
Nutrient Concentrations Induced Abiotic Stresses to Sweet Pepper Seedlings in Hydroponic Culture

 ,
,  ,
,  and
and
Abstract
:1. Introduction
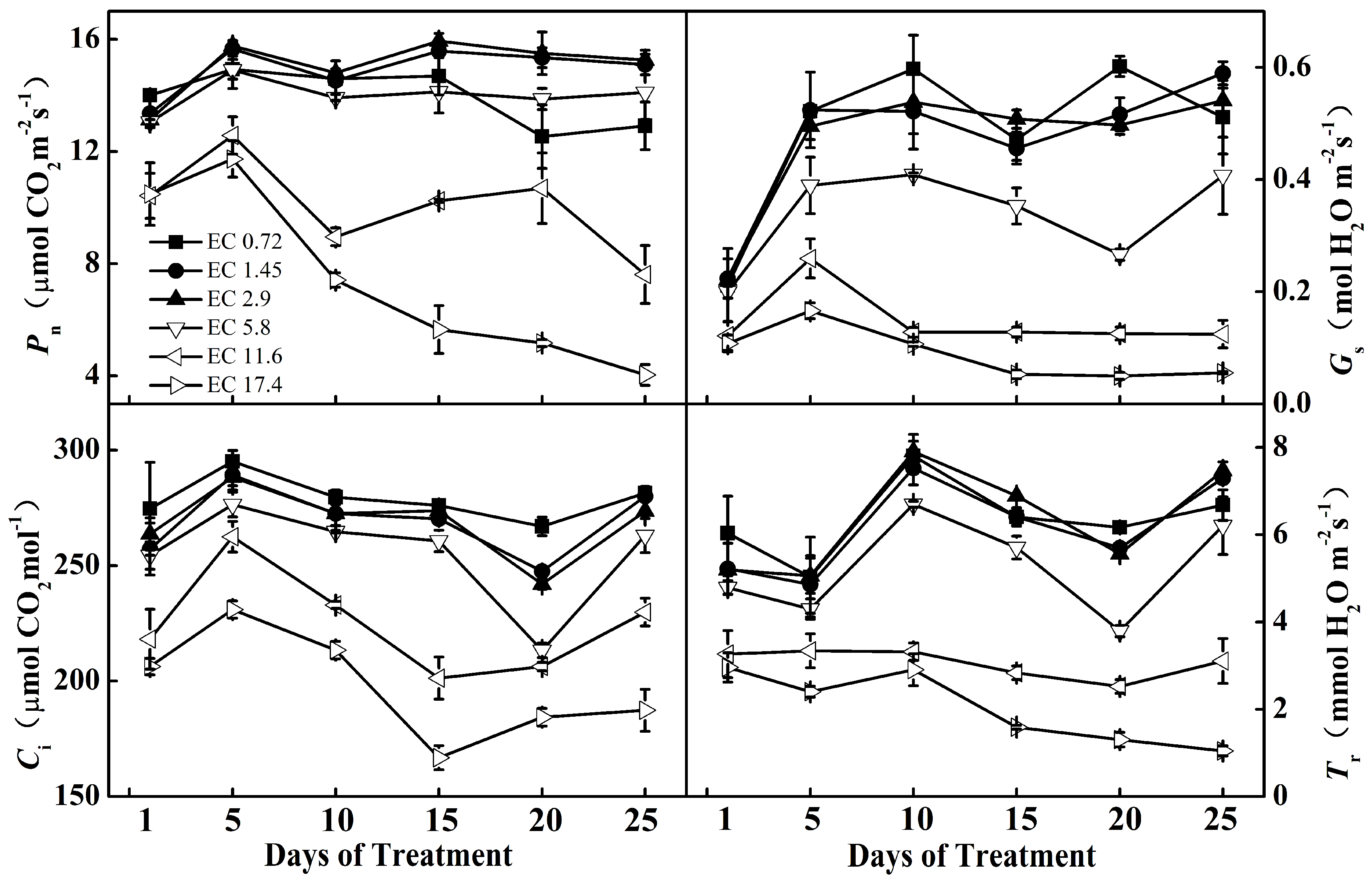
2. Results
2.1. Plant Growth Analysis
2.2. Leaf Gas Exchange Analysis
2.3. Analyses of the Contents of Leaf Chlorophyll (Chl) and Carotenoids (Car)
2.4. Root Activity Analysis
2.5. AE Analysis
2.6. Analyses of the Contents of Soluble Protein, MDA, and Proline
2.7. PME Content Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. NS Treatments
4.3. Measurements of Sweet Pepper Plant Height, Stem Diameter, Shoot Fresh Weight (FW), Dry Weight (DW), and Shoot Moisture Content (MC)
4.4. Measurements of Leaf Gas Exchange Parameters
4.5. Measurements of Leaf Chlorophyll Content, Carotenoid Content, and Root Activity
4.6. AE Activity Assays
4.7. Measurements of Proline, Soluble Protein, and MDA Contents
4.8. PME Determination
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Pn | Gs | Ci | Tr | ||||
|---|---|---|---|---|---|---|---|---|
| F Value | p Value | F Value | p Value | F Value | p Value | F Value | p Value | |
| EC treatment | 148.84 | <0.001 | 141.59 | <0.001 | 128.90 | <0.001 | 119.93 | <0.001 |
| Date | 15.34 | <0.001 | 30.15 | <0.001 | 34.63 | <0.001 | 16.45 | <0.001 |
| EC treatment× Date | 5.75 | <0.001 | 4.26 | <0.001 | 2.54 | <0.001 | 3.05 | <0.001 |
References
- Savvas, D.; Gianquinto, G.; Tuzel, Y.; Gruda, N. Soilless culture. In Good Agricultural Practices for Greenhouse Vegetable Crops. Principles for Mediterranean Climate Areas; Baudoin, W., Ed.; Food and Agricultural Organization of the United Nations: Rome, Italy, 2013; pp. 303–354. [Google Scholar]
- Putra, P.A.; Yuliando, H. Soilless culture system to support water use efficiency and product quality: A review. Agric. Agric. Sci. Procedia 2015, 3, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Othman, Y.; Bataineh, K.; Al-Ajlouni, M.; Alsmairat, N.; Hilaire, R.S. Soilless culture: Management of growing substrate, water, nutrient, salinity, microorganism and product quality. Fresen Environ. Bull. 2019, 28, 3249–3260. [Google Scholar]
- Marques, G.; Aleixo, D.; Pitarma, R. Enhanced hydroponic agriculture environmental monitoring: An internet of things approach. In Computational Science—ICCS 2019, Proceedings of the 19th International Conference, Faro, Portugal, 12–14 June 2019; Lecture Notes in Computer Science; Rodrigues, J.M.F., Cardoso, P.J.S., Monteiro, J., Lam, R., Krzhizhanovskaya, V.V., Michael, H., Lees, M.H., Dongarra, J.J., Sloot, P.M.A., Eds.; Springer: Cham, Switzerland, 2019; Volume 11538, pp. 658–669. [Google Scholar] [CrossRef]
- Cho, W.J.; Kim, H.J.; Jung, D.H.; Kim, D.W.; Ahn, T.I.; Son, J.E. On-site ion monitoring system for precision hydroponic nutrient management. Comput. Electron. Agric. 2018, 146, 51–58. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, W.M.; Martínez, V.; Nieves, M.; Simón, I.; Lidón, V.; Fernandez-Zapata, J.; Martinez-Nicolas, J.; Cámara-Zapata, J.M.; García-Sánchez, F. Agricultural and physiological responses of tomato plants grown in different soilless culture systems with saline water under greenhouse conditions. Sci. Rep. 2019, 9, 6733. [Google Scholar] [CrossRef]
