
Diversity and Content of Carotenoids and Other Pigments in the Transition from the Green to the Red Stage of Haematococcus pluvialis Microalgae Identified by HPLC-DAD and LC-QTOF-MS

 ,
,
Abstract
:1. Introduction
2. Results
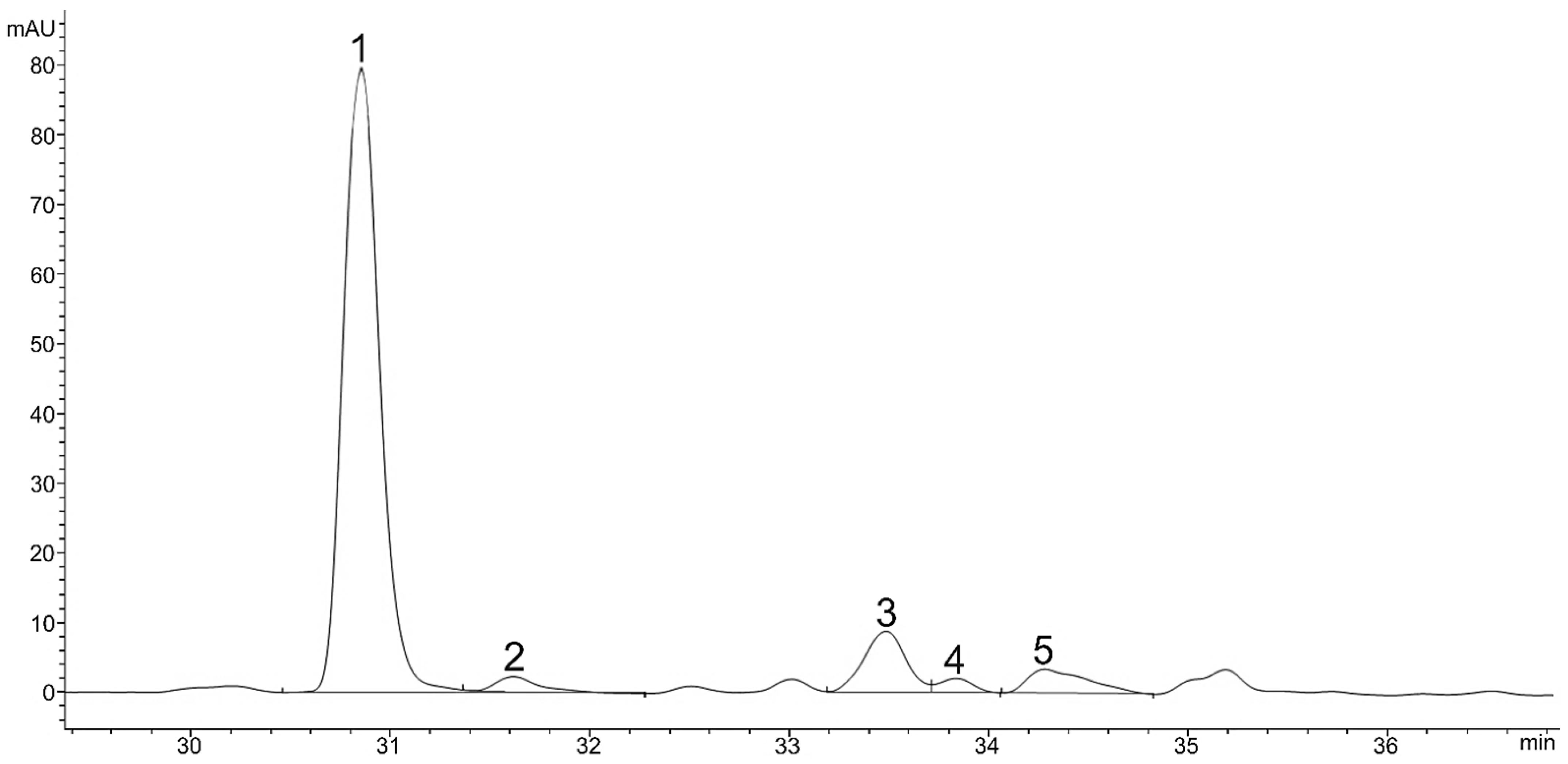
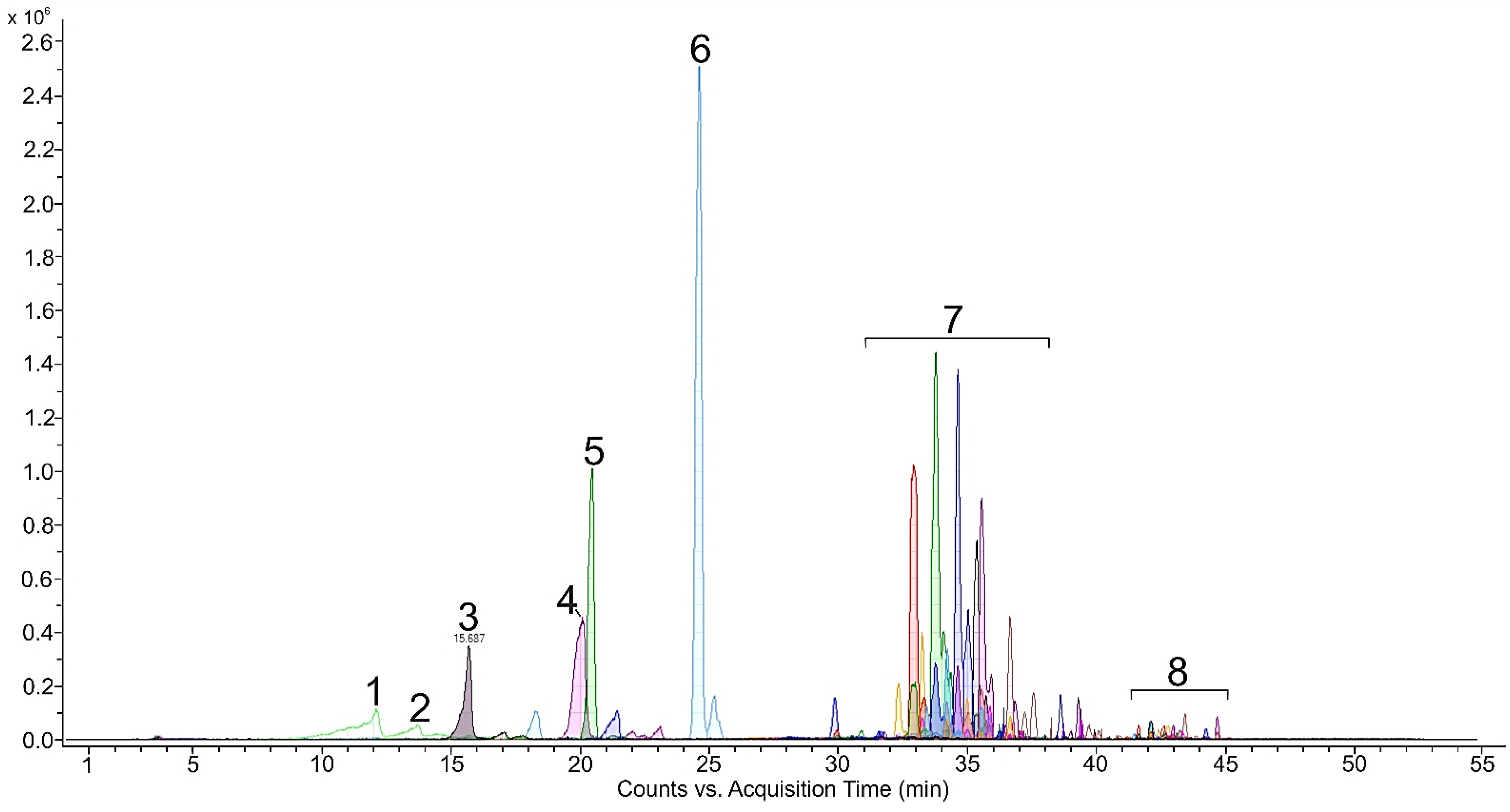
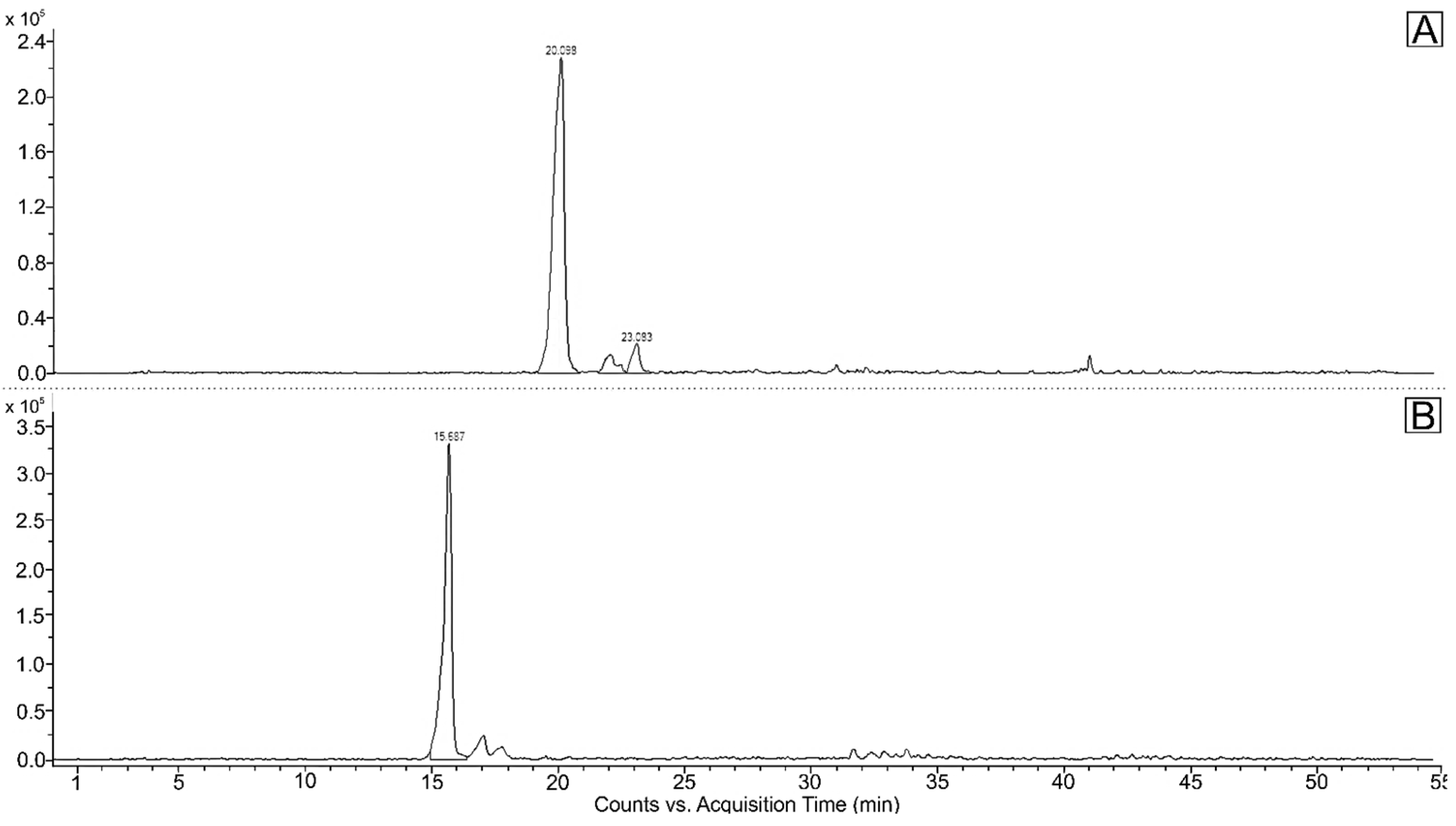
2.1. Carotenoids and Other Pigments in Green Vegetative Stage of Microalga H. pluvialis
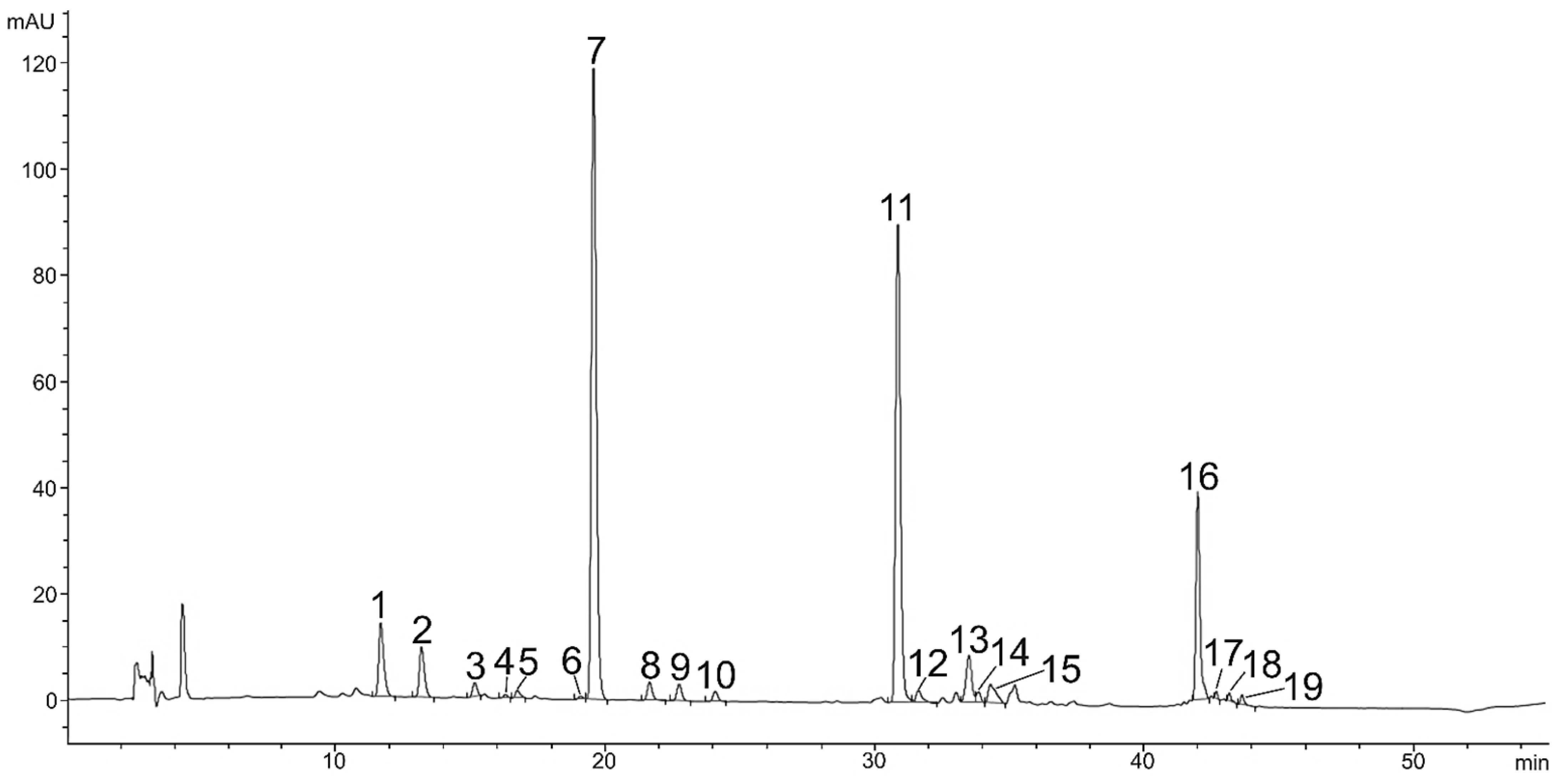
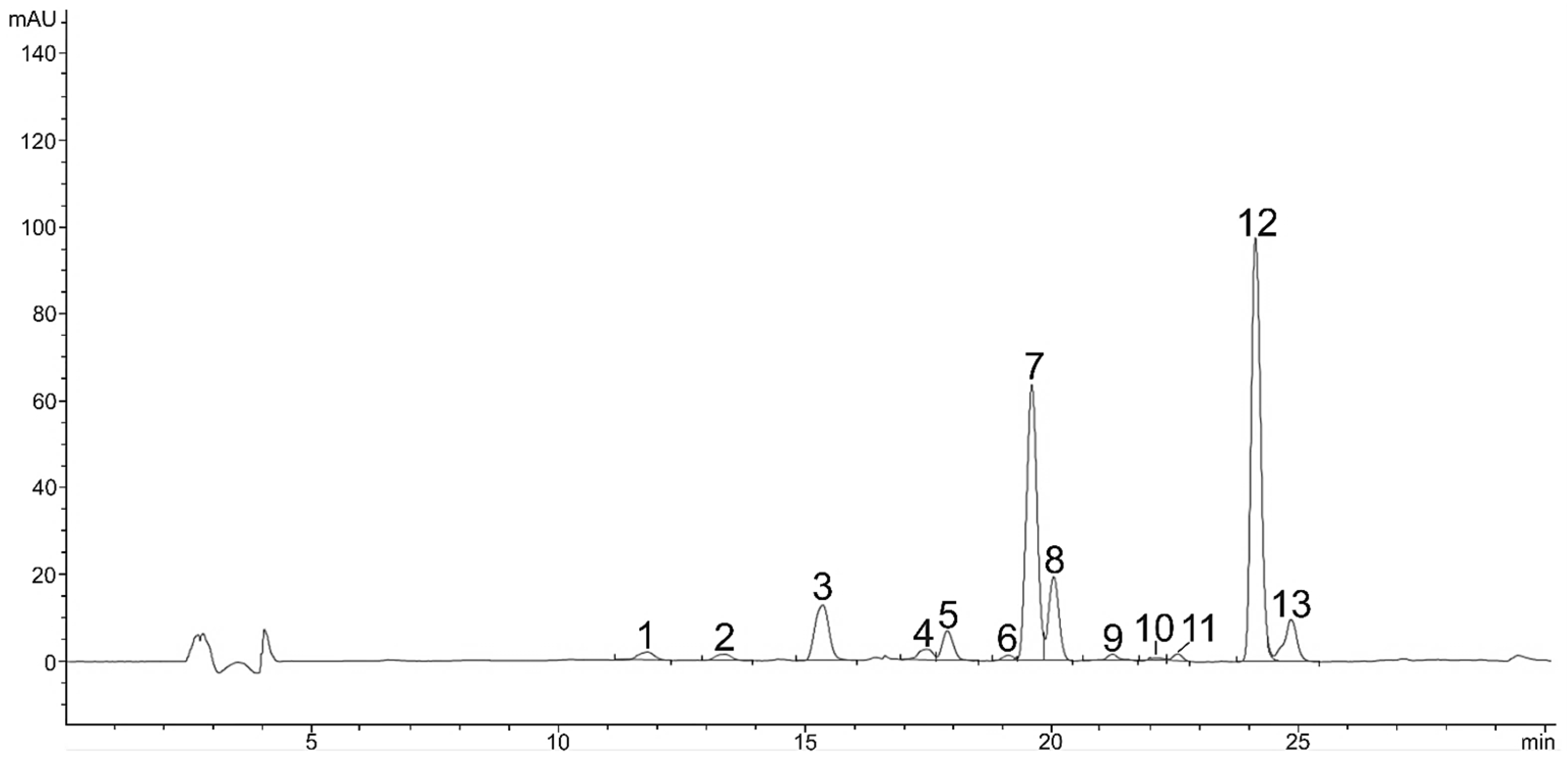
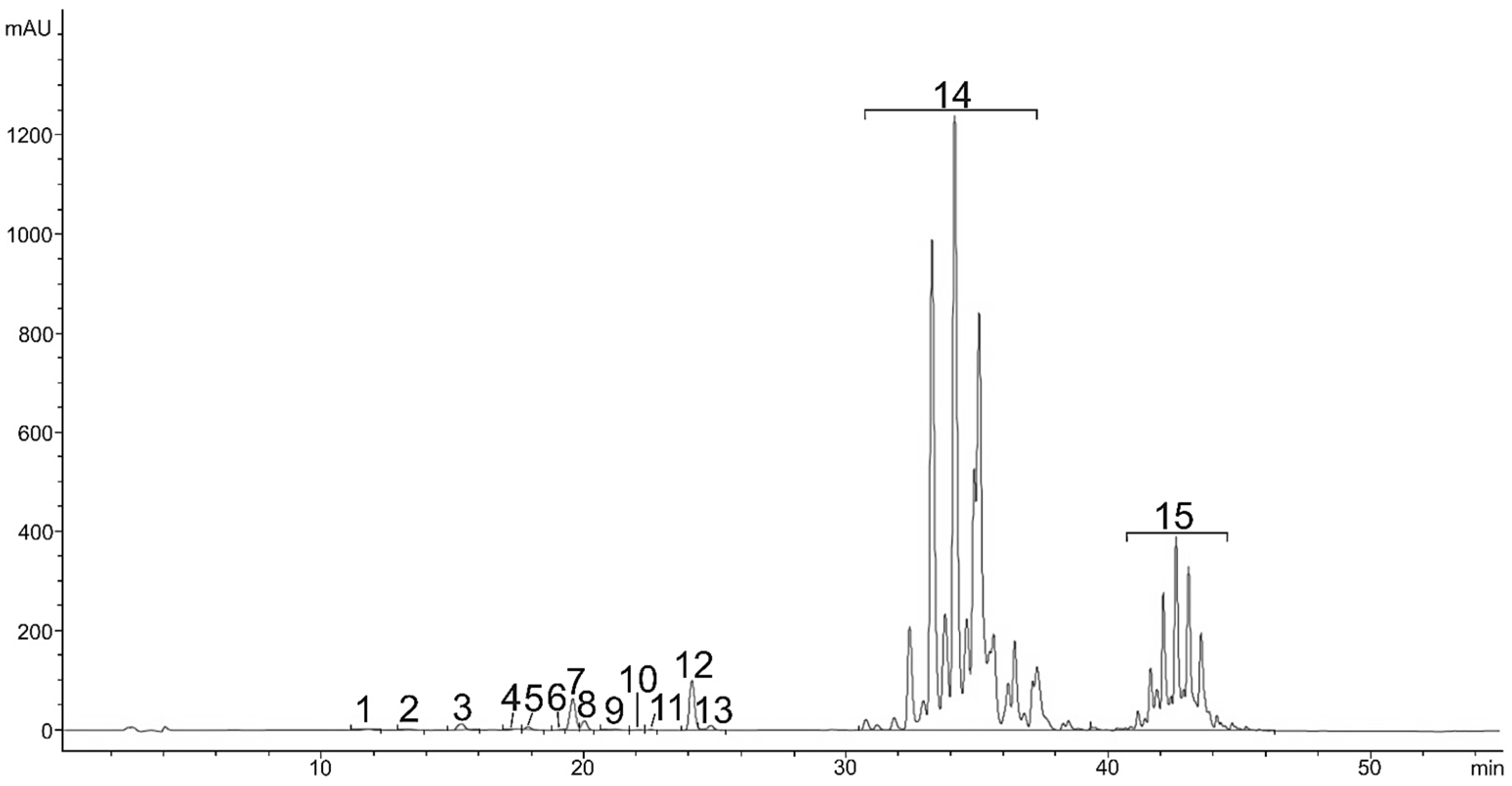
2.2. Carotenoids in Red Stage of Microalga H. pluvialis
3. Discussion
3.1. Green Vegetative Stage
3.2. Red Stage
4. Materials and Methods
4.1. Microalgal Culture and Growth Conditions
4.2. Dry Weight Determination
4.3. Carotenoid Extraction
4.4. HPLC-DAD and LC-QTOF-MS Analysis of Carotenoids
4.5. HPLC Standards
4.6. Method Validation
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal Prospects of Antioxidants: A Review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Zehiroglu, C.; Ozturk Sarikaya, S.B. The Importance of Antioxidants and Place in Today’s Scientific and Technological Studies. J. Food Sci. Technol. 2019, 56, 4757–4774. [Google Scholar] [CrossRef] [PubMed]
- El-Baz, F.K.; Salama, A.; Ali, S.I.; Elgohary, R. Haematococcus Pluvialis Carotenoids Enrich Fractions Ameliorate Liver Fibrosis Induced by Thioacetamide in Rats: Modulation of Metalloproteinase and Its Inhibitor. BioMed Res. Int. 2021, 2021, 6631415. [Google Scholar] [CrossRef] [PubMed]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.-F. Microalgal Carotenoids: A Review of Production, Current Markets, Regulations, and Future Direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and Chlorophylls as Antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef]
- Fang, L.; Zhang, J.; Fei, Z.; Wan, M. Chlorophyll as Key Indicator to Evaluate Astaxanthin Accumulation Ability of Haematococcus Pluvialis. Bioresour. Bioprocess. 2019, 6, 52. [Google Scholar] [CrossRef]
