
High Temperature Alters Anthocyanin Concentration and Composition in Grape Berries of Malbec, Merlot, and Pinot Noir in a Cultivar-Dependent Manner


Abstract
:1. Introduction
2. Results
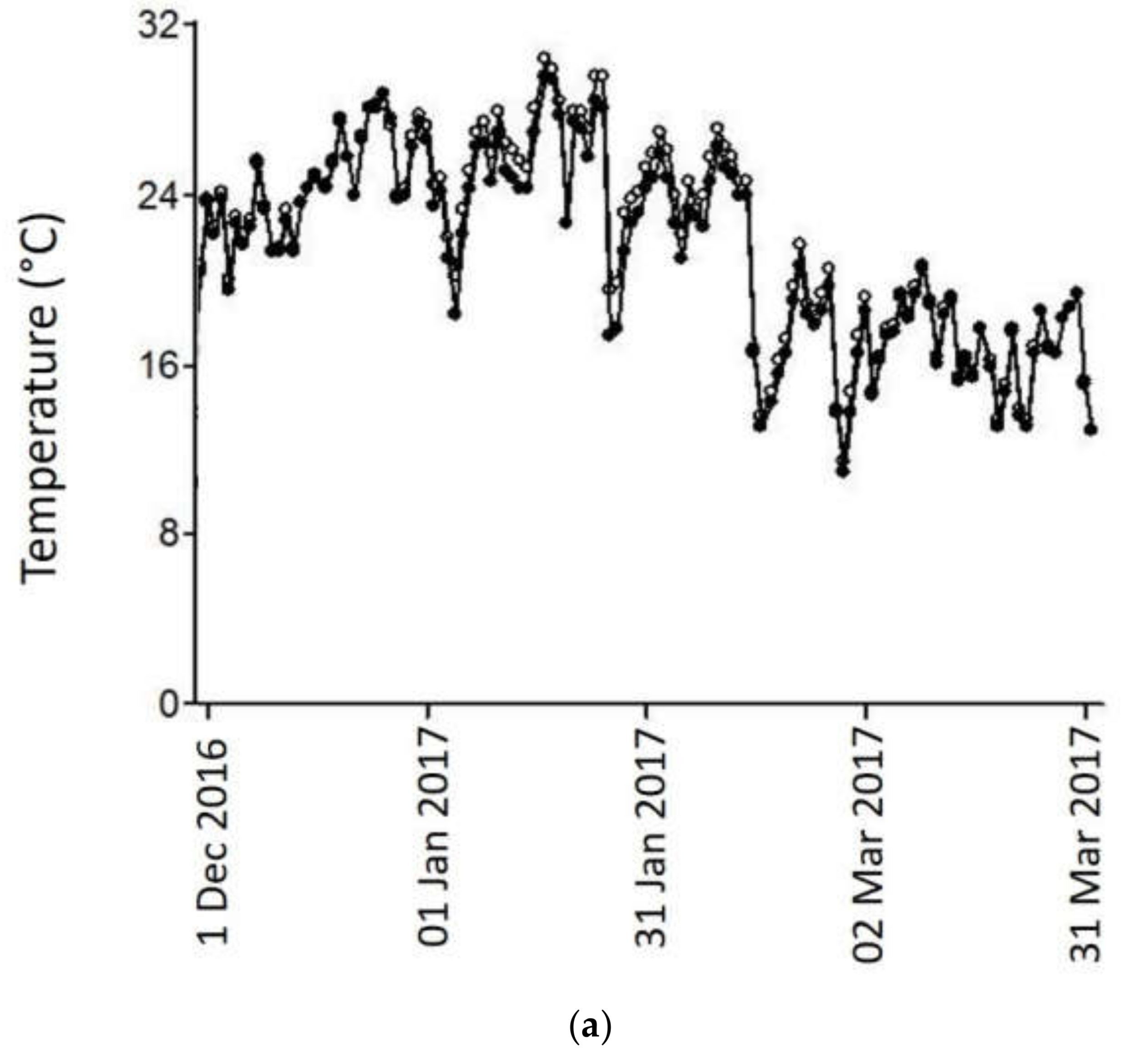
2.1. Temperature Treatments, Berry Soluble Solids and pH
2.2. Anthocyanin Concentration and Composition
3. Discussion
3.1. Temperature Regimes
3.2. Anthocyanin Concentration and Composition in Berries under High Temperature
4. Materials and Methods
4.1. Plant Materials, Temperature Conditions, and Experimental Design
4.2. Phenolics Extraction and HPLC Analysis of Anthocyanins and Resveratrol
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Biasi, R.; Brunori, E.; Ferrara, C.; Salvati, L. Assessing Impacts of Climate Change on Phenology and Quality Traits of Vitis vinifera L.: The Contribution of Local Knowledge. Plants 2019, 8, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-González, M.; Rodríguez-Rajo, F.J.; Escuredo, O.; Aira, M.J. Influence of thermal requirement in the aerobiological and phenological behaviour of two grapevine varieties. Aerobiologia 2013, 29, 523–535. [Google Scholar] [CrossRef]
- Dalla Marta, A.; Grifoni, D.; Mancini, M.; Storchi, P.; Zipoli, G.; Orlandini, S. Analysis of the relationships between climate variability and grapevine phenology in the Nobile di Montepulciano wine production area. J. Agric. Sci. 2010, 148, 657–666. [Google Scholar] [CrossRef]