- Atzori, G.; Mancuso, S.; Masi, E. Seawater potential use in soilless culture: A review. Sci. Hortic. 2019, 249, 199–207. [Google Scholar] [CrossRef]
- Ding, X.; Jiang, Y.; Zhao, H.; Guo, D.; He, L.; Liu, F.; Zhou, Q.; Nandwani, D.; Hui, D.; Yu, J. Electrical conductivity of nutrient solution influenced photosynthesis, quality, and antioxidant enzyme activity of pakchoi (Brassica campestris L. ssp. Chinensis) in a hydroponic system. PLoS ONE 2018, 13, e0202090. [Google Scholar] [CrossRef]
- Signore, A.; Serio, F.; Santamaria, P. A targeted management of the nutrient solution in a soilless tomato crop according to plant needs. Front. Plant Sci. 2016, 7, 391. [Google Scholar] [CrossRef] [Green Version]
- Fratianni, F.; d’Acierno, A.; Cozzolino, A.; Spigno, P.; Riccardi, R.; Raimo, F.; Pane, C.; Zaccardelli, M.; Tranchida Lombardo, V.; Tucci, M. Biochemical characterization of traditional varieties of sweet pepper (Capsicum Annuum L.) of the Campania Region, Southern Italy. Antioxidants 2020, 9, 556. [Google Scholar] [CrossRef]
- Singh, H.; Dunn, B.L.; Payton, M.; Brandenberger, L. Selection of fertilizer and cultivar of sweet pepper and eggplant for hydroponic production. Agronomy 2019, 9, 433. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Stoleru, V.V.; Munteanu, N.C.; Sellitto, V.M.; Teliban, G.C.; Burducea, M.; Tenu, I.; Morano, G.; Butnariu, M. Quality performances of sweet pepper under farming management. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 458–464. [Google Scholar] [CrossRef] [Green Version]
- Raffo, A.; Baiamonte, I.; Nardo, N.; Paoletti, F. Internal quality and antioxidants content of cold-stored red sweet peppers as affected by polyethylene bag packaging and hot water treatment. Eur. Food Res. Technol. 2007, 225, 395–405. [Google Scholar] [CrossRef]
- Mamphogoro, T.P.; Babalola, O.O.; Aiyegoro, O.A. Sustainable management strategies for bacterial wilt of sweet peppers (Capsicum annuum) and other Solanaceous crops. J. Appl. Microbiol. 2020, 129, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Khaitov, B.; Umurzokov, M.; Cho, K.M.; Lee, Y.J.; Park, K.W.; Sung, J. Importance and production of chilli pepper; heat tolerance and efficient nutrient use under climate change conditions. Korean J. Agric. Sci. 2019, 46, 769–779. [Google Scholar] [CrossRef]
- Abdel-Mawgoud, A.M.R.; El-Abd, S.O.; Stanghellini, C.; Böhme, M.; Abou-Hadid, A.F. Sweet pepper crop responses to greenhouse climate manipulation under saline conditions. Acta Hortic. 2004, 659, 431–438. [Google Scholar] [CrossRef]
- Rubio, J.S.; Pereira, W.E.; Garcia-Sanchez, F.; Murillo, L.; García, A.L.; Martínez, V. Sweet pepper production in substrate in response to salinity, nutrient solution management and training system. Hortic. Bras. 2011, 29, 275–281. [Google Scholar] [CrossRef]