- Roca, M.; Pérez-Gálvez, A. Metabolomics of Chlorophylls and Carotenoids: Analytical Methods and Metabolome-Based Studies. Antioxidants 2021, 10, 1622. [Google Scholar] [CrossRef]
- Mulders, K.J.M.; Lamers, P.P.; Martens, D.E.; Wijffels, R.H. Phototrophic Pigment Production with Microalgae: Biological Constraints and Opportunities. J. Phycol. 2014, 50, 229–242. [Google Scholar] [CrossRef]
- Gong, M.; Bassi, A. Carotenoids from Microalgae: A Review of Recent Developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef]
- Takaichi, S. Carotenoids in Algae: Distributions, Biosyntheses and Functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef]
- Hu, J.; Lu, W.; Lv, M.; Wang, Y.; Ding, R.; Wang, L. Extraction and Purification of Astaxanthin from Shrimp Shells and the Effects of Different Treatments on Its Content. Rev. Bras. Farmacogn. 2019, 29, 24–29. [Google Scholar] [CrossRef]
- Rammuni, M.N.; Ariyadasa, T.U.; Nimarshana, P.H.V.; Attalage, R.A. Comparative Assessment on the Extraction of Carotenoids from Microalgal Sources: Astaxanthin from H. Pluvialis and β-Carotene from D. Salina. Food Chem. 2019, 277, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Casella, P.; Iovine, A.; Mehariya, S.; Marino, T.; Musmarra, D.; Molino, A. Smart Method for Carotenoids Characterization in Haematococcus Pluvialis Red Phase and Evaluation of Astaxanthin Thermal Stability. Antioxidants 2020, 9, 422. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-Producing Green Microalga Haematococcus Pluvialis: From Single Cell to High Value Commercial Products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [Green Version]
- Panis, G.; Carreon, J.R. Commercial Astaxanthin Production Derived by Green Alga Haematococcus Pluvialis: A Microalgae Process Model and a Techno-Economic Assessment All through Production Line. Algal Res. 2016, 18, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and Other Nutrients from Haematococcus Pluvialis—Multifunctional Applications. Mar. Drugs 2020, 18, 459. [Google Scholar] [CrossRef]
- Li, J.; Zhu, D.; Niu, J.; Shen, S.; Wang, G. An Economic Assessment of Astaxanthin Production by Large Scale Cultivation of Haematococcus Pluvialis. Biotechnol. Adv. 2011, 29, 568–574. [Google Scholar] [CrossRef]
