
Combinational Variation Temperature and Soil Water Response of Stomata and Biomass Production in Maize, Millet, Sorghum and Rice

 ,
,
Abstract
:1. Introduction
2. Results
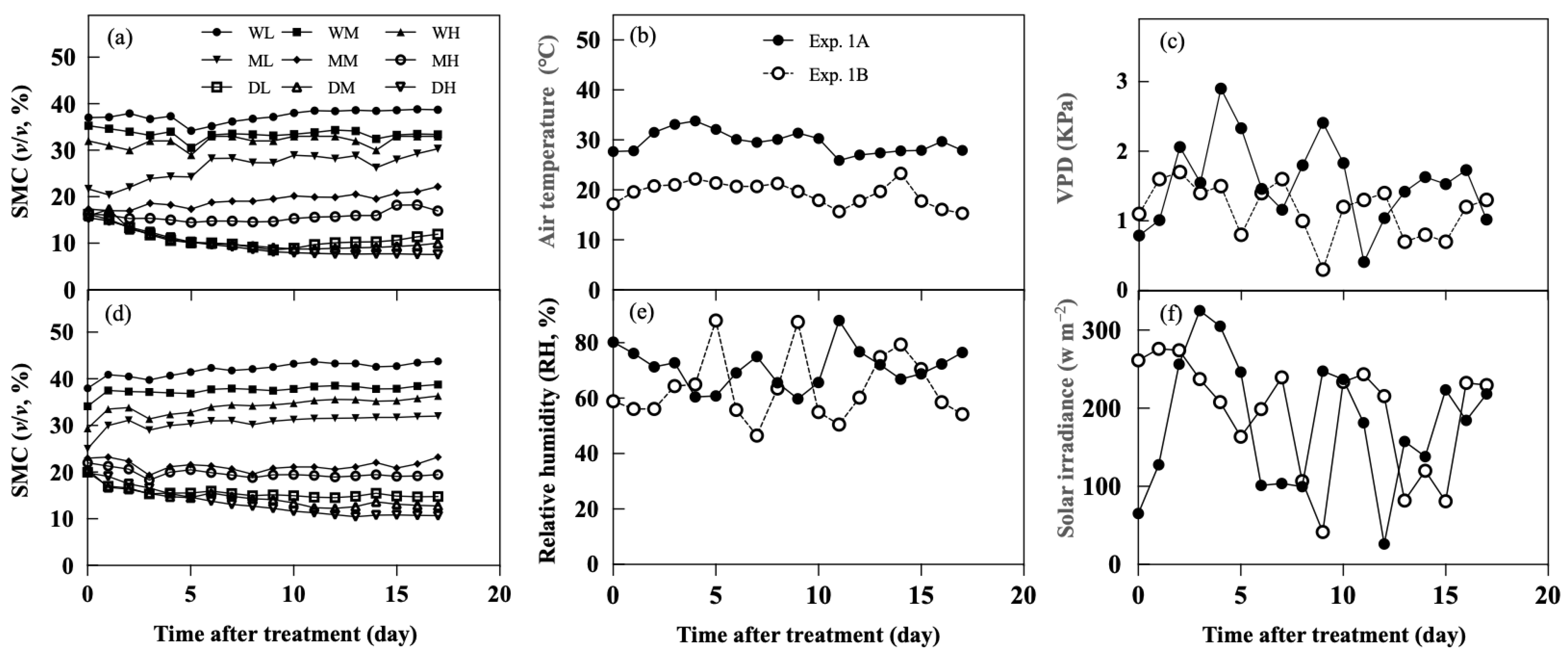
2.1. Experiment 1
2.1.1. Soil Control and Atmospheric Environment
2.1.2. The Correlation between Soil Moisture Status and Shoot Biomass, LA, and gs
2.2. Experiment 2
2.2.1. Crop Response to a Combination of Soil Moisture Status and Temperature on Shoot Biomass, LA, and Gas Exchange
2.2.2. Changing of Gas Exchange
2.2.3. Correlation between gs, A, and E and Influence of Atmospheric Environment and gs on Shoot Biomass
3. Discussion
3.1. The gs Responses to Soil Moisture Status and Environmental Influence on Biomass Production
3.2. A Plant’s Ability to Maintain Gas Exchange Is Important for Maintaining the Biomass Production
3.3. The Influence of Soil Moisture Content, Air Temperature and gs on Shoot Biomass of Each Crop
4. Materials and Methods
4.1. Seedling Preparation
4.2. Experiment 1: Four Crops on Nine Different Water Conditions
4.2.1. Experimental Site
4.2.2. Treatment
4.2.3. Soil moisture Content, Leaf Area, Shoot Biomass, and gs
4.3. Experiment 2: The Effect of Soil Water Statuses and Temperature Combination on Four Crops
4.3.1. Experimental Site and Growing Media Preparation
4.3.2. Method and Treatment
4.3.3. Shoot Biomass, LA, A, gs, and E
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beillouin, D.; Schauberger, B.; Bastos, A.; Ciais, P.; Makowski, D. Impact of extreme weather conditions on European crop production in 2018. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190510. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, G.; Ahmed, K.F.; Adugna, B.; Eggen, M.; Atsbeha, E.; You, L.; Koo, J.; Anagnostou, E. The role of climate in the trend and variability of Ethiopia’s cereal crop yields. Sci. Total Environ. 2020, 723, 137893. [Google Scholar] [CrossRef] [PubMed]
- Pickson, R.B.; He, G.; Ntiamoah, E.B.; Li, C. Cereal production in the presence of climate change in China. Environ. Sci. Pollut. Res. 2020, 27, 45802–45813. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Statistic Database; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Giorgi, F.; Raffaele, F.; Coppola, E. The response of precipitation characteristics to global warming from climate projections. Earth Syst. Dyn. 2019, 10, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Berg, A.; De Noblet-Ducoudre, N.; Sultan, B.; Lengaigne, M.; Guimberteau, M. Projections of climate change impacts on potential C4 crop productivity over tropical regions. Agric. For. Meteorol. 2013, 170, 89–102. [Google Scholar] [CrossRef]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and Drought Stresses in Crops and Approaches for Their Mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef]
- Maulana, F.; Tesso, T.T. Cold Temperature Episode at Seedling and Flowering Stages Reduces Growth and Yield Components in Sorghum. Crop Sci. 2013, 53, 564–574. [Google Scholar] [CrossRef]
