
Low Concentration of Anti-Auxin and Anti-Fungal Agent Accelerates the PLB Regeneration of Dendrobium okinawense under Green LED



Abstract
:1. Introduction
2. Results
2.1. Evaluation of the Sucrose Concentrations
2.2. LEDs Lights for PLB Regeneration
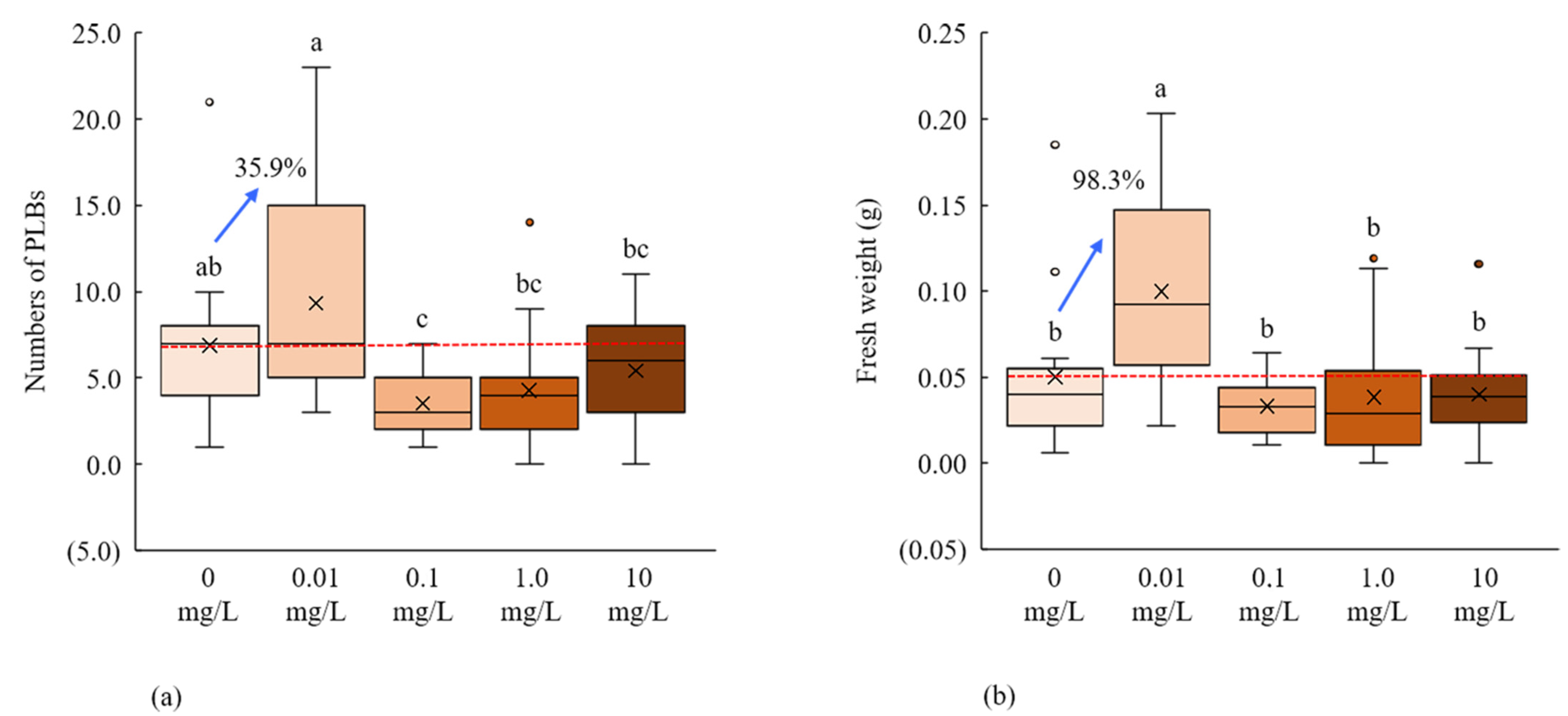
2.3. Anti-Auxin, PCIB, for PLB Regeneration
2.4. An Antifungal Agrochemical, HMI for PLB Regeneration of D. okinawense
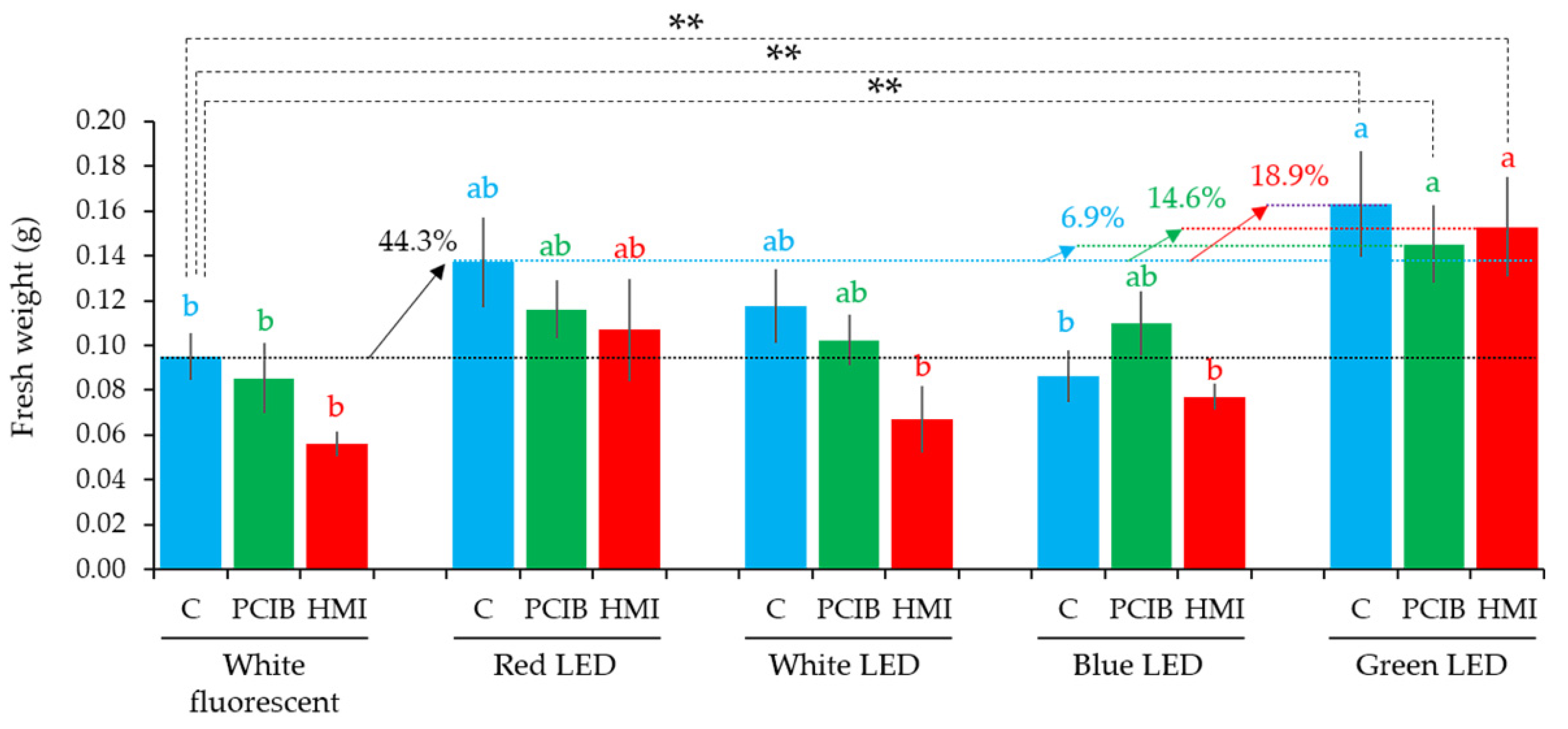
2.5. Effect of LEDs on PCIB- and HMI-Manipulated Culture Media for the PLB Regeneration
3. Discussion
4. Materials and Methods
4.1. Modification of MS Media, pH Adjustment, and Solidification
4.2. Preparation of Explants
4.3. Volume of Culture Media, Number of Explants in Each Culture Vessel, and Photoperiod
4.4. Evaluation of the Sucrose Level for PLB Regeneration
4.5. Manipulation of the Environment and Culture Media
4.5.1. Manipulation of the Environment by Light-Emitting Diodes
4.5.2. Manipulation of the Culture Media by Anti-Auxin and Antifungal Agent
4.5.3. PCIB and HMI Manipulated Culture Media under LEDs
4.6. Data Collection and Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arditti, J.; Ghani, A.K.A. Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, E.C. A perspective on orchid seed and protocorm development. Bot. Stud. 2017, 58, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Huh, Y.S.; Paek, K.Y. Common protocols in orchid micropropagation. In Orchid Propagation: From Laboratories to Greenhouses—Methods and Protocols, Springer Protocols Handbooks, 1st ed.; Lee, Y.I., Yeung, E.C.-T., Eds.; Humana Press: New York, NY, USA, 2018; pp. 1–524. [Google Scholar]
- Roberto, S.R.; Colombo, R.C. Innovation in propagation of fruit, vegetable and ornamental plants. Horticulturae 2020, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, J.C.; Zanello, C.A.; Chen, J.-T. An overview of orchid protocorm-like bodies: Mass propagation, biotechnology, molecular aspects, and breeding. Int. J. Mol. Sci. 2020, 21, 985. [Google Scholar] [CrossRef] [Green Version]
- Chugh, S.; Guha, S.; Rao, U. Micropropagation of orchids: A review on the potential of different explants. Sci. Hortic. 2009, 122, 507–520. [Google Scholar] [CrossRef]
- Cui, H.-Y.; Murthy, H.N.; Moh, S.H.; Cui, Y.; Lee, E.-J.; Paek, K.-Y. Protocorm culture of Dendrobium candidum in balloon type bubble bioreactors. Biochem. Eng. J. 2014, 88, 26–29. [Google Scholar] [CrossRef]
- Habiba, S.U.; Shimasaki, K.; Ahsan, M.M.; Alam, M.M. Effects of different light quality on growth and development of protocorm like bodies (PLBs) in Dendrobium kingianum cultured in vitro. Bangladesh Res. Pub. J. 2014, 10, 223–227. [Google Scholar]