- Petrie, P.R.; Sadras, V.O. Advancement of grapevine maturity in Australia between 1993 and 2006: Putative causes, magnitude of trends and viticulture consequences. Aust. J. Grape Wine Res. 2008, 14, 33–45. [Google Scholar] [CrossRef]
- Bonfante, A.; Monaco, E.; Langella, G.; Mercogliano, P.; Bucchignani, E.; Manna, P.; Terribile, F. A dynamic viticultural zoning to explore the resilience of terroir concept under climate change. Sci. Total Environ. 2018, 624, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Perrino, E.V.; Calabrese, G. Vascular flora of vineyards in the DOC area “Gioia del Colle” (Apulia, Southern Italy): Preliminary data. Nat. Croat. 2018, 27, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Tonietto, J.; Carbonneau, A. A multicriteria climatic classification system for grape-growing regions worldwide. Agric. For. Meteorol. 2004, 124, 81–97. [Google Scholar] [CrossRef] [Green Version]
- INV, Instituto Nacional de Vitivinicultura. Informe Anual de Superficie 2020; INV: Mendoza, Argentina, 2020; Available online: www.inv.gob.ar (accessed on 21 November 2021).
- Cabré, M.F.; Quénol, H.; Nuñez, M. Regional climate change scenarios applied to viticultural zoning in Mendoza. Argentina. Int. J. Biometeorol. 2016, 60, 1325–1340. [Google Scholar] [CrossRef]
- OIV. International Organization of Vine and Wine; OIV Press: Paris, France, 2022; Available online: www.OIV.org/es/noticias (accessed on 11 March 2022).
- OIV. International Organization of Vine and Wine; OIV Press: Paris, France, 2015; Available online: https://www.oiv.int/public/medias/5882/grapevine-varieties-area-by-country-2015 (accessed on 11 March 2022).
