
Dynamics of Bioactive Compounds during Spontaneous Fermentation of Paste Obtained from Capsicum ssp.—Stage towards a Product with Technological Application


 , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Moisture and Fruit Components Content
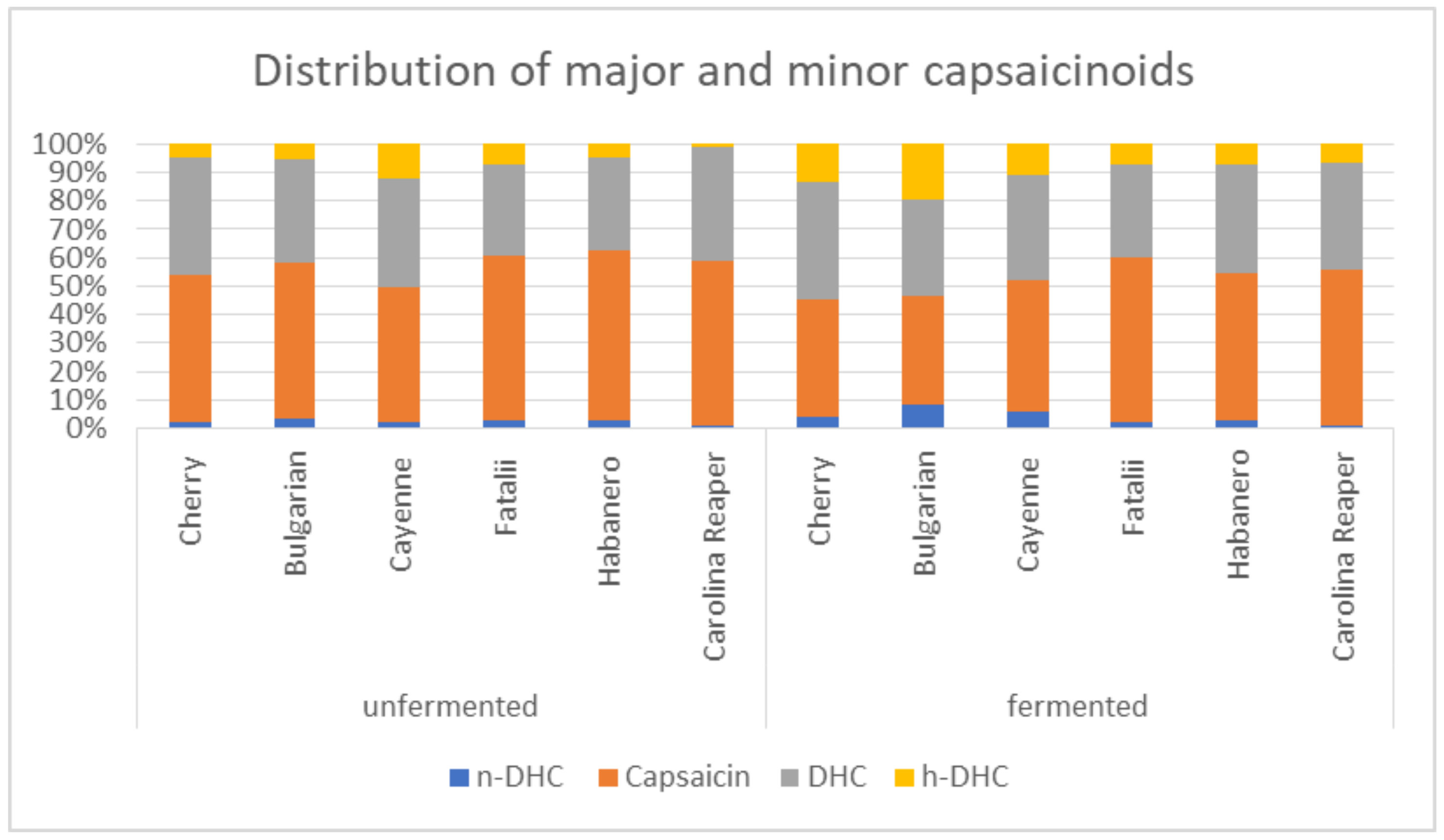
2.2. Capsaicinoids
2.3. Ascorbic Acid
2.4. Phenolic Compounds
2.5. Sugars and Acids
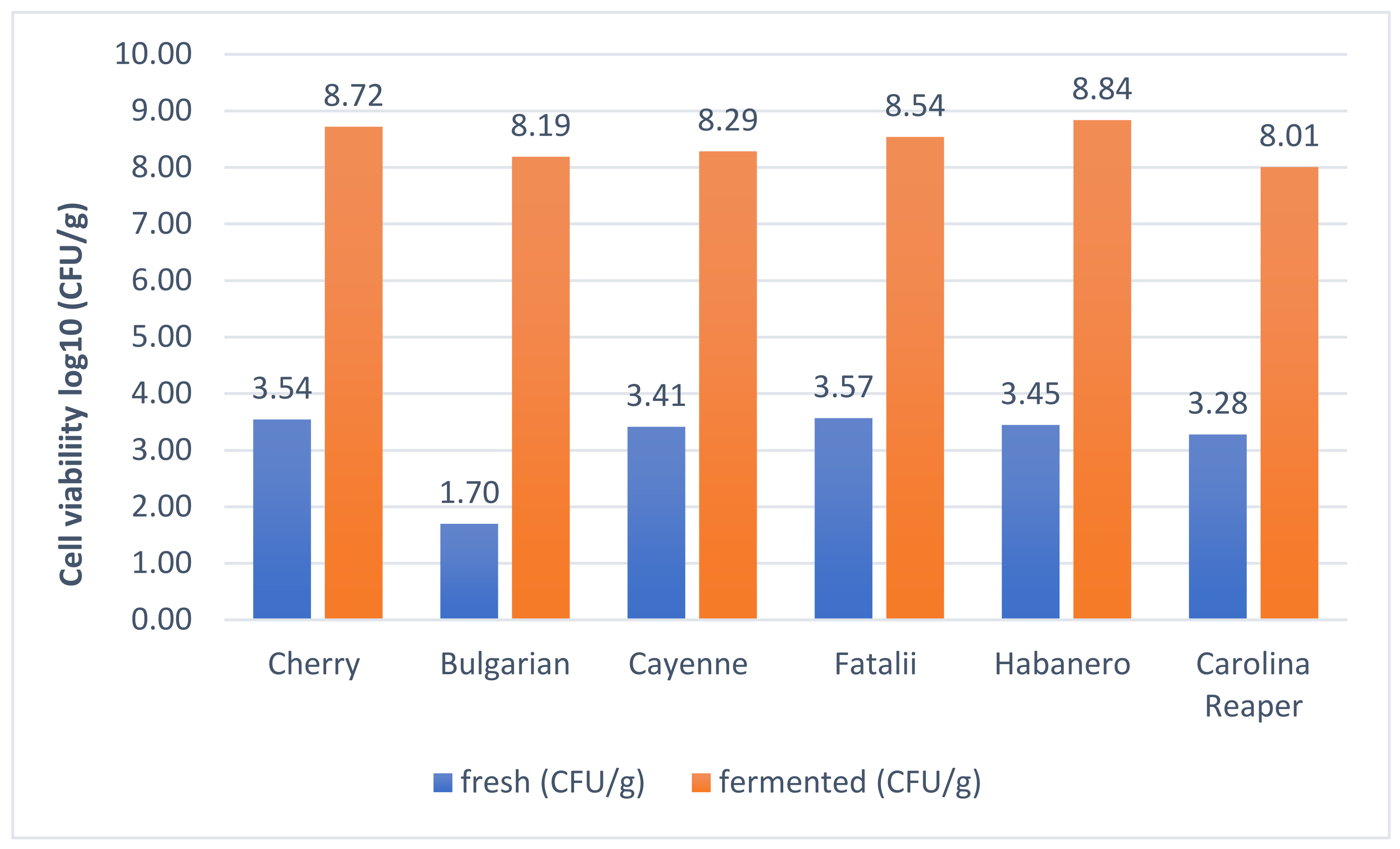
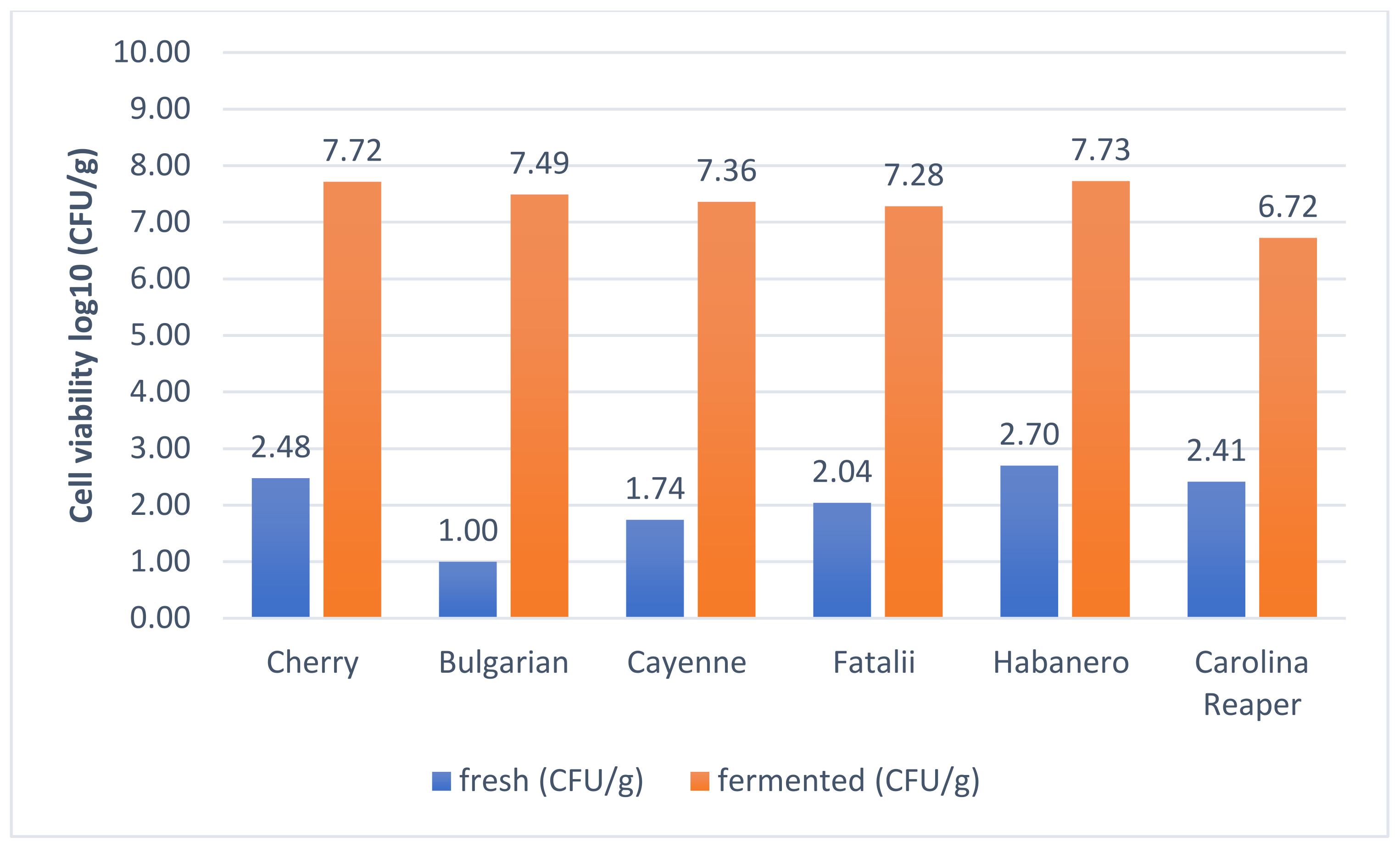
2.6. Viability Assay
3. Materials and Methods
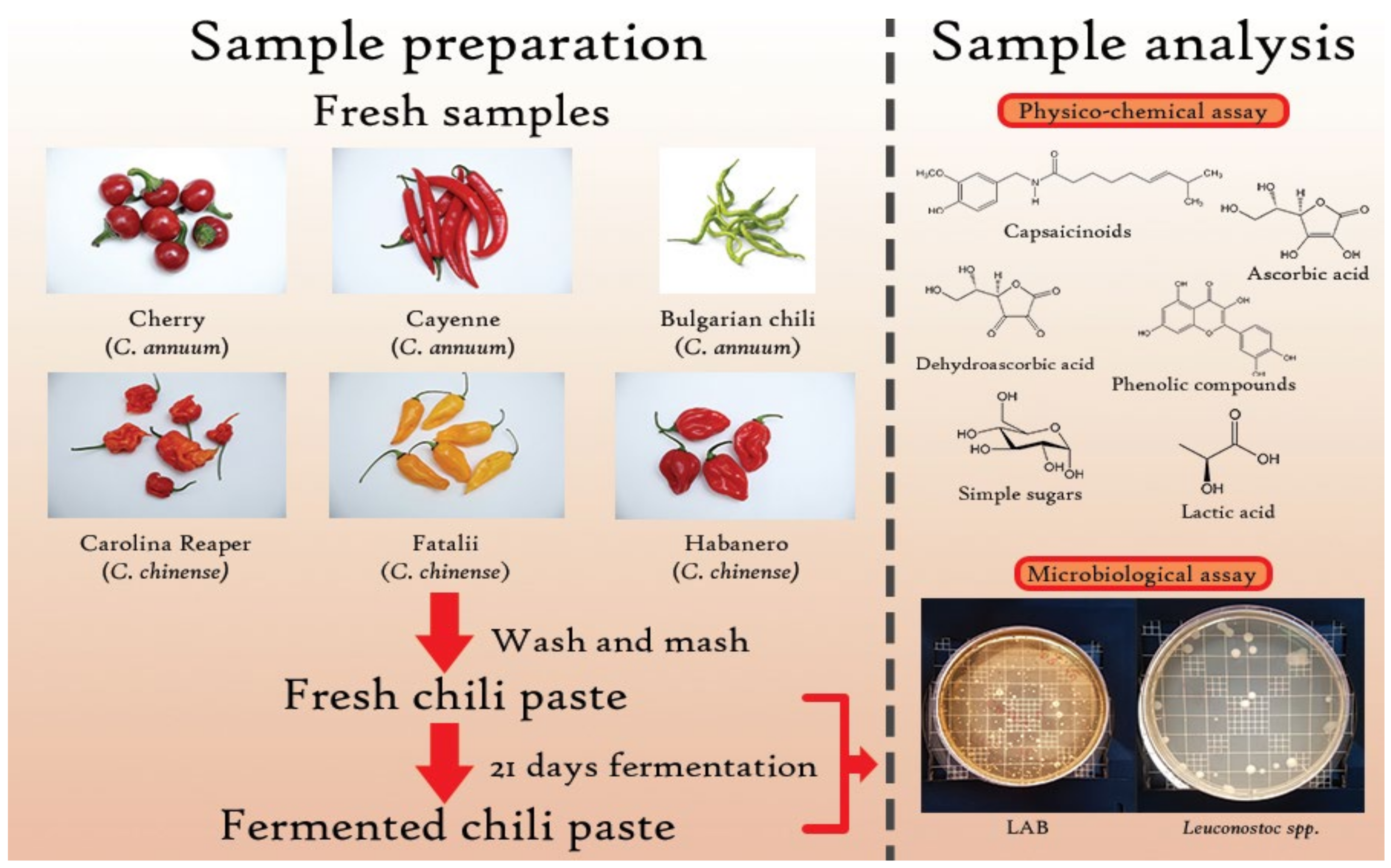
3.1. Sample Preparation
3.2. Sample Analysis
3.2.1. Ascorbic Acid and Dehydroascorbic Acid Quantification (HPLC-DAD-ESI+)
3.2.2. Phenolic Compounds and Capsaicinoids Extraction
3.2.3. Microbiological Analysis
3.2.4. Statistical Interpretation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| LAB | Lactic Acid Bacteria |
| UCP | Unfermented Chili Pastes |
| FCP | Fermented Chili Pastes |
| FW | Fresh Weight |
| DHC | Dihydrocapsaicin |
| n-DHC | Nordihydrocapsaicin |
| h-DHC | Homodihydrocapsaicin |
| DW | Dry Weight |
| SHU | Scoville Heat Units |
| ppm | parts per million |
| AAB | Acetic Acid Bacteria |
References
- Spence, J.D. Chili pepper consumption and cardiovascular mortality. J. Am. Coll. Cardiol. 2019, 74, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Bacon, K.; Boyer, R.; Denbow, C.; O’Keefe, S.; Neilson, A.; Williams, R. Antibacterial activity of jalapeño pepper (Capsicum annuum var. annuum) extract fractions against select foodborne pathogens. Food Sci. 2017, 5, 730–738. [Google Scholar]
- Valkova, V.; Duranova, H.; Ivanisova, E.; Galovicova, L.; Godocikova, L.; Borotova, P.; Kunova, S.; Miklasova, K.; Lopasovsky, L.; Mnahoncakova, E.; et al. Antioxidant and Antimicrobial Activities of Fruit Extracts from Different Fresh Chili Peppers. Acta Sci. Pol.-Technol. Aliment. 2021, 20, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Kirii, E.; Goto, T.; Yoshida, Y.; Yasuba, K.-i.; Tanaka, Y. Non-pungency in a Japanese Chili Pepper Landrace (Capsicum annuum) is Caused by a Novel Loss-of-function Pun1 Allele. Hortic. J. 2017, 86, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Antonio, A.S.; Wiedemann, L.S.M.; Veiga, V.F. The genus Capsicum: A phytochemical review of bioactive secondary metabolites. RSC Adv. 2018, 8, 25767–25784. [Google Scholar] [CrossRef] [Green Version]
- Erwin, A.T. Peppers; State College of Agriculture Experiment Station and Mechanic Arts: Des Moines, IA, USA, 1932. [Google Scholar]
- Mohd Hassan, N.; Yusof, N.A.; Yahaya, A.F.; Mohd Rozali, N.N.; Othman, R. Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes. Antioxidants 2019, 8, 469. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Perez, T.; Gomez-Garcia, M.D.R.; Valverde, M.E.; Paredes-Lopez, O. Capsicum annuum (hot pepper): An ancient Latin-American crop with outstanding bioactive compounds and nutraceutical potential. A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2972–2993. [Google Scholar] [CrossRef]