- Habiba, S.U.; Shimasaki, K.; Ahsan, M.M.; Alam, M.M. Effect of 6-benzylaminopurine (BA) and hyaluronic acid (HA) under white light emitting diode (LED) on regeneration in protocorm-like bodies (PLBs) of Dendrobium kingianum. Am. Eurasian J. Agric. Environ. Sci. 2014, 14, 605–609. [Google Scholar]
- Parthibhan, S.; Rao, M.V.; Kumar, T.S. In vitro regeneration from protocorm in Dendrobium aqueum Lindley—An imperiled orchid. J. Genet. Eng. Biotechnol. 2015, 13, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.; Lu, S. Dendrobium okinawense Hatusima & Ida (Orchidaceae): A newly recorded species in Taiwan. Taiwania 2007, 52, 106–112. [Google Scholar]
- Hatsushima, S.; Ida, R. A new Dendrobium from Isl. Okinawa. J. Geobot. 1970, 18, 77–79. [Google Scholar]
- Kim, S.-K.; Chang, S.C.; Lee, E.J.; Chung, W.-S.; Kim, Y.-S.; Hwang, S.; Lee, J.S. Involvement of brassinosteroids in the gravitropic response of primary root of maize. Plant Physiol. 2000, 123, 997–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Frugis, G.; Colgan, D.; Chua, N.H. Arabidopsis NAC1 transduced auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 2000, 14, 3024–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burström, H. Studies on growth and metabolism of roots. IV. Positive and negative auxin effects on cell elongation. Physiol. Plant 1950, 3, 277–292. [Google Scholar] [CrossRef]
- MacRae, D.H.; Bonner, J. Chemical structure and antiauxin activity. Physiol. Plant 1953, 6, 485–510. [Google Scholar] [CrossRef]
- Ma, Q.; Grones, P.; Robert, S. Auxin signaling: A big question to be addressed by small molecules. J. Exp. Bot. 2018, 69, 313–328. [Google Scholar] [CrossRef] [Green Version]
- Novak, S.D.; Luna, L.J.; Gamage, R.N. Role of auxin in orchid development. Plant Signal Behav. 2014, 9, e2972277. [Google Scholar] [CrossRef] [Green Version]
- Ota, Y. Plant growth-promoting activities of 3-hydroxy-5-methyl-isoxazole. Jpn. Agric. Res. Q. 1975, 9, 1–7. [Google Scholar]
- Harveson, R.M.; Windels, C.E.; Smith, J.A.; Brantner, J.R.; Cattnach, A.W.; Giles, J.F.; Hubbell, L.; Cattnach, N.R. Fungicide registration and a small niche market: A case history of hymexazol seed treatment and the U.S. sugar beet industry. Plant Dis. 2007, 91, 780–790. [Google Scholar]
- Yan, L.; Yukun, Q.; Song, L.; Pengcheng, L.; Rong’e, X. Preparation, characterization, and antifungal activity of hymexazol-linked chitosan derivatives. Chin. J. Oceanol. Limnol. 2017, 35, 1079–1085. [Google Scholar]