- Fanzone, M.; Zamora, F.; Jofré, V.; Assof, M.; Gómez-Cordovés, C.; Peña-Neira, Á. Phenolic characterisation of red wines from different grape varieties cultivated in Mendoza province (Argentina). J. Sci. Food Agric. 2012, 92, 704–718. [Google Scholar] [CrossRef]
- Kliewer, W.M.; Torres, R.E. Effect of controlled day and night temperatures on grape coloration. Am. J. Enol. Vitic. 1972, 23, 71–77. [Google Scholar]
- Keller, M.; Mills, L.J.; Wample, R.L.; Spayd, S.E. Cluster thinning effects on three deficit-irrigated Vitis vinifera L. Am. J. Enol. Vitic. 2005, 56, 91–103. [Google Scholar]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.L. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv Merlot Berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Tarara, J.M.; Lee, J.; Spayd, S.E.; Scagel, C.F. Berry temperature and solar radiation alter acylation, proportion, and concentration of anthocyanin in merlot grapes. Am. J. Enol. Vitic. 2008, 59, 235–247. [Google Scholar]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Ali, H.M.; Almagribi, W.; Al-Rashidi, M.N. Antiradical and reductant activities of anthocyanidins and anthocyanins, structure-activity relationship and synthesis. Food Chem. 2016, 1, 1275–1282. [Google Scholar] [CrossRef]
- Sadras, V.O.; Petrie, P.R. Climate shifts in south-eastern Australia: Early maturity of Chardonnay, Shiraz and Cabernet Sauvignon is associated with early onset rather than faster ripening. Aust. J. Grape Wine Res. 2011, 17, 199–205. [Google Scholar] [CrossRef]
- Mori, K.; Sugaya, S.; Gemma, H. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic. 2005, 105, 319–330. [Google Scholar] [CrossRef]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K. Loss of anthocyanins in red wine grape under high temperature. J. Exp. Bot. 2007, 58, 1935–1945. [Google Scholar] [CrossRef]
- Yamane, T.; Jeong, S.K.; Goto-Yamamoto, N.; Koshita, Y.; Kobayashi, S. Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am. J. Enol. Vitic. 2006, 57, 54–59. [Google Scholar]
- Van Leeuwen, C.; Destrac-Irvine, A. Modified grape composition under climate change conditions requires adaptations in the vineyard. OENO One 2017, 51, 147–154. [Google Scholar] [CrossRef]
- Rocchetti, G.; Ferrari, F.; Trevisan, M.; Bavaresco, L. Impact of climatic conditions on the resveratrol concentration in blend of Vitis vinifera L. cvs. Barbera and Croatina grape wines. Molecules 2021, 26, 401. [Google Scholar] [CrossRef]
- Bavaresco, L.; Lucini, L.; Busconi, M.; Flamini, R.; De Rosso, M. Wine resveratrol: From the ground up. Nutrients 2016, 8, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeandet, P.; Sbaghi, M.; Bessis, R.; Meunier, P. The potential relationship of stilbene (resveratrol) synthesis to anthocyanin content in grape berry skins. Vitis 1995, 34, 91–94. [Google Scholar]