- Yang, F.; Zheng, J. Understand spiciness: Mechanism of TRPV1 channel activation by capsaicin. Protein Cell 2017, 8, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Escogido, M.D.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and Pharmacological Aspects of Capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef] [Green Version]
- Aza-Gonzalez, C.; Nunez-Palenius, H.G.; Ochoa-Alejo, N. Molecular biology of capsaicinoid biosynthesis in chili pepper (Capsicum spp.). Plant Cell Rep. 2011, 30, 695–706. [Google Scholar] [CrossRef]
- Stewart, C., Jr.; Mazourek, M.; Stellari, G.M.; O’Connell, M.; Jahn, M. Genetic control of pungency in C. chinense via the Pun1 locus. J. Exp. Bot. 2007, 58, 979–991. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Ismail, M.R.; Malek, M.A.; Latif, M.A. Capsaicin and Dihydrocapsaicin Determination in Chili Pepper Genotypes Using Ultra-Fast Liquid Chromatography. Molecules 2014, 19, 6474–6488. [Google Scholar] [CrossRef] [Green Version]
- Uarrota, V.G.; Maraschin, M.; de Bairros, A.F.M.; Pedreschi, R. Factors affecting the capsaicinoid profile of hot peppers and biological activity of their non-pungent analogs (Capsinoids) present in sweet peppers. Crit. Rev. Food Sci. Nutr. 2021, 61, 649–665. [Google Scholar] [CrossRef]
- Hamed, M.; Kalita, D.; Bartolo, M.E.; Jayanty, S.S. Capsaicinoids, Polyphenols and Antioxidant Activities of Capsicum annuum: Comparative Study of the Effect of Ripening Stage and Cooking Methods. Antioxidants 2019, 8, 364. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, K.; Murota, K.; Shimura, H.; Furuya, M.; Togawa, Y.; Matsumura, T.; Masuta, C. Evidence of capsaicin synthase activity of the Pun1-encoded protein and its role as a determinant of capsaicinoid accumulation in pepper. BMC Plant Biol. 2015, 15, 93. [Google Scholar] [CrossRef] [Green Version]
- Winter, J.; Bevan, S.; Campbell, E.A. Capsaicin and pain mechanisms. Br. J. Anaesth. 1995, 75, 157–168. [Google Scholar] [CrossRef] [Green Version]
- van Gorkom, G.N.Y.; Lookermans, E.L.; Van Elssen, C.; Bos, G.M.J. The Effect of Vitamin C (Ascorbic Acid) in the Treatment of Patients with Cancer: A Systematic Review. Nutrients 2019, 11, 977. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Kyndt, T.; Hancock, R.D. Vitamin C in Plants: Novel Concepts, New Perspectives, and Outstanding Issues. Antioxid. Redox Signal. 2020, 32, 463–485. [Google Scholar] [CrossRef]
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in Plants: From Functions to Biofortification. Antioxidants 2019, 8, 519. [Google Scholar] [CrossRef] [Green Version]
- Mahendran, S.; Bandara, D. Effects of soil moisture stress at different growth stages on vitamin C, capsaicin and β-carotene contents of chilli (Capsicum annuum L.) fruits and their impact on yield. Trop. Agric. Res. 2000, 12, 95–106. [Google Scholar]
- Washko, P.W.; Welch, R.W.; Dhariwal, K.R.; Wang, Y.; Levine, M. Ascorbic acid and dehydroascorbic acid analyses in biological samples. Anal. Biochem. 1992, 204, 1–14. [Google Scholar] [CrossRef]