- Kobayashi, M. Astaxanthin Biosynthesis Enhanced by Reactive Oxygen Species in the Green AlgaHaematococcus Pluvialis. Biotechnol. Bioprocess Eng. 2003, 8, 322–330. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M.; Godoy, H.T.; Amaya-Farfan, J. Updated Brazilian Database on Food Carotenoids: Factors Affecting Carotenoid Composition. J. Food Compos. Anal. 2008, 21, 445–463. [Google Scholar] [CrossRef]
- Gallego, R.; Arena, K.; Dugo, P.; Mondello, L.; Ibáñez, E.; Herrero, M. Application of Compressed Fluid–Based Extraction and Purification Procedures to Obtain Astaxanthin-Enriched Extracts from Haematococcus Pluvialis and Characterization by Comprehensive Two-Dimensional Liquid Chromatography Coupled to Mass Spectrometry. Anal. Bioanal. Chem. 2020, 412, 589–599. [Google Scholar] [CrossRef]
- Butler, T.; McDougall, G.; Campbell, R.; Stanley, M.; Day, J. Media Screening for Obtaining Haematococcus Pluvialis Red Motile Macrozooids Rich in Astaxanthin and Fatty Acids. Biology 2017, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.H.; Kim, J.M.; Kim, S.; Yoon, M.J.; Park, K.S. Chemical Transformation of Astaxanthin from Haematococcus Pluvialis Improves Its Antioxidative and Anti-Inflammatory Activities. ACS Omega 2020, 5, 19120–19130. [Google Scholar] [CrossRef] [PubMed]
- Fábryová, T.; Tůmová, L.; da Silva, D.C.; Pereira, D.M.; Andrade, P.B.; Valentão, P.; Hrouzek, P.; Kopecký, J.; Cheel, J. Isolation of Astaxanthin Monoesters from the Microalgae Haematococcus Pluvialis by High Performance Countercurrent Chromatography (HPCCC) Combined with High Performance Liquid Chromatography (HPLC). Algal Res. 2020, 49, 101947. [Google Scholar] [CrossRef]
- Miao, F.; Lu, D.; Li, Y.; Zeng, M. Characterization of Astaxanthin Esters in Haematococcus Pluvialis by Liquid Chromatography–Atmospheric Pressure Chemical Ionization Mass Spectrometry. Anal. Biochem. 2006, 352, 176–181. [Google Scholar] [CrossRef]