- Matsuura, A.; An, P.; Murata, K.; Inanaga, S. Effect of pre- and post-heading waterlogging on growth and grain yield of four millets. Plant Prod. Sci. 2016, 19, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Vanga, S.K.; Saxena, R.; Orsat, V.; Raghavan, V. Effect of Climate Change on the Yield of Cereal Crops: A Review. Climate 2018, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.C.; Cowan, I.R.; Farquhar, G. Stomatal conductance correlates with photosynthetic capacity. Nature 1979, 282, 424–426. [Google Scholar] [CrossRef]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.M.; Condon, A.G.; Saavedra, A.L. Wheat Yield Progress Associated with Higher Stomatal Conductance and Photosynthetic Rate, and Cooler Canopies. Crop. Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Blatt, M.R. Cellular Signaling and Volume Control in Stomatal Movements in Plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Struik, P.C.; Yin, X.; Yang, J. Stomatal conductance, mesophyll conductance, and transpiration efficiency in relation to leaf anatomy in rice and wheat genotypes under drought. J. Exp. Bot. 2017, 68, 5191–5205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, T.; Abe, F.; Tsutsumi, N.; Nakazono, M. Root Cortex Provides a Venue for Gas-Space Formation and Is Essential for Plant Adaptation to Waterlogging. Front. Plant Sci. 2019, 10, 259. [Google Scholar] [CrossRef]
- Yamauchi, T.; Pedersen, O.; Nakazono, M.; Tsutsumi, N. Key root traits of Poaceae for adaptation to soil water gradients. New Phytol. 2020, 229, 3133–3140. [Google Scholar] [CrossRef]
- Sakagami, J.-I.; Kawano, N. Survival of submerged rice in a flood-prone region of West Africa. Tropics 2011, 20, 55–66. [Google Scholar] [CrossRef]
- Lizarazu, W.Z.; Iijima, M. Deep Root Water Uptake Ability and Water Use Efficiency of Pearl Millet in Comparison to Other Millet Species. Plant Prod. Sci. 2005, 8, 454–460. [Google Scholar] [CrossRef]
- Khanthavong, P.; Yabuta, S.; Asai, H.; Hossain, M.A.; Akagi, I.; Sakagami, J.-I. Root Response to Soil Water Status via Interaction of Crop Genotype and Environment. Agronomy 2021, 11, 708. [Google Scholar] [CrossRef]
- Galamay, T.O.; Kono, Y.; Yamauchi, A.; Shimizu, M. Comparative root anatomy of seminal and nodal root axes of summer cereals with special reference to the development of hypodermis and cortical sclerenchyma. Jpn. J. Crop Sci. 1991, 60, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Parent, C.; Capelli, N.; Berger, A.; Crevecoeur, M.; Dat, J.F. An overview of plant responses to soil waterlogging. Plant Stress 2008, 2, 20–27. [Google Scholar]
- Zhang, F.; Zhu, K.; Wang, Y.; Zhang, Z.; Lu, F.; Yu, H.; Zou, J. Changes in photosynthetic and chlorophyll fluorescence characteristics of sorghum under drought and waterlogging stress. Photosynthetica 2019, 57, 1156–1164. [Google Scholar] [CrossRef] [Green Version]
- Promkhambu, A.; Younger, A.; Polthanee, A.; Akkasaeng, C. Morphological and Physiological Responses of Sorghum (Sorghum bicolor L. Moench) to Waterlogging. Asian J. Plant Sci. 2010, 9, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Li, J.; Bi, W.; Zuo, S.; Li, L.; Li, W.; Sun, L. Effects of waterlogging stress at different growth stages on the photosynthetic characteristics and grain yield of spring maize (Zea mays L.) Under field conditions. Agric. Water Manag. 2019, 218, 250–258. [Google Scholar] [CrossRef]