- Deutsch, J.C. Dehydroascorbic acid. J. Chromatogr. A 2000, 881, 299–307. [Google Scholar] [CrossRef]
- Wilson, J.X. The physiological role of dehydroascorbic acid. FEBS Lett. 2002, 527, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Tolbert, B.M.; Ward, J.B. Dehydroascorbic Acid. Adv. Chem. Ser. 1982, 200, 101–123. [Google Scholar]
- May, J.M.; Cobb, C.E.; Mendiratta, S.; Hill, K.E.; Burk, R.F. Reduction of the ascorbyl free radical to ascorbate by thioredoxin reductase. J. Biol. Chem. 1998, 273, 23039–23045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zsuzsa, N. Chili Paprika Minőségi Paramétereinek Alakulása Különböző Színű Árnyékoló Hálók, Genotípusok és Érésfázisok Függvényében. Ph.D. Thesis, Szent István University, Gödöllő, Hungary, 2018. [Google Scholar]
- da Silveira Agostini-Costa, T.; da Silva Gomes, I.; de Melo, L.A.M.P.; Reifschneider, F.J.B.; da Costa Ribeiro, C.S. Carotenoid and total vitamin C content of peppers from selected Brazilian cultivars. J. Food Compos. Anal. 2017, 57, 73–79. [Google Scholar] [CrossRef]
- Shaha, R.K.; Rahman, S.; Asrul, A. Bioactive compounds in chilli peppers (Capsicum annuum L.) at various ripening (green, yellow and red) stages. Ann. Biol. Res. 2013, 4, 27–34. [Google Scholar]
- Simonne, A.; Simonne, E.; Eitenmiller, R.; Mills, H.; Green, N. Ascorbic acid and provitamin a contents in unusually colored bell peppers (Capsicum annuum L.). J. Food Compos. 1997, 10, 299–311. [Google Scholar] [CrossRef]
- Martinez, S.; Lopez, M.; Gonzalez-Raurich, M.; Alvarez, A.B. The effects of ripening stage and processing systems on vitamin C content in sweet peppers (Capsicum annuum L.). Int. J. Food Sci. Nutr. 2005, 56, 45–51. [Google Scholar] [CrossRef]
- Kubo, I.; Muroi, H.; Kubo, A. Structural functions of antimicrobial long-chain alcohols and phenols. Bioorg. Med. Chem. 1995, 3, 873–880. [Google Scholar] [CrossRef]
- Raccach, M. The antimicrobial activity of phenolic antioxidants in foods: A review. J. Food Saf. 1984, 6, 141–170. [Google Scholar] [CrossRef]
- De Sá Mendes, N.; de Andrade Gonçalves, É.C.B. The role of bioactive components found in peppers. Trends Food Sci. Technol. 2020, 99, 229–243. [Google Scholar] [CrossRef]
- Ashaolu, T.J. Immune boosting functional foods and their mechanisms: A critical evaluation of probiotics and prebiotics. Biomed. Pharmacother. 2020, 130, 110625. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, J.; Chen, L.; Hu, T.; Shi, L.; Wan, S.; Wang, M. Analysis and control of microbial gas production in fermented chili paste. J. Food Process. Preserv. 2020, 44, e14806. [Google Scholar] [CrossRef]
- Karasu, N.; Şimşek, Ö.; Çon, A.H. Technological and probiotic characteristics of Lactobacillus plantarum strains isolated from traditionally produced fermented vegetables. Ann. Microbiol. 2010, 60, 227–234. [Google Scholar] [CrossRef]