- Ranga, R.; Sarada, A.R.; Baskaran, V.; Ravishankar, G.A. Identification of Carotenoids from Green Alga Haematococcus Pluvialis by HPLC and LC-MS (APCI) and Their Antioxidant Properties. J. Microbiol. Biotechnol. 2009, 19, 1333–1341. [Google Scholar]
- Todorović, B.; Grujić, V.J.; Krajnc, A.U.; Kranvogl, R.; Ambrožič-Dolinšek, J. Identification and Content of Astaxanthin and Its Esters from Microalgae Haematococcus Pluvialis by HPLC-DAD and LC-QTOF-MS after Extraction with Various Solvents. Plants 2021, 10, 2413. [Google Scholar] [CrossRef]
- Saha, S.K.; Moane, S.; Murray, P. Effect of Macro- and Micro-Nutrient Limitation on Superoxide Dismutase Activities and Carotenoid Levels in Microalga Dunaliella Salina CCAP 19/18. Bioresour. Technol. 2013, 147, 23–28. [Google Scholar] [CrossRef]
- Zhou, Q.; Xu, J.; Yang, L.; Gu, C.; Xue, C. Thermal stability and oral absorbability of astaxanthin esters from Haematococcus pluvialis in Balb/c mice. J. Sci. Food Agric. 2019, 99, 3662–3671. [Google Scholar] [CrossRef]
- Bauer, A.; Minceva, M. Direct Extraction of Astaxanthin from the Microalgae Haematococcus Pluvialis Using Liquid–Liquid Chromatography. RSC Adv. 2019, 9, 22779–22789. [Google Scholar] [CrossRef] [Green Version]
- Gebregziabher, B.S.; Zhang, S.; Qi, J.; Azam, M.; Ghosh, S.; Feng, Y.; Huai, Y.; Li, J.; Li, B.; Sun, J. Simultaneous Determination of Carotenoids and Chlorophylls by the HPLC-UV-VIS Method in Soybean Seeds. Agronomy 2021, 11, 758. [Google Scholar] [CrossRef]
- Mariutti, L.R.B.; Mercadante, A.Z. Carotenoid Esters Analysis and Occurrence: What Do We Know so Far? Arch. Biochem. Biophys. 2018, 648, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Metličar, V.; Kranjc, K.; Albreht, A. Utilization of Plant-Based Wastes for a Sustainable Preparation of Xanthophyll Esters via Acid Anhydrides Using β-Pinene as a Bio-Derived Solvent. ACS Sustain. Chem. Eng. 2021, 9, 10651–10661. [Google Scholar] [CrossRef]