- Amalfitano, C.A.; Del Vacchio, L.D.V.; Somma, S.; Cuciniello, A.C.; Caruso, G. Effects of cultural cycle and nutrient solution electrical conductivity on plant growth, yield and fruit quality of ‘Friariello’ pepper grown in hydroponics. Hortic. Sci. 2017, 44, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Lu, N.; Bernardo, E.L.; Tippayadarapanich, C.; Takagaki, M.; Kagawa, N.; Yamori, W. Growth and accumulation of secondary metabolites in perilla as affected by photosynthetic photon flux density and electrical conductivity of the nutrient solution. Front. Plant Sci. 2017, 8, 708. [Google Scholar] [CrossRef] [Green Version]
- Lam, V.P.; Kim, S.J.; Park, J.S. Optimizing the electrical conductivity of a nutrient solution for plant growth and bioactive compounds of Agastache rugosa in a plant factory. Agronomy 2020, 10, 76. [Google Scholar] [CrossRef] [Green Version]
- Albornoz, F.; Lieth, J.H. Over fertilization limits lettuce productivity because of osmotic stress. Chil. J. Agric. Res. 2015, 75, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Jiang, Y.; Hao, T.; Jin, H.; Zhang, H.; He, L.; Zhou, Q.; Huang, D.; Hui, D.; Yu, J. Effects of heat shock on photosynthetic properties, antioxidant enzyme activity, and downy mildew of cucumber (Cucumis sativus L.). PLoS ONE 2016, 11, e0152429. [Google Scholar] [CrossRef]
- Yan, K.; Chen, P.; Shao, H.; Zhao, S.; Zhang, L.; Zhang, G.; Sun, X. Responses of photosynthesis and photosystem II to higher temperature and salt stress in Sorghum. J. Agron. Crop Sci. 2012, 198, 218–225. [Google Scholar] [CrossRef]
- Albornoz, F.; Lieth, J.H.; González-Fuentes, J.A. Effect of different day and night nutrient solution concentrations on growth, photosynthesis, and leaf NO3-content of aeroponically grown lettuce. Chil. J. Agric. Res. 2014, 74, 240–245. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Ding, X.; Zhang, D.; Deng, Q.; Yu, C.; Zhou, S.; Hui, D. Soil salinity increases the tolerance of excessive sulfur fumigation stress in tomato plants. Environ. Exp. Bot. 2017, 133, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, A.; Tanaka, R. Chlorophyll metabolism. Curr. Opin. Plant Biol. 2006, 9, 248–255. [Google Scholar] [CrossRef]
- Havaux, M. Carotenoids as membrane stabilizers in chloroplasts. Trends Plant Sci. 1998, 3, 147–151. [Google Scholar] [CrossRef]
- Stefanov, M.; Yotsova, E.; Rashkov, G.; Ivanova, K.; Markovska, Y.; Apostolova, E.L. Effects of salinity on the photosynthetic apparatus of two Paulownia lines. Plant Physiol. Biochem. 2016, 101, 54–59. [Google Scholar] [CrossRef]
- Xu, C.; Mou, B. Responses of spinach to salinity and nutrient deficiency in growth, physiology, and nutritional value. J. Am. Soc. Hortic. Sci. 2016, 141, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, H.; Yu, F.; Hu, B.; Jia, Y.; Sha, H.; Zhao, H. Differential activity of the antioxidant defence system and alterations in the accumulation of osmolyte and reactive oxygen species under drought stress and recovery in rice (Oryza sativa L.) tillering. Sci. Rep. 2019, 9, 8543. [Google Scholar] [CrossRef]