- IPCC. Cambio climático: Informe de síntesis. Contribución de los Grupos de trabajo I, II y III al Cuarto Informe de evaluación del Grupo Intergubernamental de Expertos sobre el Cambio Climático. In Equipo de Redacción Principal; Pachauri, R.K., Reisinger, A., Eds.; IPCC: Ginebra, Switzerland, 2007; pp. 1–104. [Google Scholar]
- Laitinen, R.A.E.; Nikoloski, Z. Genetic basis of plasticity in plants. J. Exp. Bot. 2019, 70, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Perrino, E.V.; Wagensommer, R.P. Crop Wild Relatives (CWRs) Threatened and Endemic to Italy: Urgent Actions for Protection and Use. Biology 2022, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Boninsegna, J.R.; Villalba, R. Documento sobre la oferta hídrica en los oasis de riego de Mendoza y San Juan en escenarios de Cambio Climático. II–Los escenarios de Cambio Climático y el impacto en los caudales. In Informe a la Secretaría de Ambiente y Desarrollo Sustentable de la Nación; MYNCYT Press: Buenos Aires, Argentina, 2006; pp. 1–26. [Google Scholar]
- Masiokas, M.H.; Villalba, R.; Luckman, B.H.; Le Quesne, C.; Aravena, J.C. Snowpack variations in the central Andes of Argentina and Chile, 1951–2005: Large-scale atmospheric influences and implications for water resources in the region. J. Clim. 2006, 19, 6334–6352. [Google Scholar] [CrossRef]
- Duchêne, E. How can grapevine genetics contribute to the adaptation to climate change? OENO One 2016, 50, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Fanzone, M.; Peña-Neira, A.; Gil, M.; Jofré, V.; Assof, M.; Zamora, F. Impact of phenolic and polysaccharidic composition on commercial value of Argentinean Malbec and Cabernet Sauvignon wines. Food Res. Int. 2012, 45, 402–414. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Z.Z.; Yuan, L.; Gao, X.T.; Li, Q. Difference and characteristics of anthocyanin from Cabernet Sauvignon and Merlot cultivated at five regions in Xinjiang. Food Sci. Technol. 2020, 41, 72–80. [Google Scholar] [CrossRef]
- Van Buren, J.P.; Bertino, J.J.; Robinson, W.B. The Stability of Wine Anthocyanins on Exposure to Heat and Light. Am. J. Enol Vitic. 1968, 19, 147–154. [Google Scholar]
- Rinaldo, A.R.; Cavallini, E.; Jia, Y.; Moss, S.M.A.; McDavid, D.A.J.; Hooper, L.C.; Robinson, S.P.; Tornielli, G.B.; Zenoni, S.; Ford, C.M.; et al. A grapevine anthocyanin acyltransferase, transcriptionally regulated by vvmyba, can produce most acylated anthocyanins present in grape skins. Plant. Physiol. 2015, 169, 1897–1916. [Google Scholar] [CrossRef]
- Lecourieux, F.; Kappel, C.; Pieri, P.; Charon, J.; Pillet, J.; Hilbert, G.; Renaud, C.; Gomès, E.; Delrot, S.; Lecourieux, D. Dissecting the biochemical and transcriptomic effects of a locally applied heat treatment on developing cabernet sauvignon grape berries. Front. Plant. Sci. 2017, 8, 53. [Google Scholar] [CrossRef] [Green Version]