- Tomasik, P.; Tomasik, P. Probiotics, Non-Dairy Prebiotics and Postbiotics in Nutrition. Appl. Sci. 2020, 10, 1470. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.A.; Neto, J.H.P.L.; Cardarelli, H.R. Exopolysaccharides produced by Lactobacillus plantarum: Technological properties, biological activity, and potential application in the food industry. Ann. Microbiol. 2019, 69, 321–328. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, M.; Zhao, J.; Xia, Y.; Lai, P.F.; Ai, L. A Surface Protein From Lactobacillus plantarum Increases the Adhesion of Lactobacillus Strains to Human Epithelial Cells. Front. Microbiol. 2018, 9, 2858. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, S.A.; Salman, M.; Numan, M.; Javed, M.R.; Zubair, M.; Mustafa, G. Characterization of antifungal metabolites produced by Lactobacillus plantarum and Lactobacillus coryniformis isolated from rice rinsed water. Mol. Biol. Rep. 2020, 47, 1871–1881. [Google Scholar] [CrossRef]
- Guimaraes, A.; Santiago, A.; Teixeira, J.A.; Venancio, A. Anti-aflatoxigenic effect of organic acids produced by Lactobacillus plantarum. Int. J. Food Microbiol. 2018, 264, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervantes-Hernandez, F.; Alcala-Gonzalez, P.; Martinez, O.; Ordaz-Ortiz, J.J. Placenta, Pericarp, and Seeds of Tabasco Chili Pepper Fruits Show a Contrasting Diversity of Bioactive Metabolites. Metabolites 2019, 9, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillen, N.G.; Tito, R.; Mendoza, N.G. Capsaicinoids and pungency in Capsicum chinense and Capsicum baccatum fruits. Pesqui. Agropecu. Trop. 2018, 48, 237–244. [Google Scholar] [CrossRef]
- Do, T.L.Z.; Adam, V.; Zítka, O. Determination of the content of capsaicin and dihydrocapsaicin in twelve varieties of chilli peppers using liquid chromatography with UV/VIS detection. In Proceedings of the MendelNet 2017, Brno, Czech Republic, 8–9 November 2017. [Google Scholar]
- Duelund, L.; Mouritsen, O.G. Contents of capsaicinoids in chillies grown in Denmark. Food Chem. 2017, 221, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Koh, F.M. Physicochemical Properties of Pepper Mash Fermented in Wood and Plastic. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2005. [Google Scholar]
- de Aguiar, A.C.; Coutinho, J.P.; Barbero, G.F.; Godoy, H.T.; Martínez, J. Comparative Study of Capsaicinoid Composition in Capsicum Peppers Grown in Brazil. Int. J. Food Prop. 2015, 19, 1292–1302. [Google Scholar] [CrossRef] [Green Version]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef]
- Zamljen, T.; Jakopic, J.; Hudina, M.; Veberic, R.; Slatnar, A. Influence of intra and inter species variation in chilies (Capsicum spp.) on metabolite composition of three fruit segments. Sci. Rep. 2021, 11, 4932. [Google Scholar] [CrossRef]
- Agyemang Duah, S.; Silva e Souza, C.; Nagy, Z.; Pék, Z.; Neményi, A.; Daood, H.G.; Vinogradov, S.; Helyes, L. Effect of Water Supply on Physiological Response and Phytonutrient Composition of Chili Peppers. Water 2021, 13, 1284. [Google Scholar] [CrossRef]
- Teodoro, A.F.P.; Alves, R.D.N.; Ribeiro, L.B.; Reis, K.; Reifschneider, F.J.B.; Fonseca, M.E.D.; da Silva, J.P.; Agostini-Costa, T.D. Vitamin C content in Habanero pepper accessions (Capsicum chinense). Hortic. Bras. 2013, 31, 59–62. [Google Scholar] [CrossRef] [Green Version]