- Arough, Y.K.; Sharifi, R.S.; Sedghi, M.; Barmaki, M. Effect of zinc and bio fertilizers on antioxidant enzymes activity, chlorophyll content, soluble sugars and proline in triticale under salinity condition. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.W.; Stals, E.; Panis, B.; Keulemans, J.; Swennen, R.L. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem. 2005, 347, 201–207. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Kusvuran, S.; Kiran, S.; Ellialtioglu, S.S. Antioxidant enzyme activities and abiotic stress tolerance relationship in vegetable crops. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives, 1st ed.; InTech: Rijeka, Croatia, 2016; pp. 481–503. [Google Scholar]
- Talhouni, M.; Sönmez, K.; Kiran, S.; Beyaz, R.; Yildiz, M.; Kuşvuran, Ş.; Ellialtıoğlu, Ş.Ş. Comparison of salinity effects on grafted and non-grafted eggplants in terms of ion accumulation, MDA content and antioxidative enyzme activities. Adv. Hortic. Sci. 2019, 33, 87–95. [Google Scholar]
- Ahmadi, M.; Souri, M.K. Nutrient uptake, proline content and antioxidant enzymes activity of pepper (Capsicum Annuum L.) under higher electrical conductivity of nutrient solution created by nitrate or chloride salts of potassium and calcium. Acta Sci. Pol. Hortorum Cultus 2019, 18, 113–122. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Soares, C.; Carvalho, M.E.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Roza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Uarrota, V.G.; Moresco, R.; Schmidt, E.C.; Bouzon, Z.L.; Nunes, E.C.; Neubert, E.O.; Peruch, L.A.M.; Rocha, M.; Maraschin, M. The role of ascorbate peroxidase, guaiacol peroxidase, and polysaccharides in cassava (Manihot esculenta Crantz) roots under postharvest physiological deterioration. Food Chem. 2016, 197, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol. Rep. 2011, 5, 353–365. [Google Scholar] [CrossRef]
- Yang, Z.; Li, Y.; Li, P.; Zhang, F.; Thomas, B. Effect of difference between day and night temperature on tomato (Lycopersicon esculentum Mill.) root activity and low molecular weight organic acid secretion. Soil Sci. Plant Nutr. 2016, 62, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J. Subsoil root activity in tree-based cropping systems. In Roots: The Dynamic Interface between Plants and the Earth; Springer: Dordrecht, The Netherlands, 2003; pp. 319–331. [Google Scholar]
- Dresbøll, D.B.; Thorup-Kristensen, K. Spatial variation in root system activity of tomato (Solanum lycopersicum L.) in response to short and long-term waterlogging as determined by 15 N uptake. Plant Soil 2012, 357, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Gai, Z.; Zhang, J.; Li, C. Effects of starter nitrogen fertilizer on soybean root activity, leaf photosynthesis and grain yield. PLoS ONE 2017, 12, e0174841. [Google Scholar] [CrossRef] [Green Version]