- Torres, N.; Hilbert, G.; Luquin, J.; Goicoechea, N.; Antolín, M.C. Flavonoid and amino acid profiling on Vitis vinifera L. cv tempranillo subjected to deficit irrigation under elevated temperatures. J. Food Compos. Anal. 2017, 62, 51–62. [Google Scholar] [CrossRef]
- Pastore, C.; Dal Santo, S.; Zenoni, S.; Movahed, N.; Allegro, G.; Valentini, G.; Filippetti, I.; Tornielli, G.B. Whole plant temperature manipulation affects flavonoid metabolism and the transcriptome of grapevine berries. Front. Plant. Sci. 2017, 8, 929. [Google Scholar] [CrossRef]
- Yan, Y.; Song, C.; Falginella, L.; Castellarin, S.D. Day temperature has a stronger effect than night temperature on anthocyanin and flavonol accumulation in “merlot” (Vitis vinifera L.) grapes during ripening. Front. Plant. Sci. 2020, 11, 1095. [Google Scholar] [CrossRef]
- Deis, L.; de Rosas, M.I.; Malovini, E.; Cavagnaro, M.; Cavagnaro, J.B. Climate change impact in Mendoza. Climate variation on the last 50 years. A view to grapevine physiology. Rev. Fac. Cienc. Agrar. 2015, 47, 67–92. [Google Scholar]
- Movahed, N.; Pastore, C.; Cellini, A.; Allegro, G.; Valentini, G.; Zenoni, S.; Cavallini, E.; D’Incà, E.; Tornielli, G.B.; Filippetti, I. The grapevine VviPrx31 peroxidase as a candidate gene involved in anthocyanin degradation in ripening berries under high temperature. J. Plant. Res. 2016, 129, 513–526. [Google Scholar] [CrossRef]
- De Rosas, I.; Deis, L.; Ponce, M.T.; Malovini, E.; Cavagnaro, J.B.; Cavagnaro, P.F. Loss of anthocyanins and modification of the anthocyanin profiles in grape berries of Malbec and Bonarda grown under high temperature conditions. Plant. Sci. 2017, 258, 137–145. [Google Scholar] [CrossRef]
- Sadras, V.O.; Moran, M.A. Elevated temperature decouples anthocyanins and sugars in berries of Shiraz and Cabernet Franc. Aust. J. Grape Wine Res. 2012, 18, 115–122. [Google Scholar] [CrossRef]
- Sadras, V.O.; Moran, M.A.; Bonada, M. Effects of elevated temperature in grapevine. I berry sensory traits. Aust. J. Grape Wine Res. 2013, 19, 95–106. [Google Scholar] [CrossRef]
- Fernandes de Oliveira, A.; Mercenaro, L.; Del Caro, A.; Pretti, L.; Nieddu, G. Distinctive anthocyanin accumulation responses to temperature and natural UV radiation of two field-grown (Vitis vinifera L.) cultivars. Molecules 2015, 20, 2061–2080. [Google Scholar] [CrossRef] [PubMed]
- Arrizabalaga, M.; Morales, F.; Oyarzun, M.; Delrot, S.; Gômes, E.; Irigoyen, J.J.; Hilbert, G.; Pascual, I. Tempranillo clones differ in the response of berry sugar and anthocyanin accumulation to elevated temperature. Plant. Sci. 2018, 267, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Jackman, R.L.; Smith, J.L. Anthocyanins and betalains. In Natural Food Colorants; Hendry, G.A.F., Houghton, J.D., Eds.; Springer: Boston, MA, USA, 1996; pp. 244–309. [Google Scholar]