- Amaki, W.; Yamazaki, N.; Ichimura, M.; Watanabe, H. Effects of light quality on the growth and essential oil content in sweet basil. Acta Hortic. 2011, 907, 91–94. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Virsilė, A.; Samuolienė, G.; Jankauskienė, J.; Sakalauskienė, S.; Sirtautas, R.; Novičkovas, A.; Dabasinskas, L.; Vastakaite, V.; Miliauskienė, J.; et al. Light quality: Growth and nutritional value of microgreens under indoor and greenhouse conditions. Acta Hortic. 2016, 1134, 277–284. [Google Scholar] [CrossRef]
- Bugbee, B. Toward an optimal spectral quality for plant growth and development: The importance of radiation capture. Acta Hortic. 2016, 1134, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.T.; Bertolucci, S.K.V.; da Cunha, S.H.B.; Lazzarini, L.E.S.; Tavares, M.C.; Pinto, J.E.B.P. Effect of light and natural ventilation systems on the growth parameters and carvacrol content in the in vitro cultures of Plectranthus amboinicus (Lour.) Spreng. Plant Cell Tiss. Org. 2017, 129, 501–510. [Google Scholar] [CrossRef]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z.; Liu, X.; Han, X. Effects of different light sources on the growth of non-heading Chinese cabbage (Brassica campestris L.). J. Agric. Sci. 2012, 4, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue light added with red LEDs enhance growth characteristics, pigments content, and antioxidant capacity in lettuce, spinach, kale, basil, and sweet pepper in a controlled environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.; Yoshihara, T. Effect of green light wavelength and intensity on photomorphogenesis and photosynthesis in Lactuca sativa. Environ. Expt. Bot. 2012, 75, 128–133. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Sirtautas, R.; Novičkovas, A.; Duchovskis, P. The effect of supplementary LED lighting on the antioxidant and nutritional properties of lettuce. Acta Hortic. 2012, 952, 835–841. [Google Scholar] [CrossRef]
- Samuolienė, G.; Sirtautas, R.; Brazaitytė, A.; Duchovskis, P. LED lighting and seasonality effects antioxidant properties of baby leaf lettuce. Food Chem. 2012, 134, 1494–1499. [Google Scholar] [CrossRef]
- Stutte, G.W.; Edney, S.; Skerritt, T. Photoregulation of bioprotectant content of red leaf lettuce with light-emitting diodes. HortScience 2009, 44, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Expt. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Kaewjampa, N.; Shimasaki, K. Effects of green LED lighting on organogenesis and superoxide dismutase (SOD) activities in protocorm-like bodies (PLBs) of Cymbidium cultured in vitro. Environ. Control. Biol. 2012, 50, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Nahar, S.J.; Shimasaki, K.; Haque, S.M. Effect of different light and two polysaccharides on the proliferation of protocrom-like bodies of Cymbidium cultured in vitro. Acta Hortic. 2012, 956, 307–313. [Google Scholar] [CrossRef]