- Antonious, G.F.; Kochhar, T.S.; Jarret, R.L.; Snyder, J.C. Antioxidants in hot pepper: Variation among accessions. J. Environ. Sci. Health Part B-Pestic. Food Contam. Agric. Wastes 2006, 41, 1237–1243. [Google Scholar] [CrossRef]
- Roman-Arce, B.; Quiroz-Gonzalez, B.; Garcia-Mateos, R.; Ybarra-Moncada, C.; Corrales-Garcia, J. Nutraceutical value of fresh and dried chili (Capsicum annuum L.) in different stages of maturity. Bioagro 2019, 31, 123–132. [Google Scholar]
- Denev, P.; Todorova, V.; Ognyanov, M.; Georgiev, Y.; Yanakieva, I.; Tringovska, I.; Grozeva, S.; Kostova, D. Phytochemical composition and antioxidant activity of 63 Balkan pepper (Capsicum annuum L.) accessions. J. Food Meas. Charact. 2019, 13, 2510–2520. [Google Scholar] [CrossRef]
- Jamiołkowska, A.; Buczkowska, H.; Thanoon, A.H. Effect of biological preparations on the health state of pepper fruits and content of saccharides. Acta Sci. Pol. Hortorum Cultus 2016, 15, 95–107. [Google Scholar]
- Jarret, R.L.; Berke, T.; Baldwin, E.A.; Antonious, G.F. Variability for free sugars and organic acids in Capsicum chinense. Chem. Biodivers. 2009, 6, 138–145. [Google Scholar] [CrossRef]
- Redzepovic, S.; Orlic, S.; Majdak, A.; Kozina, B.; Volschenk, H.; Viljoen-Bloom, M. Differential malic acid degradation by selected strains of Saccharomyces during alcoholic fermentation. Int. J. Food Microbiol. 2003, 83, 49–61. [Google Scholar] [CrossRef]
- McFeeters, R.; Fleming, H.; Daeschel, M. Malic acid degradation and brined cucumber bloating. J. Food Sci. 1984, 49, 999–1002. [Google Scholar] [CrossRef]
- Shim, S.-M.; Kim, J.Y.; Lee, S.M.; Park, J.-B.; Oh, S.-K.; Kim, Y.-S. Profiling of fermentative metabolites in kimchi: Volatile and non-volatile organic acids. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 463–469. [Google Scholar] [CrossRef]
- Su, H.Y.; Li, H.Y.; Xie, C.Y.; Fei, Q.; Cheng, K.K. Co-production of acetoin and succinic acid by metabolically engineered Enterobacter cloacae. Biotechnol. Biofuels 2021, 14, 26. [Google Scholar] [CrossRef]
- Ferone, M.; Raganati, F.; Olivieri, G.; Marzocchella, A. Bioreactors for succinic acid production processes. Crit. Rev. Biotechnol. 2019, 39, 571–586. [Google Scholar] [CrossRef]
- Pederson, C.S.; Albury, M.N. The Sauerkraut Fermentation; Cornell University: Geneva, IL, USA, 1969. [Google Scholar]
- Hemme, D.; Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 2004, 14, 467–494. [Google Scholar] [CrossRef]
- Gonzalez-Quijano, G.K.; Dorantes-Alvarez, L.; Hernandez-Sanchez, H.; Jaramillo-Flores, M.E.; de Jesus Perea-Flores, M.; Vera-Ponce de Leon, A.; Hernandez-Rodriguez, C. Halotolerance and survival kinetics of lactic acid bacteria isolated from jalapeno pepper (Capsicum annuum L.) fermentation. J. Food Sci. 2014, 79, M1545–M1553. [Google Scholar] [CrossRef] [PubMed]