- Cseresnyés, I.; Rajkai, K.; Szitár, K.; Radimszky, L.; Onodi, G.; Kroel-Dulay, G. Root capacitance measurements allow non-intrusive in-situ monitoring of the seasonal dynamics and drought response of root activity in two grassland species. Plant Soil 2020, 449, 423–437. [Google Scholar] [CrossRef] [Green Version]
- Xuan, L.; Hussain, N.; Wang, Z.; Jiang, Y.; Chen, M.; Jiang, L. Comparison of vitality between seedlings germinated from black-coated and yellow-coated seeds of a turnip rape (Brassica rapa L.) subjected to NaCl and CdCl2 stresses. Plant Growth Regul. 2015, 76, 61–70. [Google Scholar] [CrossRef]
- Belouchrani, A.S.; Bouderbala, A.; Drouiche, N.; Lounici, H. The interaction effect to fertilization on the mineral nutrition of canola under different salinity levels. J. Plant Growth Regul. 2021, 40, 848–854. [Google Scholar] [CrossRef]
- Erdinc, C. Changes in ion (K, Ca and Na) regulation, antioxidant enzyme activity and photosynthetic pigment content in melon genotypes subjected to salt stress—A mixture modeling analysis. Acta Sci. Pol. Hortorum Cultus 2018, 17, 165–183. [Google Scholar] [CrossRef]
- Qiu, R.; Yang, Z.; Jing, Y.S.; Liu, C.W.; Luo, X.S.; Wang, Z.C. Effects of irrigation water salinity on the growth, gas exchange parameters, and ion concentration of hot pepper plants modified by leaching fractions. HortScience 2018, 53, 1050–1055. [Google Scholar] [CrossRef] [Green Version]
- Komosa, A.; Górniak, T. The effect of chloride on yield and nutrient interaction in greenhouse tomato (Lycopersicon Esculentum Mill.) grown in rockwool. J. Plant Nutr. 2015, 38, 355–370. [Google Scholar] [CrossRef]
- Souri, M.K.; Hatamian, M. Aminochelates in plant nutrition: A review. J. Plant. Nutr. 2019, 42, 67–78. [Google Scholar] [CrossRef]
- Stanghellini, C.; Oosfer, B.; Heuvelink, E. Greenhouse Horticulture: Technology for Optimal Crop Production; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019. [Google Scholar]
- Clemensson-Lindell, A.; Persson, H. Fine-root vitality in a Norway spruce stand subjected to various nutrient supplies. Plant Soil 1995, 168, 167–172. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, G.; Bian, X.; Zhao, Q. Effects of root interaction and nitrogen fertilization on the chlorophyll content, root activity, photosynthetic characteristics of intercropped soybean and microbial quantity in the rhizosphere. Plant Soil Environ. 2013, 59, 80–88. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Durner, J.; Klessig, D.F. Salicylic acid is a modulator of tobacco and mammalian catalases. J. Biol. Chem. 1996, 271, 28492–28501. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1980, 22, 867–880. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Koca, H.; Bor, M.; Özdemir, F.; Türkan, İ. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ. Exp. Bot. 2007, 60, 344–351. [Google Scholar] [CrossRef]
