance gene in Triticum aestivum L. × Triticum timopheevii Zhuk. derivative ‘Selection G12’. J. Genet. 2017, 96, 291–297. [Google Scholar] [CrossRef]
- Mallick, N.; Agarwal, P.; Jha, S.K.; Niranjana, M.; Vinod. Marker-assisted breeding for rust management in wheat. Indian Phytopathol. 2021, 74, 365–370. [Google Scholar] [CrossRef]
- Bansal, M.; Kaur, S.; Dhaliwal, H.S.; Bains, N.S.; Bariana, H.S.; Chhuneja, P.; Bansal, U.K. Mapping of Aegilops umbellulata-derived leaf rust and stripe rust resistance loci in wheat. Plant Pathol. 2017, 66, 38–44. [Google Scholar] [CrossRef]
- Kaur, J. Marker Assisted Pyramiding of Resistance Genes Yr15 and Lr57-Yr40 in High Grain Weight Versions of PBW 550. Master’s Thesis, Punjab Agricultural University, Ludhiana, India, 2018. [Google Scholar]
- Suryavanshi, P.; Singh, Y. Assessment of Unnat PBW 343 (Rust Resistant Wheat Variety) in Mohali, Punjab. Indian J. Pure Appl. Biosci. 2020, 8, 64–69. [Google Scholar] [CrossRef]
- ICAR. Annual Report, (2011–2012); ICAR—Directorate of Wheat Research (Indian Institute of Wheat and Barley Research): Karnal, India, 2012.
- Indian Express. Available online: https://indianexpress.com/article/india/new-blockbuster-wheat-from-iari-6007865/ (accessed on 22 January 2022).
- Mishra, A.N.; Yadav, S.R.; Shirsekar, G.S.; Dubey, V.G.; Kaushal, K.; Sai Prasad, S.V. Diversity for Resistance to Stem and Leaf Rusts in Indian Wheat Germplasm. Indian J. Plant Genet. Resour. 2011, 24, 283–291. [Google Scholar]
- Gireesh, C.; Vinod; Sharma, J.B.; Prabhu, K.V. Inheritance and molecular mapping of leaf rust resistance in Triticum turgidum var. durum cv. Trinakria. Indian J. Genet. 2014, 74, 10–15. [Google Scholar] [CrossRef]
- Agarwal, P.; Jha, S.K.; Sharma, N.K.; Raghunanadan, K.; Mallick, N.; Niranjana, M.; Vinod. Identification of the improved genotypes with 2NS/2AS translocation through molecular markers for imparting resistance to multiple biotic stresses in wheat. Indian J. Genet. 2021, 81, 522–528. [Google Scholar]
- Prasad, P.; Bhardwaj, S.C.; Gangwar, O.P.; Kumar, S.; Khan, H.; Kumar, S.; Rawal, H.C.; Sharma, T.R. Population differentiation of wheat leaf rust fungus Puccinia triticina in South Asia. Curr. Sci. 2017, 112, 2073–2083. [Google Scholar] [CrossRef]
- Rosemary, B.; Kerstin, F.; Mogens, H.; Claude, V.P. Breakdown of the Yr17 resistance to yellow rust of wheat in northern Europe. Agronomie. EDP Sci. 2000, 20, 805–811. [Google Scholar]
- Murray, M.G.; Thompson, W. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berloo, R.V. GGT 2.0: Versatile software for visualization and analysis of genetic data. J. Hered. 2008, 99, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Helguera, M.; Khan, I.A.; Kolmer, J.; Lijavetzky, D.; Zhong-Qi, L.; Dubcovsky, J. PCR assays for the Lr37-Yr17-Sr38 cluster of rust resistance genes and their use to develop isogenic hard red spring wheat lines. Crop Sci. 2003, 43, 1839–1847. [Google Scholar] [CrossRef]
- Stakman, E.C.; Steward, D.M.; Loegering, W.Q. Identification of Physiologic Races of Puccinia graminis var. tritici; USDA: Washington, DC, USA, 1962.
- Peterson, R.F.; Champbell, A.B.; Hannah, A.E. A diagramatic scale for estimating rust intensity of leaves and stem of cereals. Can. J. Res. Sect. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Sheoran, O.P. Online Statistical Analysis (OPSTAT) Software Developed by Chaudhary Charan Singh Haryana Agricultural University, Hisar, India. 2010. Available online: http://www.hau.ernet.in/opstat.html (accessed on 15 December 2021).
 : Plants homozygous for LrTrk.
: Plants homozygous for LrTrk.
: Plants homozygous for LrTrk.
: Plants homozygous for LrTrk.