- Matsuura, A.; Kato, Y.; Suzuki, T.; Murata, K.; An, P. Hypoxia tolerance of four millet species is attributable to constitutive aerenchyma formation and root hair development of adventitious roots. Plant Prod. Sci. 2022, 1–15. [Google Scholar] [CrossRef]
- Pardales, J., Jr.; Kono, Y.; Yamauchi, A. Response of the different root system components of sorghum to incidence of waterlogging. Environ. Exp. Bot. 1991, 31, 107–115. [Google Scholar] [CrossRef]
- Pareek, A.; Sopory, S.K.; Bohnert, H. Abiotic Stress Adaptation in Plants; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. Sustain. Agric. 2009, 29, 185–212. [Google Scholar]
- Hadebe, S.T.; Modi, A.T.; Mabhaudhi, T. Drought Tolerance and Water Use of Cereal Crops: A Focus on Sorghum as a Food Security Crop in Sub-Saharan Africa. J. Agron. Crop Sci. 2016, 203, 177–191. [Google Scholar] [CrossRef]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophysics 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Munns, R.; James, R.; Sirault, X.; Furbank, R.; Jones, H. New phenotyping methods for screening wheat and barley for beneficial responses to water deficit. J. Exp. Bot. 2010, 61, 3499–3507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludlow, M.; Santamaria, J.; Fukai, S. Contribution of osmotic adjustment to grain yield in Sorghum bicolor (L.) Moench under water-limited conditions. II. Water stress after anthesis. Aust. J. Agric. Res. 1990, 41, 67–78. [Google Scholar] [CrossRef]
- Wang, X.; Du, T.; Huang, J.; Peng, S.; Xiong, D. Leaf hydraulic vulnerability triggers the decline in stomatal and mesophyll conductance during drought in rice. J. Exp. Bot. 2018, 69, 4033–4045. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Glob. Chang. Biol. 2013, 20, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Sood, S.; Khulbe, R.K.; Gupta, A.K.; Agrawal, P.K.; Upadhyaya, H.D.; Bhatt, J.C. Barnyard millet—A potential food and feed crop of future. Plant Breed. 2015, 134, 135–147. [Google Scholar] [CrossRef]
- Osborne, C.P.; Wythe, E.J.; Ibrahim, U.G.; Gilbert, M.; Ripley, B.S. Low temperature effects on leaf physiology and survivorship in the C3 and C4 subspecies of Alloteropsis semialata. J. Exp. Bot. 2007, 59, 1743–1754. [Google Scholar] [CrossRef] [Green Version]
- Crafts-Brandner, S.J.; Salvucci, M.E.; Schultz, C.J.; Rumsewicz, M.P.; Johnson, K.L.; Jones, B.J.; Gaspar, Y.M.; Bacic, A. Sensitivity of Photosynthesis in a C4 Plant, Maize, to Heat Stress. Plant Physiol. 2002, 129, 1773–1780. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Zhou, Y.; Yue, Z.; Chen, X.; Cao, X.; Ai, X.; Jiang, B.; Xing, Y. The leaf-air temperature difference reflects the variation in water status and photosynthesis of sorghum under waterlogged conditions. PLoS ONE 2019, 14, e0219209. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.T.; Yao, Y.T.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Climate change 2014: Synthesis report. In Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Chan; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Lawson, T.; Blatt, M.R.; Yu, L.; Shi, D.; Li, J.; Kong, Y.; Yu, Y.; Chai, G.; Hu, R.; Wang, J.; et al. Stomatal Size, Speed, and Responsiveness Impact on Photosynthesis and Water Use Efficiency. Plant Physiol. 