- Nostar, O.; Ozdemir, F.; Bor, M.; Turkan, I.; Tosun, N. Combined effects of salt stress and cucurbit downy mildew (Pseudoperospora cubensis Berk. and Curt. Rostov.) infection on growth, physiological traits and antioxidant activity in cucumber (Cucumis sativus L.) seedlings. Physiol. Mol. Plant Pathol. 2013, 83, 84–92. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Song, J.; Huang, H.; Hao, Y.; Song, S.; Zhang, Y.; Su, W.; Liu, H. Nutritional quality, mineral and antioxidant content in lettuce affected by interaction of light intensity and nutrient solution concentration. Sci. Rep. 2020, 10, 2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]







| Treatment | Plant Height (cm) | Stem Diameter (cm) | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Shoot Moisture Content (%) |
|---|---|---|---|---|---|
| EC0.72 | 57.0 ± 1.58 b | 0.85 ± 0.050 b | 94.4 ± 6.89 c | 13.43 ± 0.94 b | 85.77 ± 0.14 c |
| EC1.45 | 60.6 ± 1.95 a | 0.89 ± 0.037 a | 114.3 ± 7.52 ab | 15.14 ± 0.80 ab | 86.74 ± 0.18 b |
| EC2.9 | 62.4 ± 1.14 a | 0.90 ± 0.060 a | 125.3 ± 7.54 a | 16.09 ± 2.17 a | 87.17 ± 0.26 a |
| EC5.8 | 56.4 ± 1.14 b | 0.84 ± 0.032 b | 107.5 ± 1.36 b | 15.36 ± 0.05 a | 85.71 ± 0.19 c |
| EC11.6 | 45.0 ± 2.35 c | 0.79 ± 0.019 c | 64.0 ± 4.28 d | 10.55 ± 0.73 c | 83.50 ± 0.22 d |
| EC17.4 | 39.2 ± 1.30 d | 0.71 ± 0.036 d | 50.8 ± 5.49 e | 9.17 ± 0.97 c | 81.93 ± 0.31 e |
| Treatment | Soluble Protein Content (mg g−1 FW) | MDA Content (mg g−1 FW) | Proline Content (μg g−1 FW) |
|---|---|---|---|
| EC0.72 | 2.03 ± 0.14 e | 1.35 ± 0.11 d | 3.70 ± 0.19 d |
| EC1.45 | 3.11 ± 0.07 d | 1.58 ± 0.03 c | 5.11 ± 0.67 d |
| EC2.9 | 3.44 ± 0.08 c | 1.24 ± 0.07 d | 9.83 ± 0.53 d |
| EC5.8 | 3.62 ± 0.05 b | 1.61 ± 0.07 c | 23.87 ± 2.18 c |
| EC11.6 | 3.97 ± 0.10 a | 1.85 ± 0.11 b | 111.05 ± 1.57 b |
| EC17.4 | 3.68 ± 0.07 b | 2.06 ± 0.07 a | 153.40 ± 7.82 a |
| A | kg/1000 L | A | g/1000 L | B | kg/1000 L |
|---|---|---|---|---|---|
| Ca(NO3)2·4H2O | 130 | MnSO4·H2O | 175 | KNO3 | 50 |
| EDPA-Fe (13%Fe) | 1.4 | ZnSO4·7H2O | 125 | KH2PO4 | 25 |
| Na2B4O7·4H2O | 375 | K2SO4 | 15 | ||
| CuSO4·5H2O | 25 | MgSO4·7H2O | 50 | ||
| Na2MoO4·2H2O | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Zhang, H.; Qian, T.; He, L.; Jin, H.; Zhou, Q.; Yu, J. Nutrient Concentrations Induced Abiotic Stresses to Sweet Pepper Seedlings in Hydroponic Culture. Plants 2022, 11, 1098. https://doi.org/10.3390/plants11081098
Ding X, Zhang H, Qian T, He L, Jin H, Zhou Q, Yu J. Nutrient Concentrations Induced Abiotic Stresses to Sweet Pepper Seedlings in Hydroponic Culture. Plants. 2022; 11(8):1098. https://doi.org/10.3390/plants11081098
Chicago/Turabian StyleDing, Xiaotao, Hongmei Zhang, Tingting Qian, Lizhong He, Haijun Jin, Qiang Zhou, and Jizhu Yu. 2022. "Nutrient Concentrations Induced Abiotic Stresses to Sweet Pepper Seedlings in Hydroponic Culture" Plants 11, no. 8: 1098. https://doi.org/10.3390/plants11081098