- Hrazdina, G.; Parsons, G.F.; Mati1ck, L.R. Physiological and biochemical events during development and maturation of grape berries. Amer. J. Enol. Viticult. 1985, 35, 220–227. [Google Scholar]
- Revilla, E.; Ryan, J.M.; Martin-Ortega, G. Comparison of several procedures used for the extraction of anthocyanins from red grapes. J. Agric. Food Chem. 1998, 46, 4592–4597. [Google Scholar] [CrossRef]
- OIV. International organization of vine and wine. Method OIV-MA-AS315-11. In Compendium of International Methods of Wine and Must Analysis; OIV Press: Paris, France, 2014; pp. 1–13. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatment | Malbec | Merlot | Pinot Noir | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Veraison | Half Ripeness | Harvest | Veraison | Half Ripeness | Harvest | Veraison | Half Ripeness | Harvest | ||||||||||||
| DDs | SS § | pH | SS | pH | SS | pH | SS | pH | SS | pH | SS | pH | SS | pH | S | pH | SS | pH | ||
| 2017 | HT | 541 | 11.9 | 3.2 | 18.6 | 3.4 | 24.0 | 3.4 | 11.5 | 3.1 | 20.6 | 3.6 | 23.6 | 3.7 | 16.1 | 3.7 | 19.5 | 3.8 | 21.2 | 4.0 |
| C | 482 | 11.5 | 3.1 | 18.0 | 3.3 | 23.8 | 3.3 | 12.5 | 3.1 | 19.8 | 3.5 | 24.0 | 3.6 | 16.9 | 3.6 | 18.1 | 3.7 | 21.6 | 3.8 | |
| ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |||
| 2018 | HT | 706 | 14.7 | 3.0 | 17.6 | 3.3 | 24.9 | 3.9 | 13.8 | 3.2 | 18.0 | 3.5 | 22.3 | 3.8 | 12.8 | 3.1 | 16.4 | 3.6 | 21.8 | 4.0 |
| C | 612 | 15.3 | 3.0 | 17.0 | 3.3 | 24.1 | 3.8 | 13.6 | 3.1 | 18.1 | 3.4 | 23.5 | 3.8 | 13.1 | 3.2 | 15.7 | 3.4 | 20.3 | 3.8 | |
| ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |||
| Compound | Abbreviation | RT |
|---|---|---|
| delphinidin-3-glucoside | Df | 12.87 |
| cyanidin-3-glucoside | Cn | 15.33 |
| petunidin-3-glucoside | Pt | 17.11 |
| peonidin-3-glucoside | Po | 19.57 |
| malvidin-3-glucoside | Mv | 20.85 |
| peonidin-3-O-acetylglucoside | PoAc | 28.81 |
| malvidin-3-O-acetyl-glucoside | MvAc | 29.30 |
| peonidin-3-O-coumaroyl-glucoside | PoCu | 34.56 |
| malvidin-3-O-coumaroyl-glucoside | MvCu | 35.33 |
| Anthocyanin Pigment §/Year | MALBEC | |||||
|---|---|---|---|---|---|---|
| Veraison | Half Ripeness | Harvest | ||||
| HT | C | HT | C | HT | C | |
| Df | ||||||
| 2017 | 4.3 ± 0.3 | 4.5 ± 0.2 | 4.5 ± 0.3 | 6.1 ± 0.6 * | 4.3 ± 0.3 | 4.8 ± 0.4 |
| 2018 | 6.7 ± 0.6 | 7.5 ± 0.4 | 4.7 ± 0.4 | 5.4 ± 0.3 | 4.3 ± 0.1 | 4.5 ± 0.3 |
| Cn | ||||||
| 2017 | 1.0 ± 0.1 | 1.1 ± 0.1 | 1.2 ± 0.1 | 1.0 ± 0.2 | 1.1 ± 0.1 | 1.0 ± 0.1 |
| 2018 | 0.8 ± 0.1 | 1.1 ± 0.1 * | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.1 ± 0.01 | 0.1 ± 0.02 |
| Pt | ||||||