- Nahar, S.J.; Shimasaki, K.; Haque, S.M. Chondroitin sulfate can be a new plant growth regulator for Cymbidium micropropagation. Acta Hortic. 2013, 1014, 449–455. [Google Scholar] [CrossRef]
- Nahar, S.J.; Haque, S.M.; Shimasaki, K. Organogenesis of Cymbidium finlaysonianum under different sources of lights. Am.-Eurasian J. Agric. Environ. Sci. 2015, 15, 2095–2101. [Google Scholar]
- Kamal, M.M.; Shimasaki, K.; Akhter, N. Effect of light emutting diode (LED) lamps and N-Acetyleglucoseamine (NAG) on organogenesis in protocorm-like-bodies (PLBs) of Cymbidium hybrid cultured In Vitro. Plant Tissue Cult. Biotechnol. 2014, 24, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Sultana, K.S.; Hasan, K.M.; Hasan, K.M.; Sultana, S.; Mehraj, H.; Shimasaki, K.; Habiba, S.U. Effect of hyaluronic acid (HA) on organogenesis in protocorm-like-bodies (PLBs) of Phalaenopsis ‘Fmk02010′ cultured in vitro. Am. Eurasian J. Agric. Environ. Sci. 2015, 15, 1721–1724. [Google Scholar]
- Sultana, K.S.; Hasan, K.M.; Hasan, K.M.; Sultana, S.; Mehraj, H.; Shimasaki, K.; Habiba, S.U. Effect of two elicitors on organogenesis in protocorm-like bodies (PLBs) of Phalaenopsis ‘Fmk02010′ cultured in vitro. World Appl. Sci. J. 2015, 33, 1528–1532. [Google Scholar]
- Mehraj, H.; Alam, M.M.; Habiba, S.U.; Mehbub, H. LEDs combined with CHO sources and CCC priming PLB regeneration of Phalaenopsis. Horticulturae 2019, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.D.; Jatothu, B. Fundamentals and applications of light-emitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A. The response of protocorm-like bodies of nine hybrid Cymbidium cultivars to light-emitting diodes. Environ. Exp. Biol. 2014, 12, 155–159. [Google Scholar]
- Okinawa Prefecture. Threatened Wildlife in Okinawa, 2nd ed.; Fungi and Plants; Red Data Okinawa; Nature Conservation Division, Department of Cultural & Environmental Affairs, Okinawa Prefectural Government: Naha, Japan, 2006; p. 251. Available online: http://www.pref.okinawa.jp/site/kankyo/shizen/hogo/okinawa_rdb_kinrui_syokubutu.html (accessed on 1 March 2021). (In Japanese)
- Kyushu Regional Environmental Office of Ministry of the Environment (MoE). Notice of a Leaflet for Transfer Restrictions of Okinawa-Sekkoku (etc.). 2006. Available online: http://kyushu.env.go.jp/to_2006/0904a.html (accessed on 1 March 2021).
- Ministry of the Environment (MoE). Threatened Wildlife of Japan. In Red Data Book 2014; Gyosei Corporation: Tokyo, Japan, 2015; Volume 8, p. 347. (In Japanese) [Google Scholar]