| Recipient Parent | Target Gene | Generation | No. of Plants Screened with Linked Molecular Marker | No. of Plants Carrying Target Gene | No. of Plants Selected for Background Selection | Number of Plants Backcrossed/Selfed/Selected | Recurrent Parent Genome (RPG) Recovery (%) | |
|---|---|---|---|---|---|---|---|---|
| Heterozygous | Homozygous | |||||||
| HD2967 | LrTrk | BC1F1 | 145 | 60 | - | 10 | 1 | 78.91–83.13 |
| BC2F1 | 66 | 39 | - | 10 | 1 | 90.36–93.37 | ||
| BC2F2 | 200 | 98 | 61 | 61 | 32 | 95.18–98.79 | ||
| No. of plants screened for leaf and stripe rust | 32 |
| No. of plants resistant to leaf rust only | 16 |
| No. of plants resistant to both leaf and stripe rust | 14 |
| No. of plants selected for replicated trials | 6 |
| (RPG) recovery (%) of selected plants in replicated trial | 97.59–98.79 |
| S. No. | Near Isogenic Lines of HD2967 | ITs for Leaf Rust Race 77-5 | Response to Stripe Rust Race 110S119 |
|---|---|---|---|
| 1 | HD2967 + LrTrk/YrTrk-137-21-82 | ;1 | 10R |
| 2 | HD2967 + LrTrk/YrTrk-137-21-19 | ;1 | 10R |
| 3 | HD2967 + LrTrk-137-21-28 | ;1 | 40S |
| 4 | HD2967 + LrTrk-137-21-16 | ;1 | 30S |
| 5 | HD2967 + LrTrk-137-21-161 | ;1 | 40S |
| 6 | HD2967 + LrTrk-137-21-163 | ;1 | 40S |
| 7 | HD2967 | 3 | 60S |
| 8 | Trinakria | ; | 10R |
| NILs | PH | SL | NSpl | NS | TKW (gm) | YLD (kg) |
|---|---|---|---|---|---|---|
| HD2967 + LrTrk/YrTrk-137-21-82 | 111.40 * | 13.72 * | 25.80 * | 75.60 | 37.25 | 3.94 |
| HD2967 + LrTrk/YrTrk-137-21-19 | 104.40 | 12.62 | 23.40 | 73.20 | 37.75 | 3.69 |
| HD2967 + LrTrk-137-21-28 | 100.20 | 11.04 | 22.20 | 71.40 | 42.00 * | 4.02 |
| HD2967 + LrTrk-137-21-16 | 103.20 | 10.52 * | 21.80 * | 68.20 | 37.00 | 3.42 |
| HD2967 + LrTrk-137-21-161 | 99.80 | 11.68 | 22.60 | 71.80 | 42.00 * | 4.11 |
| HD2967 + LrTrk -137-21-163 | 101.40 | 13.18 * | 24.20 | 75.20 | 37.00 | 3.71 |
| HD2967 | 101.20 | 11.78 | 23.40 | 72.00 | 36.50 | 3.63 |
| Mean | 103.08 | 12.07 | 23.34 | 72.48 | 38.5 | 3.78 |
| SD | 4.46 | 1.34 | 1.54 | 5.59 | 2.52 | 0.28 |
| CD | 3.58 | 1.18 | 1.31 | 7.37 | 3.8 | 0.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mallick, N.; Jha, S.K.; Agarwal, P.; Mall, A.; M., N.; Kumar, S.; Choudhary, M.K.; Bansal, S.; Saharan, M.S.; Sharma, J.B.; et al. Marker-Assisted Improvement of Bread Wheat Variety HD2967 for Leaf and Stripe Rust Resistance. Plants 2022, 11, 1152. https://doi.org/10.3390/plants11091152
Mallick N, Jha SK, Agarwal P, Mall A, M. N, Kumar S, Choudhary MK, Bansal S, Saharan MS, Sharma JB, et al. Marker-Assisted Improvement of Bread Wheat Variety HD2967 for Leaf and Stripe Rust Resistance. Plants. 2022; 11(9):1152. https://doi.org/10.3390/plants11091152
Chicago/Turabian StyleMallick, Niharika, Shailendra K. Jha, Priyanka Agarwal, Anchal Mall, Niranjana M., Sachin Kumar, Manish K. Choudhary, Shreshtha Bansal, M. S. Saharan, J. B. Sharma, and et al. 2022. "Marker-Assisted Improvement of Bread Wheat Variety HD2967 for Leaf and Stripe Rust Resistance" Plants 11, no. 9: 1152. https://doi.org/10.3390/plants11091152