2014, 164, 1556–1570. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, D.W. Low Temperature Effects on Early Vegetative Growth, Leaf Gas Exchange and Water Potential of Chilling-sensitive and Chilling-tolerant Crop Species*. Ann. Bot. 1991, 67, 205–212. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and Drought Stresses in Crop Plants: Implications, Cross Talk, and Potential Management Opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Wilkinson, S.; Clephan, A.L.; Davies, W.J. Rapid Low Temperature-Induced Stomatal Closure Occurs in Cold-Tolerant Commelina communis Leaves But Not in Cold-Sensitive Tobacco Leaves, via a Mechanism That Involves Apoplastic Calcium But Not Abscisic Acid. Plant Physiol. 2001, 126, 1566–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assmann, S.M.; Shimazaki, K.-I. The Multisensory Guard Cell. Stomatal Responses to Blue Light and Abscisic Acid1. Plant Physiol. 1999, 119, 809–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyne, E.G.; Brunson, A.M. Genetic Studies of Heat and Drought Tolerance in Maize 1. Agron. J. 1940, 32, 803–814. [Google Scholar] [CrossRef]
- Craufurd, P.Q.; Peacock, J.M. Effect of Heat and Drought Stress on Sorghum (Sorghum Bicolor). II. Grain Yield. Exp. Agric. 1993, 29, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When Defense Pathways Collide. The Response of Arabidopsis to a Combination of Drought and Heat Stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Bajwa, A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.; Xu, S.-Z.; Zou, X.-L.; Zheng, Y.-L.; Qiu, F.-Z. Changes of Antioxidative Enzymes and Lipid Peroxidation in Leaves and Roots of Waterlogging-Tolerant and Waterlogging-Sensitive Maize Genotypes at Seedling Stage. Agric. Sci. China 2010, 9, 651–661. [Google Scholar] [CrossRef]
- Becklin, K.M.; Ward, J.K.; Way, D.A. Photosynthesis, Respiration, and Climate Change; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Orzechowska, A.; Trtílek, M.; Tokarz, K.; Szymańska, R.; Niewiadomska, E.; Rozpądek, P.; Wątor, K. Thermal Analysis of Stomatal Response under Salinity and High Light. Int. J. Mol. Sci. 2021, 22, 4663. [Google Scholar] [CrossRef]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef]
- Ren, B.; Zhang, J.; Li, X.; Fan, X.; Dong, S.; Liu, P.; Zhao, B. Effects of waterlogging on the yield and growth of summer maize under field conditions. Can. J. Plant Sci. 2014, 94, 23–31. [Google Scholar] [CrossRef]
- Abiko, T.; Kotula, L.; Shiono, K.; Malik, A.I.; Colmer, T.D.; Nakazono, M. Enhanced formation of aerenchyma and induction of a barrier to radial oxygen loss in adventitious roots of Zea nicaraguensis contribute to its waterlogging tolerance as compared with maize (Zea mays ssp. mays). Plant Cell Environ. 2012, 35, 1618–1630. [Google Scholar] [CrossRef]
- Promkhambut, A.; Polthanee, A.; Akkasaeng, C.; Younger, A. Growth, yield and aerenchyma formation of sweet and multipurpose sorghum (Sorghum bicolor L. Moench) as affected by flooding at different growth stages. Aust. J. Crop Sci. 2011, 5, 954–965. [Google Scholar]