- Jaime, L.; Rodríguez-Meizoso, I.; Cifuentes, A.; Santoyo, S.; Suarez, S.; Ibáñez, E.; Señorans, F.J. Pressurized Liquids as an Alternative Process to Antioxidant Carotenoids’ Extraction from Haematococcus Pluvialis Microalgae. LWT-Food Sci. Technol. 2010, 43, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Renstrøm, B.; Liaaen-Jensen, S. Fatty Acid Composition of Some Esterified Carotenols. Comp. Biochem. Physiol. Part B Comp. Biochem. 1981, 69, 625–627. [Google Scholar] [CrossRef]
- Harker, M.; Tsavalos, A.J.; Young, A.J. Autotrophic Growth and Carotenoid Production of Haematococcus Pluvialis in a 30 Liter Air-Lift Photobioreactor. J. Ferment. Bioeng. 1996, 82, 113–118. [Google Scholar] [CrossRef]
- Grewe, C.B.; Griehl, C. 8 The Carotenoid Astaxanthin from Haematococcus Pluvialis. In Microalgal Biotechnology: Integration and Economy; Posten, C., Walter, C., Eds.; Walter de Gruyter: Berlin, Germany, 2012; pp. 129–144. ISBN 978-3-11-029827-7. [Google Scholar]
- Fu, W.; Magnúsdóttir, M.; Brynjólfson, S.; Palsson, B.Ø.; Paglia, G. UPLC-UV-MSE Analysis for Quantification and Identification of Major Carotenoid and Chlorophyll Species in Algae. Anal. Bioanal. Chem. 2012, 404, 3145–3154. [Google Scholar] [CrossRef]
- Sluiman, H. Phycology; Lee, R.E., Ed.; US160 (Hardback); Cambridge University Press: Cambridge, UK, 2008; 560p, ISBN 978 0 521 68277 0. [Google Scholar]
- Grewe, C.; Griehl, C. Time- and Media-Dependent Secondary Carotenoid Accumulation in Haematococcus Pluvialis. Biotechnol. J. 2008, 3, 1232–1244. [Google Scholar] [CrossRef]
- Dragoş, N.; Bercea, V.; Bica, A.; Drugă, B.; Nicoară, A.; Coman, C. Astaxanthin Production from a New Strain of Haematococcus Pluvialis Grown in Batch Culture. Ann. Rom. Soc. Cell Biol. 2010, 15, 353–361. [Google Scholar]
- Castillo, A.; Pereira, S.; Otero, A.; Fiol, S.; Garcia-Jares, C.; Lores, M. Matrix Solid-Phase Dispersion as a Greener Alternative to Obtain Bioactive Extracts from Haematococcus Pluvialis. Characterization by UHPLC-QToF. RSC Adv. 2020, 10, 27995–28006. [Google Scholar] [CrossRef]
- Willey, N. Environmental Plant Physiology; Garland Science: New York, NY, USA, 2018; ISBN 1-317-20623-1. [Google Scholar]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis: Second Edition; Princeton University Press: Princeton, NJ, USA, 2007; ISBN 978-1-4008-4972-7. [Google Scholar]