- Abe, S.; Kotaka, N.; Takashima, A.; Abe, T.; Saito, K.; Masaki, T. Host selection and distribution of Dendrobium okinawense, an endangered epiphytic orchid in Yambaru, Japan. Ecol. Res. 2018, 33, 1069–1073. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Martin, K.P.; Madassery, J. Rapid in vitro propagation of Dendrobium hybrids through direct shoot formation from foliar explants, and protocorm-like bodies. Sci. Hort. 2006, 108, 95–99. [Google Scholar] [CrossRef]
- Habiba, S.; Ahasan, M.M.; Shimasaki, K. Effects of ethrel on organogenesis of protocorm-like bodies in Dendrobium kingianum in vitro. Plant Tissue Cult. Biotechnol. 2018, 28, 141–146. [Google Scholar] [CrossRef]
- Swamy, M.K.; Sudipta, K.M.; Balasubramanya, S.; Anuradha, M. Effect of different carbon sources on in vitro morphogenetic response of patchouli (Pogostemon cablin Benth.). J. Phytol. 2010, 2, 11–17. [Google Scholar]
- de Faria, R.T.; Rodrigues, F.N.; Olivera, L.V.R.; Muller, C. In vitro Dendrobium nobile plant growth and rooting in different sucrose concentrations. Hortic. Bras. 2004, 22, 780–783. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.J.; Murthy, H.N.; Hahn, E.J.; Paek, K.-Y. Biosmass production of Anoectochilus formosanus hayata in bioreactor system. J. Plant Biol. 2007, 50, 573–576. [Google Scholar] [CrossRef]
- Baque, M.A.; Shin, Y.-K.; Elshmari, T.; Lee, E.-J.; Paek, K.-Y. Effect of light quality, sucrose and coconut water concentration on the micropropagation of Calanthe hybrids (‘Bukduseong’ × ‘Hyesung’ and ‘Chunkwang’ × ‘Hyesung’). Aust. J. Crop Sci. 2011, 5, 1247–1254. [Google Scholar]
- Borisjuk, L.; Walenta, S.; Weber, H.; Rollerschek, H.; Mueller-Klieser, W.; Wobus, U.; Weber, H. Spatial analysis of plant metabolism: Sucrose imaging within Vicia faba in cotyledons reveals specific developmental patterns. Plant J. 2003, 29, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Chin, D.P.; Mishiba, K.; Mii, M. Agrobacterium-mediated transformation of protocorm-like bodies in Cymbidium. Plant Cell Rep. 2007, 26, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Li, J.; Li, B.; He, T.; Chun, Z. Effects of light quality on growth and development of protocorm-like bodies of Dendrobium officinale in vitro. Plant Cell Tissue Organ Cult. 2011, 105, 329–335. [Google Scholar] [CrossRef]
- Azmi, N.S.; Ahmad, R.; Ibrahim, R. Fluorescent light (FL), red led and blue led spectrums effects on in vitro shoots multiplication. J. Teknol. 2016, 78, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Manivannan, A.; Soundararajan, P.; Park, Y.G.; Wei, H.; Kim, S.H.; Jeong, B.R. Blue and red light-emitting diodes improve the growth and physiology of in vitro-grown carnations ‘Green Beauty’ and ‘Purple Beauty’. Hortic. Environ. Biotechnol. 2017, 58, 12–20. [Google Scholar] [CrossRef]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130243. [Google Scholar] [CrossRef]