| 2017 | 8.4 ± 0.3 | 8.5 ± 0.3 | 9.5 ± 0.4 | 10.7 ± 0.7 | 8.2 ± 0.3 | 9.0 ± 0.6 |
| 2018 | 11.1 ± 0.7 | 11.9 ± 0.3 | 8.4 ± 0.6 | 9.4 ± 0.2 | 8.0 ± 0.1 | 8.3 ± 0.3 |
| Po | ||||||
| 2017 | 3.7 ± 0.4 | 3.6 ± 0.2 | 6.3 ± 0.6 * | 4.2 ± 0.1 | 6.7 ± 0.5 * | 4.7 ± 0.4 |
| 2018 | 4.4 ± 0.1 | 4.8 ± 0.3 | 2.0 ± 0.1 | 2.7 ± 0.3 | 0.3 ± 0.03 | 1.7 ± 0.8 |
| Mv | ||||||
| 2017 | 46.6 ± 0.8 | 46.5 ± 0.7 | 54.9 ± 1.1 | 55.2 ± 0.4 | 54.0 ± 0.9 | 55.3 ± 1.2 |
| 2018 | 54.3 ± 0.5 | 53.5 ± 0.8 | 50.7 ± 1.4 | 54.4 ± 0.9 * | 54.6 ± 0.3 | 55.6 ± 0.40 * |
| PoAc | ||||||
| 2017 | 0.3 ± 0.02 | 0.3 ± 0.02 | 0.2 ± 0.02 ** | 0.2 ± 0.02 | 0.1 ± 0.01 * | 0.1 ± 0.01 |
| 2018 | 0.2 ± 0.02 | 0.2 ± 0.02 | 0.2 ± 0.04 | 0.2 ± 0.02 | 0.1 ± 0.01 * | 0.1 ± 0.00 |
| MvAc | ||||||
| 2017 | 5.0 ± 0.2 | 5.4 ± 0.03 * | 3.0 ± 0.2 | 3.1 ± 0.4 | 1.4 ± 0.08 | 1.4 ± 0.06 |
| 2018 | 0.8 ± 0.02 | 1.1 ± 0.3 | 0.6 ± 0.1 * | 0.3 ± 0.1 | 0.1 ± 0.00 | 0.2 ± 0.01 |
| PoCu | ||||||
| 2017 | 2.8 ± 0.1 * | 2.5 ± 0.02 | 1.8 ± 0.2 * | 1.2 ± 0.04 | 2.2 ± 0.3 * | 1.5 ± 0.2 |
| 2018 | 2.0 ± 0.1 | 1.9 ± 0.2 | 1.5 ± 0.1 | 1.3 ± 0.01 | 1.6 ± 0.1 * | 1.3 ± 0.1 |
| MvCu | ||||||
| 2017 | 27.9 ± 1.7 | 27.7 ± 1.3 | 18.4 ± 1.1 | 18.2 ± 1.6 | 21.9 ± 1.0 | 22.2 ± 1.6 |
| 2018 | 19.8 ± 1.6 | 18.1 ± 0.8 | 31.5 ± 2.6 * | 26.0 ± 0.9 | 30.9 ± 0.4 * | 28.2 ± 0.7 |
| Total AA | ||||||
| 2017 | 36.0 | 35.9 | 23.6 | 22.7 | 25.6 | 25.2 |
| 2018 | 22.7 | 21.2 | 33.8 * | 27.7 | 32.7 * | 29.8 |
| Df | ||||||
| 2017 | 5.7 ± 0.4 | 5.5 ± 0.5 | 6.3 ± 0.2 | 8.5 ± 0.5 * | 7.8 ± 0.8 ** | 7.0 ± 0.5 |
| 2018 | 8.8 ± 1.0 | 13.1 ± 1.3 * | 7.0 ± 0.8 | 9.7 ± 1.3 | 6.6 ± 0.4 | 8.1 ± 0.3 * |
| Cn | ||||||
| 2017 | 2.8 ± 0.3 | 2.9 ± 0.4 | 3.5 ± 0.4 | 3.7 ± 0.1 | 5.7 ± 0.8 * | 3.3 ± 0.4 |
| 2018 | 2.2 ± 0.6 | 3.8 ± 1.0 * | 1.1 ± 0.2 | 2.0 ± 0.4 * | 1.2 ± 0.1 | 2.2 ± 0.3 * |
| Pt | ||||||
| 2017 | 9.1 ± 0.4 | 9.3 ± 0.4 | 11.0 ± 0.2 | 12.7 ± 0.5 * | 10.8 ± 0.6 | 10.7 ± 0.3 |
| 2018 | 12.5 ± 0.8 | 13.9 ± 0.8 ** | 10.5 ± 0.6 | 12.3 ± 0.9 | 9.5 ± 0.3 | 10.7 ± 0.3 * |
| Po | ||||||
| 2017 | 10.1 ± 0.8 | 11.6 ± 0.6 | 10.8 ± 1.2 | 9.7 ± 0.6 | 13.9 ± 0.8 * | 10.2 ± 0.9 |
| 2018 | 6.8 ± 1.1 | 8.9 ± 1.3 * | 4.6 ± 0.5 | 7.0 ± 0.8 * | 4.9 ± 0.4 | 7.4 ± 0.8 * |
| Mv | ||||||
| 2017 | 53.9 ± 0.8 | 52.8 ± 2.3 | 56.5 ± 1.0 * | 54.2 ± 0.8 | 50.4 ± 1.2 | 55.9 ± 1.3 * |
| 2018 | 59.3 ± 2.5 ** | 51.6 ± 3.2 | 62.8 ± 1.8 * | 57.6 ± 2.2 | 63.2 ± 0.9 ** | 58.5 ± 1.2 |
| PoAc | ||||||
| 2017 | 0.9 ± 0.1 | 1.0 ± 0.1 | 0.5 ± 0.04 | 0.5 ± 0.01 | 0.2 ± 0.03 | 0.2 ± 0.03 |
| 2018 | 0.1 ± 0.01 ** | 0.1 ± 0.01 | 0.2 ± 0.02 * | 0.1 ± 0.01 | 0.1 ± 0.00 ** | 0.07 ± 0.00 |
| MvAc | ||||||
| 2017 | 7.2 ± 0.2 | 6.7 ± 0.5 | 3.7 ± 0.5 | 3.9 ± 0.2 | 1.4 ± 0.2 | 1.9 ± 0.1 * |
| 2018 | 0.3 ± 0.05 * | 0.1 ± 0.02 | 0.2 ± 0.04 | 0.1 ± 0.01 | 0.4 ± 0.00 * | 0.3 ± 0.05 |
| PoCu | ||||||