- Gu, W.; Xie, X.; Gao, S.; Zhou, W.; Pan, G.; Wang, G. Comparison of Different Cells of Haematococcus Pluvialis Reveals an Extensive Acclimation Mechanism during Its Aging Process: From a Perspective of Photosynthesis. PLoS ONE 2013, 8, e67028. [Google Scholar] [CrossRef] [PubMed]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. (Eds.) Carotenoids; Birkhäuser Basel: Basel, Switzerland, 2008; ISBN 978-3-7643-7498-3. [Google Scholar]
- Jayaraj, J.; Devlin, R.; Punja, Z. Metabolic Engineering of Novel Ketocarotenoid Production in Carrot Plants. Transgenic Res. 2008, 17, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gálvez, A.; Mínguez-Mosquera, M.I. Esterification of Xanthophylls and Its Effect on Chemical Behavior and Bioavailability of Carotenoids in the Human. Nutr. Res. 2005, 25, 631–640. [Google Scholar] [CrossRef]
- Sarada, R.; Vidhyavathi, R.; Usha, D.; Ravishankar, G.A. An Efficient Method for Extraction of Astaxanthin from Green Alga Haematococcus Pluvialis. J. Agric. Food Chem. 2006, 54, 7585–7588. [Google Scholar] [CrossRef]
- Bold, H.C. The Morphology of Chlamydomonas Chlamydogama, Sp. Nov. Bull. Torrey Bot. Club 1949, 76, 101. [Google Scholar] [CrossRef]
- Bischoff, K.B. Accuracy of the Pseudo Steady State Approximation for Moving Boundary Diffusion Problems. Chem. Eng. Sci. 1963, 18, 711–713. [Google Scholar] [CrossRef]
- Ördög, V.; Stirk, W.A.; Bálint, P.; van Staden, J.; Lovász, C. Changes in Lipid, Protein and Pigment Concentrations in Nitrogen-Stressed Chlorella Minutissima Cultures. J. Appl. Phycol. 2012, 24, 907–914. [Google Scholar] [CrossRef]
- Hrvolová, B.; Martínez-Huélamo, M.; Colmán-Martínez, M.; Hurtado-Barroso, S.; Lamuela-Raventós, R.; Kalina, J. Development of an Advanced HPLC–MS/MS Method for the Determination of Carotenoids and Fat-Soluble Vitamins in Human Plasma. Int. J. Mol. Sci. 2016, 17, 1719. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | Content (mg g−1 DW) | Rt (min) HPLC-DAD | m/z | Adduct |
|---|---|---|---|---|---|
| 1 | Neoxanthin | 0.44 ± 0.04 | 11.6 | 601.4251 | [M+H]+ |