- Reis, A.; Kleinowski, A.M.; Klein, F.R.S.; Telles, R.T.; do Amarante, L.; Braga, E.J.B. Light quality on the in vitro growth and production of pigments in the genus Alternanthera. J. Crop Sci. Biotechnol. 2015, 18, 349–357. [Google Scholar] [CrossRef]
- Bantis, F.; Ouzounis, T.; Radoglou, K. Artificial LED lighting enhances growth characteristics and total phenolic content of Ocimum basilicum, but variably affects transplant success. Sci. Hort. 2016, 198, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Miler, N.; Kulus, D.; Woźny, A.; Rymarz, D.; Hajzer, M.; Wierzbowski, K.; Nelke, R.; Szeffs, L. Application of wide-spectrum light-emitting diodes in micropropagation of popular ornamental plant species: A study on plant quality and cost reduction. In Vitro Cell. Dev. Biol. Plant 2019, 55, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Solymosi, K.; Schoefs, B. Etioplast and etio-chloroplast formation under natural conditions: The dark side of chlorophyll biosynthesis in angiosperms. Photosynth. Res. 2010, 105, 143–166. [Google Scholar] [CrossRef]
- Lee, S.-H.; Tewari, R.K.; Hahn, E.-J.; Paek, K.-Y. Photon flux and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. plantlets. Plant Cell Tissue Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Folta, K.M.; Maruhnich, S.A. Green light: A signal to slow down or stop. J. Expt. Bot. 2007, 58, 3099–3111. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Schleicher, E.; Meier, S.; Viana, R.M.; Pokorny, R.; Ahmad, M.; Bittl, R.; Batschauer, A. The signaling state of Arabidopsis scryptochrome 2 contains flavin semiquinone. J. Biol. Chem. 2007, 282, 14916–14922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Maruhnich, S.A.; Mageroy, M.H.; Justice, J.R.; Folta, K.M. Phototropin 1 and cryptochrome action in response to green light in combination with other wavelengths. Planta 2013, 237, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Goggin, D.E.; Steadman, K.J. Blue and green are frequently seen: Responses of seeds to short- and mid-wavelength light. Seed Sci. Res. 2012, 22, 27–35. [Google Scholar] [CrossRef]
- Bian, Z.-H.; Cheng, R.-F.; Yang, Q.-C.; Wang, J.; Lu, C. Continuous light from red, blue, and green light-emitting diodes reduces nitrate content and enhances phytochemical concentrations and antioxidant capacity in lettuce. J. Am. Soc. Hortic. Sci. 2016, 141, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Semiarti, E.; Purwantoro, A.; Indrianto, A. In vitro culture of orchids: The roles of class-1 Knox gene in shoot development. J. Biol. Res. 2014, 20, 18–27. [Google Scholar] [CrossRef]