- Zubaidah, E.; Susanti, I.; Yuwono, S.; Rahayu, A.; Srianta, I.; Blanc, P. Effect of Lactobacillus plantarum and Leuconostoc mesenteroides starter cultures in lower salt concentration fermentation on the sauerkraut quality. Food Res. Int. 2020, 4, 1038–1044. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Pericarp (%) | Placenta (%) | Seeds (%) |
|---|---|---|---|
| Cherry | 75.42 ± 1.47 | 11.91 ± 0.25 | 12.67 ± 1.28 |
| Bulgarian | 90.68 ± 3.70 | 6.49 ± 3.12 | 2.82 ± 1.04 |
| Cayenne | 88.35 ± 1.06 | 8.90 ± 0.53 | 2.74 ± 1.37 |
| Fatalii | 80.61 ± 2.14 | 8.63 ± 1.50 | 10.75 ± 1.22 |
| Habanero | 86.54 ± 2.85 | 8.99 ± 1.61 | 4.47 ± 1.25 |
| Carolina Reaper | 86.27 ± 5.89 | 8.08 ± 2.36 | 5.65 ± 4.26 |
| Sample | Fresh | Fermented |
|---|---|---|
| Cherry | 85.48 ± 2.06 | 86.96 ± 0.59 |
| Bulgarian | 86.87 ± 1.75 | 87.98 ± 0.86 |
| Cayenne | 88.14 ± 1.02 | 90.34 ± 1.25 |
| Fatalii | 84.64 ± 0.40 | 88.30 ± 1.09 |
| Habanero | 88.58 ± 0.60 | 89.34 ± 0.39 |
| Carolina Reaper | 86.87 ± 0.02 | 89.06 ± 0.62 |
| Sample | n-DHC (μg/g) | Caps. (μg/g) | DHC (μg/g) | h-DHC (μg/g) | Total Capsaicinoids (μg/g) | |
|---|---|---|---|---|---|---|
| Fresh | Cherry | 9.62 ± 0.37 | 248.25 ± 2.01 | 198.60 ± 7.96 | 22.43 ± 1.72 | 478.90 ± 9.06 |
| Bulgarian | 5.46 ± 0.07 | 88.41 ± 2.96 | 58.31 ± 2.88 | 8.98 ± 0.12 | 161.15 ± 5.96 | |
| Cayenne | 5.14 ± 0.03 | 99.63 ± 5.18 | 79.13 ± 1.31 | 25.96 ± 0.90 | 209.84 ± 5.49 | |
| Fatalii | 117.56 ± 7.40 | 2365.16 ± 148.67 | 1317.76 ± 12.00 | 286.69 ± 6.11 | 4087.16 ± 154.61 | |
| Habanero | 21.47 ± 1.41 | 501.93 ± 3.60 | 277.40 ± 1.22 | 39.41 ± 0.50 | 840.21 ± 2.31 | |
| Carolina Reaper | 163.69 ± 9.87 | 9574.64 ± 269.15 | 6627.81 ± 177.99 | 191.87 ± 5.88 | 16,558.01 ± 313.89 | |
| Fermented | Cherry | 26.28 ± 0.92 | 262.02 ± 22.14 | 259.46 ± 3.07 | 85.53 ± 2.22 | 633.29 ± 24.30 |
| Bulgarian | 23.07 ± 1.01 | 104.11 ± 2.72 | 93.54 ± 1.00 | 52.86 ± 2.13 | 273.58 ± 4.83 | |
| Cayenne | 19.87 ± 1.02 | 152.16 ± 2.55 | 120.77 ± 4.87 | 35.88 ± 1.85 | 328.67 ± 6.62 | |
| Fatalii | 109.56 ± 3.75 | 2660.16 ± 30.01 | 1495.21 ± 40.88 | 343.06 ± 8.55 | 4607.98 ± 82.21 | |
| Habanero | 29.48 ± 2.41 | 547.74 ± 15.53 | 403.60 ± 3.16 | 77.52 ± 1.66 | 1058.33 ± 21.99 | |
| Carolina Reaper | 212.05 ± 15.21 | 9798.22 ± 159.50 | 6769.07 ± 60.84 | 1222.62 ± 22.61 | 18,001.96 ± 83.94 |
| Sample | Fresh | Fermented |
|---|---|---|
| Cherry | 51,156 ± 1224 | 71,463 ± 3304 |
| Bulgarian | 18,844 ± 723 | 31,835 ± 724 |
| Cayenne | 26,391 ± 946 | 50,279 ± 1489 |
| Fatalii | 406,524 ± 17,511 | 601,810 ± 10,611 |
| Habanero | 113,871 ± 825 | 151,563 ± 3146 |
| Carolina Reaper | 1,999,058 ± 55,578 | 2,536,507 ± 35,263 |
| Pepper Types | Ascorbic Acid | Dehydroascorbic Acid | Total Ascorbic Acid | |
|---|---|---|---|---|
| Fresh | Cherry | 159.41 ± 1.52 | 504.09 ± 1.99 | 663.50 ± 0.69 |
| Bulgarian | 224.80 ± 4.44 | 420.77 ± 17.14 | 645.58 ± 21.47 | |
| Cayenne | 954.41 ± 3.49 | 391.43 ± 10.25 | 1345.84 ± 9.26 | |
| Fatalii | 337.56 ± 16.55 | 416.69 ± 15.28 | 754.25 ± 28.97 | |
| Habanero | 360.93 ± 10.77 | 423.71 ± 21.34 | 784.64 ± 28.37 | |
| Carolina Reaper | 148.93 ± 1.73 | 417.63 ± 12.16 | 566.56 ± 13.15 | |