| 2 | Violaxanthin | 0.57 ± 0.07 | 13.1 | 601.4251 | [M+H]+ |
| 3 | Astaxanthin | 0.06 ± 0.01 | 15.1 | 597.3938 | [M+H]+ |
| 4 | Adonixanthin | 0.17 ± 0.00 | 16.3 | 583.4362 | [M+H]+ |
| 5 | Antheraxanthin | 0.04 ± 0.00 | 16.7 | 585.4302 | [M+H]+ |
| 6 | Zeaxanthin | 0.01 ± 0.00 | 19.1 | 569.4353 | [M+H]+ |
| 7 | Lutein | 1.12 ± 0.09 | 19.5 | 569.4353 | [M+H]+ |
| 8 | Lutein isomer 1 | 0.02 ± 0.01 | 21.6 | 569.4353 | [M+H]+ |
| 9 | Lutein isomer 2 | 0.04 ± 0.00 | 22.7 | 569.4353 | [M+H]+ |
| 10 | Lutein isomer 3 | 0.02 ± 0.00 | 24.1 | 569.4353 | [M+H]+ |
| 11 | Chlorophyll b | / | 30.9 | 9075.219 | [M+H]+ |
| 12 | Chlorophyll b derivate | / | 31.6 | 9075.219 | [M+H]+ |
| 13 | Chlorophyll a | 4.58 ± 0.56 | 33.5 | 8935.426 | [M+H]+ |
| 14 | Chlorophyll a derivate | 0.66 ± 0.17 | 33.7 | 895.5219 | [M+H]+ |
| 15 | Echinenone | 0.06 ± 0.01 | 34.2 | 551.4247 | [M+H]+ |
| 16 | β-Carotene | 0.89 ± 0.04 | 42.0 | 537.4455 | [M+H]+ |
| No. | Compound Name | Content (mg g−1 DW) | Rt (min) HPLC-DAD | m/z | Adduct | Product Ion (M-FA) |
|---|---|---|---|---|---|---|
| 1 | Neoxanthin | 0.12 ± 0.03 | 11.8 | 601.4251 | [M+H]+ | / |
| 2 | Violaxanthin | 0.18 ± 0.05 | 13.3 | 601.4251 | [M+H]+ | / |
| 3 | Astaxanthin | 0.23 ± 0.04 | 15.1 | 597.3938 | [M+H]+ | / |
| 4 | Adonixanthin | 0.05 ± 0.01 | 16.6 | 583.4362 | [M+H]+ | / |
| 5 | Zeaxanthin | 0.02 ± 0.00 | 19.1 | 569.4353 | [M+H]+ | / |
| 6 | Lutein | 0.55 ± 0.14 | 19.5 | 569.4353 | [M+H]+ | / |
| 7 | Adonirubin | 0.18 ± 0.03 | 20.0 | 581.3989 | [M+H]+ | / |
| 8 | Lutein isomer | 0.18 ± 0.01 | 21.2 | 569.4353 | [M+H]+ | / |
| 9 | Lutein isomer | LOQ | 21.0 | 569.4353 | [M+H]+ | / |
| 10 | Lutein isomer | LOQ | 22.5 | 569.4353 | [M+H]+ | / |
| 11 | Canthaxanthin | 0.80 ± 0.17 | 24.1 | 565.4040 | [M+H]+ | / |
| 12 | Cis-Canthaxanthin | 0.12 ± 0.03 | 24.8 | 565.4040 | [M+H]+ | / |
| 13 | M+H-C16:2 | 1.69 ± 0.07 | / | 831.5928 | [M+H]+ | 579.3840 |
| 14 | M+H-C16:1 | 7.69 ± 0.29 | / | 833.6084 | [M+H]+ | 579.3840 |
| 15 | M+H-C16:0 | 8.61 ± 0.38 | / | 835.6241 | [M+H]+ | 579.3840 |