- Neira, J.; Ortiz, M.; Morales, L.; Acevedo, E. Oxygen diffusion in soils: Understanding the factors and processes needed for modeling. Chil. J. Agric. Res. 2015, 75, 35–44. [Google Scholar] [CrossRef]
- Yamauchi, T.; Noshita, K.; Tsutsumi, N. Climate-smart crops: Key root anatomical traits that confer flooding tolerance. Breed. Sci. 2021, 71, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, P.; Armstrong, W. Soil waterlogging, root rot and conifer windthrow: Oxygen deficiency or phytoxicity? Plant Soil 1978, 49, 185–190. [Google Scholar] [CrossRef]
- Kimura, H.; Hashimoto-Sugimoto, M.; Iba, K.; Terashima, I.; Yamori, W. Improved stomatal opening enhances photosynthetic rate and biomass production in fluctuating light. J. Exp. Bot. 2020, 71, 2339–2350. [Google Scholar] [CrossRef]
- Nagai, T.; Makino, A. Differences between Rice and Wheat in Temperature Responses of Photosynthesis and Plant Growth. Plant Cell Physiol. 2009, 50, 744–755. [Google Scholar] [CrossRef] [Green Version]
- Sage, R.F.; Kubien, D.S. The temperature response of C3and C4photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef]
- Huner, N.P.A.; Migus, W.; Tollenaar, M. Leaf CO2 Exchange Rates in Winter Rye Grown at Cold-Hardening And Nonhardening Temperatures. Can. J. Plant Sci. 1986, 66, 443–452. [Google Scholar] [CrossRef]
- Yamasaki, T.; Yamakawa, T.; Yamane, Y.; Koike, H.; Satoh, K.; Katoh, S. Temperature Acclimation of Photosynthesis and Related Changes in Photosystem II Electron Transport in Winter Wheat. Plant Physiol. 2002, 128, 1087–1097. [Google Scholar] [CrossRef] [Green Version]
- Hikosaka, K.; Ishikawa, K.; Borjigidai, A.; Muller, O.; Onoda, Y. Temperature acclimation of photosynthesis: Mechanisms involved in the changes in temperature dependence of photosynthetic rate. J. Exp. Bot. 2005, 57, 291–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, T.; Simkin, A.J.; Kelly, G.; Granot, D. Mesophyll photosynthesis and guard cell metabolism impacts on stomatal behaviour. New Phytol. 2014, 203, 1064–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, S.; Yuan, Z.; Zhang, Y.; Liu, W.; Zhang, L.; Huang, J.; Wan, S. Photosynthetic responses of C3 and C4 species to seasonal water variability and competition. J. Exp. Bot. 2005, 56, 2867–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyer, E.; Colin-Belgrand, M.; Biron, P. Photosynthesis and shoot water status of seedlings from different oak species submitted to waterlogging. Ann. Sci. For. 1991, 48, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Souza, R.; Machado, E.; Silva, J.; Lagôa, A.; Silveira, J. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Kurepin, L.V.; Reid, D.M. Effects of temperature and watering regime on growth, gas exchange and abscisic acid content of canola (Brassica napus) seedlings. Environ. Exp. Bot. 2012, 75, 107–113. [Google Scholar] [CrossRef]
- Rodríguez-Gamir, J.; Xue, J.; Clearwater, M.J.; Meason, D.; Clinton, P.W.; Domec, J. Aquaporin regulation in roots controls plant hydraulic conductance, stomatal conductance, and leaf water potential in Pinus radiata under water stress. Plant Cell Environ. 2019, 42, 717–729. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef]
- Bansal, R.; Srivastava, J.P. Effect of waterlogging on photosynthetic and biochemical parameters in pigeonpea. Russ. J. Plant Physiol. 2015, 62, 322–327. [Google Scholar] [CrossRef]