| Fermented | Cherry | 141.70 ± 1.52 | 384.30 ± 13.36 | 526.00 ± 13.98 |
| Bulgarian | 202.69 ± 2.56 | 362.61 ± 16.08 | 565.30 ± 18.49 | |
| Cayenne | 747.75 ± 20.62 | 330.44 ± 10.41 | 1078.18 ± 25.79 | |
| Fatalii | 297.32 ± 11.53 | 305.08 ± 13.24 | 602.40 ± 20.58 | |
| Habanero | 349.83 ± 11.07 | 301.20 ± 11.44 | 651.02 ± 17.12 | |
| Carolina Reaper | 136.35 ± 6.33 | 295.23 ± 19.94 | 431.58 ± 24.97 |
| Sample | Catechin- Derivatives | Vanillic Acid | Ferulic Acid Glucoside | Naringenin-Diglucoside | Luteolin- Apiosyl- Glucoside | Quercetin- Glucoside | Luteolin-(Apiosyl- Glucosyl-Malonyl)- Glucoside | Luteolin-Glucoside | Myricetin | Quercetin-(Galloyl- Caffeoyl-Glucosyl)- Rhamnoside | Luteolin | Naringin- Malonate | TPC | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fresh | Cherry | 44.22 ± 1.35 | 33.51 ± 3.26 | 114.82 ± 4.22 | 106.82 ± 2.84 | 50.37 ± 2.18 | 32.37 ± 3.65 | 208.83 ± 7.31 | 224.94 ± 5.04 | 12.85 ± 1.02 | 67.62 ± 1.51 | 15.31 ± 1.53 | 123.92 ± 2.96 | 1035.57 ± 20.16 |
| Bulgarian | 116.93 ± 3.54 | 55.66 ± 5.27 | 121.64 ± 6.03 | 81.39 ± 1.81 | 68.19 ± 2.54 | 137.56 ± 4.94 | 254.51 ± 5.00 | 568.76 ± 7.22 | 112.73 ± 6.18 | 86.39 ± 3.56 | 42.23 ± 2.55 | 308.31 ± 6.37 | 1954.30 ± 29.31 | |
| Cayenne | 74.10 ± 2.95 | 9.27 ± 0.27 | 53.41 ± 2.46 | 15.96 ± 1.70 | 19.67 ± 1.00 | 31.23 ± 3.84 | 58.15 ± 2.21 | 54.73 ± 0.64 | 28.77 ± 3.19 | 21.19 ± 1.08 | 7.92 ± 0.80 | 109.79 ± 5.83 | 484.18 ± 12.65 | |
| Fatalii | 169.87 ± 4.21 | 23.25 ± 1.84 | 24.79 ± 1.86 | 92.39 ± 1.53 | 6.02 ± 0.90 | 6.59 ± 0.40 | 33.51 ± 0.87 | 39.76 ± 2.35 | 12.47 ± 1.38 | 13.04 ± 0.97 | 0.00 ± 0.00 | 88.38 ± 2.74 | 510.06 ± 12.78 | |
| Habanero | 112.17 ± 2.00 | 17.89 ± 0.29 | 14.55 ± 2.00 | 23.54 ± 2.30 | 16.26 ± 2.01 | 19.86 ± 2.45 | 50.37 ± 2.02 | 50.94 ± 2.77 | 13.61 ± 1.95 | 14.36 ± 1.33 | 0.00 ± 0.00 | 35.29 ± 0.63 | 368.85 ± 10.36 | |
| Carolina Reaper | 154.40 ± 5.14 | 69.05 ± 1.77 | 62.51 ± 3.01 | 98.04 ± 4.27 | 85.06 ± 1.96 | 82.22 ± 4.04 | 110.08 ± 5.46 | 111.97 ± 6.05 | 58.15 ± 2.54 | 29.15 ± 1.01 | 0.00 ± 0.00 | 62.06 ± 1.92 | 922.68 ± 11.52 | |
| Fermented | Cherry | 212.99 ± 2.64 | 0.00 ± 0.00 | 21.00 ± 2.97 | 55.96 ± 1.02 | 208.45 ± 11.09 | 60.61 ± 2.96 | 38.24 ± 2.09 | 218.50 ± 7.49 | 6.97 ± 0.88 | 17.40 ± 1.28 | 41.66 ± 0.63 | 150.87 ± 5.22 | 1032.65 ± 15.58 |
| Bulgarian | 294.93 ± 3.45 | 15.81 ± 1.20 | 50.57 ± 5.07 | 62.65 ± 4.11 | 229.30 ± 7.40 | 96.81 ± 4.05 | 113.49 ± 6.50 | 591.13 ± 7.44 | 33.89 ± 2.93 | 64.59 ± 2.22 | 46.21 ± 2.01 | 204.66 ± 5.15 | 1804.03 ± 15.54 | |
| Cayenne | 115.29 ± 5.01 | 0.00 ± 0.00 | 6.97 ± 0.20 | 82.13 ± 2.55 | 42.79 ± 3.83 | 18.15 ± 0.33 | 14.74 ± 1.71 | 61.18 ± 1.20 | 5.26 ± 0.25 | 11.90 ± 2.02 | 12.85 ± 1.56 | 44.21 ± 2.63 | 415.49 ± 17.60 | |
| Fatalii | 157.67 ± 2.25 | 0.00 ± 0.00 | 7.16 ± 0.18 | 4.36 ± 1.04 | 11.90 ± 0.54 | 20.62 ± 0.28 | 15.31 ± 2.01 | 45.07 ± 2.65 | 17.96 ± 1.11 | 29.53 ± 0.52 | 5.08 ± 0.19 | 69.94 ± 5.42 | 384.59 ± 10.09 | |