- Find, J.; Grace, L.; Krogstrup, P. Effect of anti-auxins on maturation of embryogenic tissue cultures of Nordmanns fir (Abies nordmanniana). Physiol. Plant. 2002, 116, 231–237. [Google Scholar] [CrossRef]
- Shimasaki, K.; Uemoto, S. Micropropagation of a terrestrial Cymbidium species using rhizomes developed from seeds and pseudobulbs. Plant Cell Tissue Organ Cult. 1990, 22, 237–244. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) Different LEDs | |||
| LEDs | PLBs (%) | Shoot (%) | Shoot numbers |
| WF | 100.0 | 20.0 | 0.40 ± 0.27 a |
| R LED | 100.0 | 26.7 | 0.93 ± 0.50 a |
| W LED | 93.0 | 26.7 | 0.67 ± 0.33 a |
| B LED | 80.0 | 26.7 | 0.40 ± 0.19 a |
| G LED | 100.0 | 26.7 | 0.67 ± 0.41 a |
| (B) Different PCIB concentrations under white fluorescent light | |||
| PCIB (mg/L) | PLBs (%) | Shoot (%) | Shoot numbers |
| 0 | 100.0 | 40.0 | 2.07 ± 0.09 a |
| 0.01 | 100.0 | 40.0 | 2.87 ± 1.32 a |
| 0.1 | 100.0 | 6.7 | 0.40 ± 0.40 b |
| 1 | 86.7 | 13.3 | 0.67 ± 0.46 b |
| 10 | 93.3 | 6.7 | 0.27 ± 0.27 b |
| (C) Different HMI concentrations under white fluorescent light | |||
| HMI (mL/L) | PLBs (%) | Shoot (%) | Shoot numbers |
| 0 | 86.7 | 40.0 | 0.47 ± 0.17 a |
| 0.001 | 86.7 | 26.7 | 0.60 ± 0.32 a |
| 0.01 | 93.9 | 53.3 | 0.67 ± 0.21 a |
| 0.1 | 60.0 | 13.3 | 0.20 ± 0.14 a |
| (A) Culture media having PCIB (0.01 mg/L) | |||
| LEDs | PLBs (%) | Shoot (%) | Shoot numbers |
| WF | 100.0 | 20.0 | 0.33 ± 0.21 b |
| R LED | 93.3 | 26.7 | 0.47 ± 0.24 b |
| W LED | 86.7 | 40.0 | 0.67 ± 0.25 b |
| B LED | 86.7 | 33.3 | 0.60 ± 0.27 b |
| G LED | 100.0 | 33.3 | 1.53 ± 0.88 a |
| (B) Culture media having HMI (0.001 mL/L) | |||
| LEDs | PLBs (%) | Shoot (%) | Shoot numbers |
| WF | 100.0 | NSF | NSF |
| R LED | 73.3 | 13.3 | 0.13 ± 0.09 a |
| W LED | 46.7 | 13.3 | 0.13 ± 0.09 a |
| B LED | 100.0 | 6.7 | 0.20 ± 0.12 a |
| G LED | 93.3 | 20.0 | 0.47 ± 0.26 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehbub, H.; Shimasaki, K.; Mehraj, H. Low Concentration of Anti-Auxin and Anti-Fungal Agent Accelerates the PLB Regeneration of Dendrobium okinawense under Green LED. Plants 2022, 11, 1082. https://doi.org/10.3390/plants11081082
Mehbub H, Shimasaki K, Mehraj H. Low Concentration of Anti-Auxin and Anti-Fungal Agent Accelerates the PLB Regeneration of Dendrobium okinawense under Green LED. Plants. 2022; 11(8):1082. https://doi.org/10.3390/plants11081082
Chicago/Turabian StyleMehbub, Hasan, Kazuhiko Shimasaki, and Hasan Mehraj. 2022. "Low Concentration of Anti-Auxin and Anti-Fungal Agent Accelerates the PLB Regeneration of Dendrobium okinawense under Green LED" Plants 11, no. 8: 1082. https://doi.org/10.3390/plants11081082