| 2017 | 1.4 ± 0.1 | 1.3 ± 0.3 | 1.2 ± 0.1 * | 1.0 ± 0.1 | 2.1 ± 0.1 * | 1.7 ± 0.1 |
| 2018 | 1.2 ± 0.1 | 1.4 ± 0.1 ** | 1.3 ± 0.1 | 1.3 ± 0.1 | 1.3 ± 0.1 | 1.8 ± 0.2 * |
| MvCu | ||||||
| 2017 | 8.9 ± 0.6 | 9.0 ± 2.1 | 6.6 ± 0.1 * | 5.9 ± 0.2 | 7.7 ± 0.2 | 9.1 ± 0.4 * |
| 2018 | 8.9 ± 1.0 * | 7.1 ± 0.9 | 12.4 ± 0.8 * | 10.0 ± 1.1 | 12.7 ± 0.2* | 10.9 ± 0.4 |
| Total AA | ||||||
| 2017 | 18.3 | 18.0 | 12.0 | 11.3 | 11.4 | 13.0 |
| 2018 | 10.5 * | 8.7 | 14.0 * | 11.5 | 14.6 * | 13.1 |
| Df | ||||||
| 2017 | 0.6 ± 0.1 * | 0.4 ± 0.1 | 0.9 ± 0.02 | 0.8 ± 0.2 | 1.3 ± 0.1 * | 1.1 ± 0.03 |
| 2018 | 1.5 ± 0.4 | 3.0 ± 0.3 * | 2.0 ± 0.2 | 3.9 ± 0.1 ** | 3.2 ± 0.3 | 3.6 ± 0.3 |
| Cn | ||||||
| 2017 | 0.3 ± 0.05 | 0.2 ± 0.04 | 0.5 ± 0.01 | 0.5 ± 0.1 | 1.0 ± 0.1 * | 0.8 ± 0.1 |
| 2018 | 0.9 ± 0.2 | 1.0 ± 0.1 | 1.3 ± 0.1 * | 1.1 ± 0.04 | 1.8 ± 0.3 | 2.3 ± 0.01 * |
| Pt | ||||||
| 2017 | 4.0 ± 0.4 * | 3.0 ± 0.6 | 4.4 ± 0.1 | 3.8 ± 0.6 | 4.8 ± 0.1 * | 4.4 ± 0.1 |
| 2018 | 4.4 ± 0.7 | 6.9 ± 0.5 * | 5.2 ± 0.2 | 7.6 ± 0.2 ** | 6.1 ± 0.4 | 6.2 ± 0.4 |
| Po | ||||||
| 2017 | 9.2 ± 0.8 | 10.8 ± 0.3 | 12.2 ± 0.9 | 13.5 ± 1.3 | 17.6 ± 0.6 | 16.2 ± 1.5 |
| 2018 | 16.1 ± 3.0 | 12.1 ± 0.3 | 18.6 ± 1.7 | 15.6 ± 1.0 | 21.6 ± 3.1 | 23.7 ± 1.0 |
| Mv | ||||||
| 2017 | 85.9 ± 0.5 | 85.7 ± 0.8 | 82.0 ± 0.8 | 81.4 ± 1.8 | 75.4 ± 0.6 | 77.6 ± 1.7 |
| 2018 | 77.3 ± 2.8 | 77.2 ± 0.9 | 73.0 ± 1.4 | 72.0 ± 1.0 | 67.3 ± 3.0 | 64.3 ± 0.6 |
| PoAc | ||||||
| 2017 | ND | ND | ND | ND | ND | ND |
| 2018 | ND | ND | ND | ND | ND | ND |
| MvAc | ||||||
| 2017 | ND | ND | ND | ND | ND | ND |
| 2018 | ND | ND | ND | ND | ND | ND |
| PoCu | ||||||
| 2017 | ND | ND | ND | ND | ND | ND |
| 2018 | ND | ND | ND | ND | ND | ND |
| MvCu | ||||||
| 2017 | ND | ND | ND | ND | ND | ND |
| 2018 | ND | ND | ND | ND | ND | ND |
| Total AA | ||||||
| 2017 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| 2018 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Rosas, I.; Deis, L.; Baldo, Y.; Cavagnaro, J.B.; Cavagnaro, P.F. High Temperature Alters Anthocyanin Concentration and Composition in Grape Berries of Malbec, Merlot, and Pinot Noir in a Cultivar-Dependent Manner. Plants 2022, 11, 926. https://doi.org/10.3390/plants11070926
de Rosas I, Deis L, Baldo Y, Cavagnaro JB, Cavagnaro PF. High Temperature Alters Anthocyanin Concentration and Composition in Grape Berries of Malbec, Merlot, and Pinot Noir in a Cultivar-Dependent Manner. Plants. 2022; 11(7):926. https://doi.org/10.3390/plants11070926
Chicago/Turabian Stylede Rosas, Inés, Leonor Deis, Yésica Baldo, Juan B. Cavagnaro, and Pablo F. Cavagnaro. 2022. "High Temperature Alters Anthocyanin Concentration and Composition in Grape Berries of Malbec, Merlot, and Pinot Noir in a Cultivar-Dependent Manner" Plants 11, no. 7: 926. https://doi.org/10.3390/plants11070926