| Habanero | 145.93 ± 4.06 | 0.84 ± 0.08 | 12.09 ± 1.40 | 16.55 ± 2.26 | 30.28 ± 0.35 | 17.21 ± 1.26 | 18.91 ± 2.06 | 42.98 ± 3.51 | 15.50 ± 1.00 | 16.45 ± 1.27 | 7.16 ± 0.14 | 33.80 ± 3.40 | 357.71 ± 8.40 | |
| Carolina Reaper | 185.63 ± 5.50 | 35.59 ± 2.47 | 32.75 ± 2.89 | 4.56 ± 0.21 | 61.56 ± 3.33 | 46.20 ± 3.29 | 31.61 ± 2.57 | 112.78 ± 2.49 | 28.77 ± 2.25 | 30.47 ± 0.71 | 42.79 ± 2.98 | 127.78 ± 10.28 | 740.50 ± 14.10 |
| Sample | Glucose | Fructose | Lactic Acid | Acetic Acid | Malic Acid | Citric | Succinic | |
|---|---|---|---|---|---|---|---|---|
| Fresh | Cherry | 17.07 ± 0.15 | 23.72 ± 0.43 | 0.28 ± 0.01 | 0.00 ± 0.00 | 3.60 ± 0.13 | 7.22 ± 0.03 | 0.21 ± 0.00 |
| Bulgarian | 16.47 ± 0.46 | 23.11 ± 0.34 | 0.32 ± 0.01 | 0.00 ± 0.00 | 3.17 ± 0.07 | 8.18 ± 0.04 | 0.22 ± 0.01 | |
| Cayenne | 19.40 ± 0.70 | 23.25 ± 0.07 | 0.17 ± 0.01 | 0.00 ± 0.00 | 2.92 ± 0.08 | 6.11 ± 0.20 | 0.18 ± 0.01 | |
| Fatalii | 13.55 ± 0.53 | 18.92 ± 0.60 | 0.08 ± 0.01 | 0.00 ± 0.00 | 1.45 ± 0.02 | 4.73 ± 0.10 | 0.16 ± 0.01 | |
| Habanero | 17.79 ± 0.37 | 22.62 ± 0.18 | 0.10 ± 0.01 | 0.00 ± 0.00 | 1.53 ± 0.08 | 5.15 ± 0.15 | 0.20 ± 0.01 | |
| Carolina Reaper | 12.86 ± 0.61 | 20.17 ± 0.29 | 0.08 ± 0.00 | 0.00 ± 0.00 | 1.22 ± 0.12 | 8.18 ± 0.15 | 0.20 ± 0.01 | |
| Fermented | Cherry | 0.34 ± 0.10 | 22.53 ± 0.18 | 11.33 ± 0.18 | 4.37 ± 0.02 | 0.94 ± 0.02 | 1.95 ± 0.03 | 0.66 ± 0.02 |
| Bulgarian | 0.04 ± 0.00 | 19.45 ± 0.10 | 13.06 ± 0.13 | 4.85 ± 0.05 | 0.02 ± 0.00 | 1.48 ± 0.11 | 0.91 ± 0.01 | |
| Cayenne | 0.86 ± 0.06 | 19.10 ± 0.12 | 12.15 ± 0.07 | 4.08 ± 0.04 | 0.34 ± 0.01 | 1.12 ± 0.12 | 0.56 ± 0.01 | |
| Fatalii | 2.34 ± 0.32 | 14.30 ± 0.05 | 10.34 ± 0.01 | 3.22 ± 0.01 | 0.18 ± 0.01 | 0.64 ± 0.02 | 1.35 ± 0.03 | |
| Habanero | 2.58 ± 0.06 | 17.25 ± 0.58 | 10.70 ± 0.05 | 3.29 ± 0.10 | 0.25 ± 0.01 | 0.52 ± 0.02 | 1.41 ± 0.01 | |
| Carolina Reaper | 0.32 ± 0.01 | 10.72 ± 0.35 | 10.32 ± 0.28 | 4.47 ± 0.12 | 0.01 ± 0.00 | 0.98 ± 0.06 | 1.92 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kádár, C.B.; Păucean, A.; Simon, E.; Vodnar, D.C.; Ranga, F.; Rusu, I.E.; Vișan, V.-G.; Man, S.; Chiș, M.S.; Drețcanu, G. Dynamics of Bioactive Compounds during Spontaneous Fermentation of Paste Obtained from Capsicum ssp.—Stage towards a Product with Technological Application. Plants 2022, 11, 1080. https://doi.org/10.3390/plants11081080
Kádár CB, Păucean A, Simon E, Vodnar DC, Ranga F, Rusu IE, Vișan V-G, Man S, Chiș MS, Drețcanu G. Dynamics of Bioactive Compounds during Spontaneous Fermentation of Paste Obtained from Capsicum ssp.—Stage towards a Product with Technological Application. Plants. 2022; 11(8):1080. https://doi.org/10.3390/plants11081080
Chicago/Turabian StyleKádár, Csaba Balázs, Adriana Păucean, Elemér Simon, Dan Cristian Vodnar, Floricuța Ranga, Iulian Eugen Rusu, Vasile-Gheorghe Vișan, Simona Man, Maria Simona Chiș, and Georgiana Drețcanu. 2022. "Dynamics of Bioactive Compounds during Spontaneous Fermentation of Paste Obtained from Capsicum ssp.—Stage towards a Product with Technological Application" Plants 11, no. 8: 1080. https://doi.org/10.3390/plants11081080