| 16 | M+H-C18:4 | 0.31 ± 0.01 | / | 855.5928 | [M+H]+ | 579.3840 |
| 17 | M+H-C18:3 | 1.69 ± 0.08 | / | 857.6084 | [M+H]+ | 579.3840 |
| 18 | M+H-C18:2 | 7.04 ± 0.06 | / | 859.6241 | [M+H]+ | 579.3840 |
| 19 | M+H-C20:2 | 1.57 ± 0.36 | 883.6241 | [M+H]+ | 579.3840 | |
| 20 | M+H-C18:1 | 0.11 ± 0.00 | / | 861.6397 | [M+H]+ | 579.3840 |
| 21 | M+H-C18:0 | 4.91 ± 0.24 | / | 863.6554 | [M+H]+ | 579.3840 |
| 22 | M+H-C20:2 | 2.27 ± 0.12 | / | 883.6241 | [M+H]+ | 579.3840 |
| 23 | M+H-C20:1 | 1.41 ± 0.15 | / | 885.6397 | [M+H]+ | 579.3840 |
| 24 | M+H-C20:0 | 0.56 ± 0.03 | / | 887.6554 | [M+H]+ | 579.3840 |
| 25 | M+H-C18:4/C18:4 | 0.75 ± 0.03 | / | 1113.7906 | [M+H]+ | 860.6783 |
| 26 | M+H-C18:4/C18:3 | 1.39 ± 0.05 | / | 1115.8062 | [M+H]+ | 862.6959 |
| 27 | M+H-C18:3/C18:3 | 0.90 ± 0.12 | / | 1117.8219 | [M+H]+ | 870.7575 |
| 28 | M+H-C18:2/C18:3 | 0.19 ± 0.00 | / | 1119.8375 | [M+H]+ | 872.7743 |
| 39 | M+H-C18:1/C18:3 | 2.24 ± 0.09 | / | 1121.8532 | [M+H]+ | 874.7896 |
| 30 | M+H-C18:1/C18:1 | 0.44 ± 0.10 | / | 1125.8845 | [M+H]+ | 888.7703 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grujić, V.J.; Todorović, B.; Kranvogl, R.; Ciringer, T.; Ambrožič-Dolinšek, J. Diversity and Content of Carotenoids and Other Pigments in the Transition from the Green to the Red Stage of Haematococcus pluvialis Microalgae Identified by HPLC-DAD and LC-QTOF-MS. Plants 2022, 11, 1026. https://doi.org/10.3390/plants11081026
Grujić VJ, Todorović B, Kranvogl R, Ciringer T, Ambrožič-Dolinšek J. Diversity and Content of Carotenoids and Other Pigments in the Transition from the Green to the Red Stage of Haematococcus pluvialis Microalgae Identified by HPLC-DAD and LC-QTOF-MS. Plants. 2022; 11(8):1026. https://doi.org/10.3390/plants11081026
Chicago/Turabian StyleGrujić, Veno Jaša, Biljana Todorović, Roman Kranvogl, Terezija Ciringer, and Jana Ambrožič-Dolinšek. 2022. "Diversity and Content of Carotenoids and Other Pigments in the Transition from the Green to the Red Stage of Haematococcus pluvialis Microalgae Identified by HPLC-DAD and LC-QTOF-MS" Plants 11, no. 8: 1026. https://doi.org/10.3390/plants11081026