- Hayashi, T.; Yoshida, T.; Fujii, K.; Mitsuya, S.; Tsuji, T.; Okada, Y.; Hayashi, E.; Yamauchi, A. Maintained root length density contributes to the waterlogging tolerance in common wheat (Triticum aestivum L.). Field Crop. Res. 2013, 152, 27–35. [Google Scholar] [CrossRef]
- Goto, K.; Yabuta, S.; Ssenyonga, P.; Tamaru, S.; Sakagami, J.-I. Response of leaf water potential, stomatal conductance and chlorophyll content under different levels of soil water, air vapor pressure deficit and solar radiation in chili pepper (Capsicum chinense). Sci. Hortic. 2021, 281, 109943. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Shoot Biomass | Leaf Area | A | gs | E | WUE |
|---|---|---|---|---|---|---|
| Crops | ns | ns | ns | ns | ns | ns |
| Soil water status (SWS) | *** | *** | *** | *** | *** | *** |
| Crops × soil water status | *** | *** | *** | *** | *** | *** |
| Maize | Sorghum | Millet | Rice | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Equation | Variation | β | t-Value | Adjusted R2 | p-Value | Equation | β | t-value | Adjusted R2 | p-Value | Equation | β | t-Value | Adjusted R2 | p-Value | Equation | β | t-Value | Adjusted R2 | p-Value |
| (1) | Intercept | 5.71 | 2.906 | 0.658 | 0.000 | (2) | 0.947 | 1.351 | 0.759 | 0.000 | (3) | −9.832 | −3.072 | 0.492 | 0.006 | (4) | 0.556 | 1.794 | 0.262 | 0.066 |
| Temp | 0.071 | 0.983 | 0.163 | 4.875 | 0.455 | 2.514 | −0.006 | −0.567 | ||||||||||||
| SMC | −0.171 | −3.271 | −0.085 | −4.602 | 0.208 | 2.126 | −0.008 | −0.896 | ||||||||||||
| gs | 0.012 | 1.877 | −0.005 | −1.293 | −0.016 | −0.750 | 0.001 | 2.231 | ||||||||||||
| No. | Abbreviation | Soil Water Statuses (Treatments) |
|---|---|---|
| 1 | WL | Low position of sub-wet soil conditions (waterlogging) |
| 2 | WM | Middle position of sub-wet soil conditions |
| 3 | WH | High position of sub-wet soil conditions |
| 4 | ML | Low position of sub-moderate soil conditions |
| 5 | MM | Middle position of sub-moderate soil conditions |
| 6 | MH | High position of sub-moderate soil conditions |
| 7 | DL | Low position of sub-dry soil conditions |
| 8 | DM | Middle position of sub-dry soil conditions |
| 9 | DH | High position of sub-dry soil conditions (severe dry soil) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khanthavong, P.; Yabuta, S.; Malik, A.I.; Hossain, M.A.; Akagi, I.; Sakagami, J.-I. Combinational Variation Temperature and Soil Water Response of Stomata and Biomass Production in Maize, Millet, Sorghum and Rice. Plants 2022, 11, 1039. https://doi.org/10.3390/plants11081039
Khanthavong P, Yabuta S, Malik AI, Hossain MA, Akagi I, Sakagami J-I. Combinational Variation Temperature and Soil Water Response of Stomata and Biomass Production in Maize, Millet, Sorghum and Rice. Plants. 2022; 11(8):1039. https://doi.org/10.3390/plants11081039
Chicago/Turabian StyleKhanthavong, Phanthasin, Shin Yabuta, Al Imran Malik, Md Amzad Hossain, Isao Akagi, and Jun-Ichi Sakagami. 2022. "Combinational Variation Temperature and Soil Water Response of Stomata and Biomass Production in Maize, Millet, Sorghum and Rice" Plants 11, no. 8: 1039. https://doi.org/10.3390/plants11081039