
Negative Effects of Rhizobacteria Association on Plant Recruitment of Generalist Predators

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
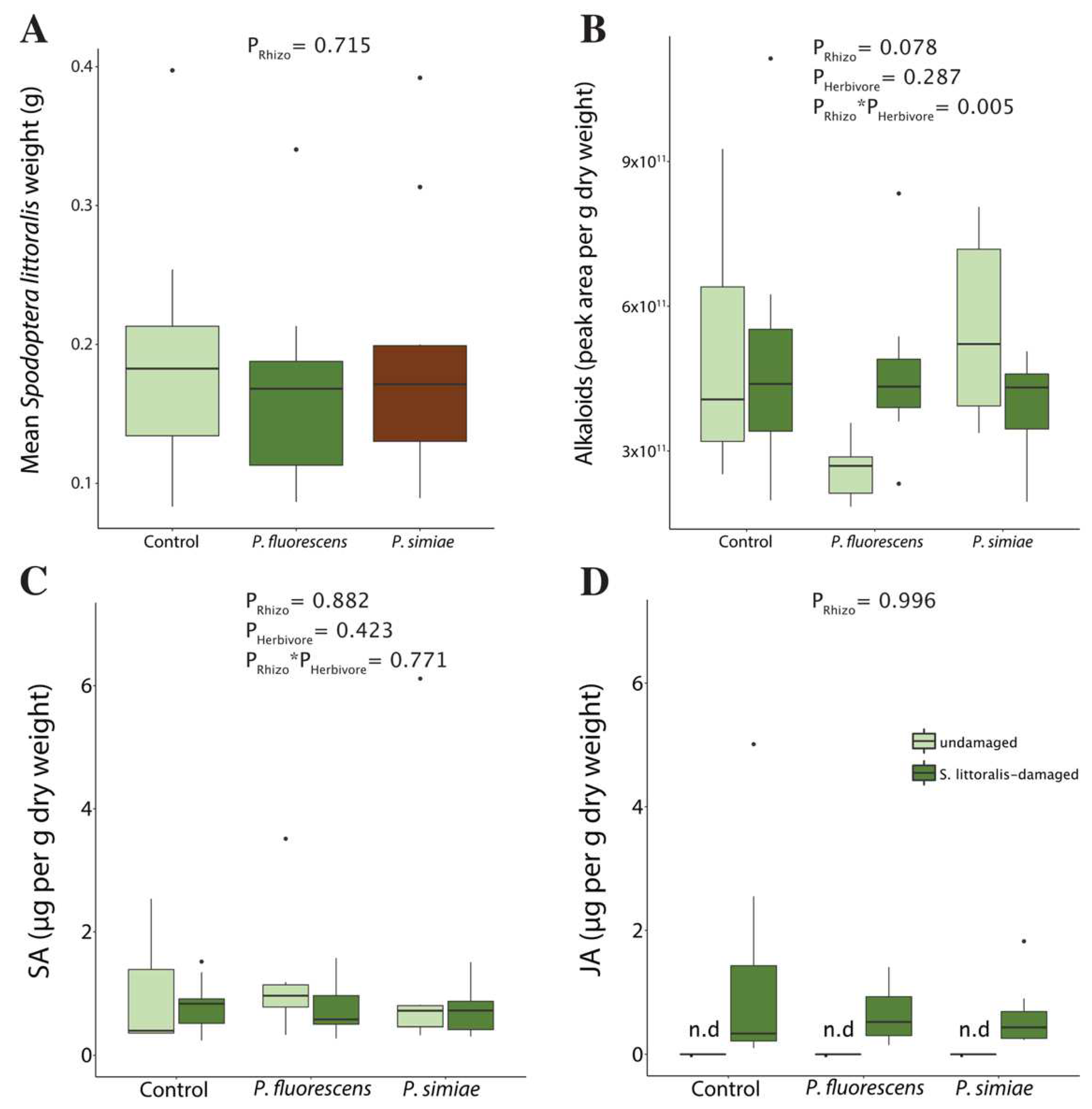
2.1. Inoculation with Rhizobacteria Did Not Affect Plant Biomass or Direct Resistance in Tomato
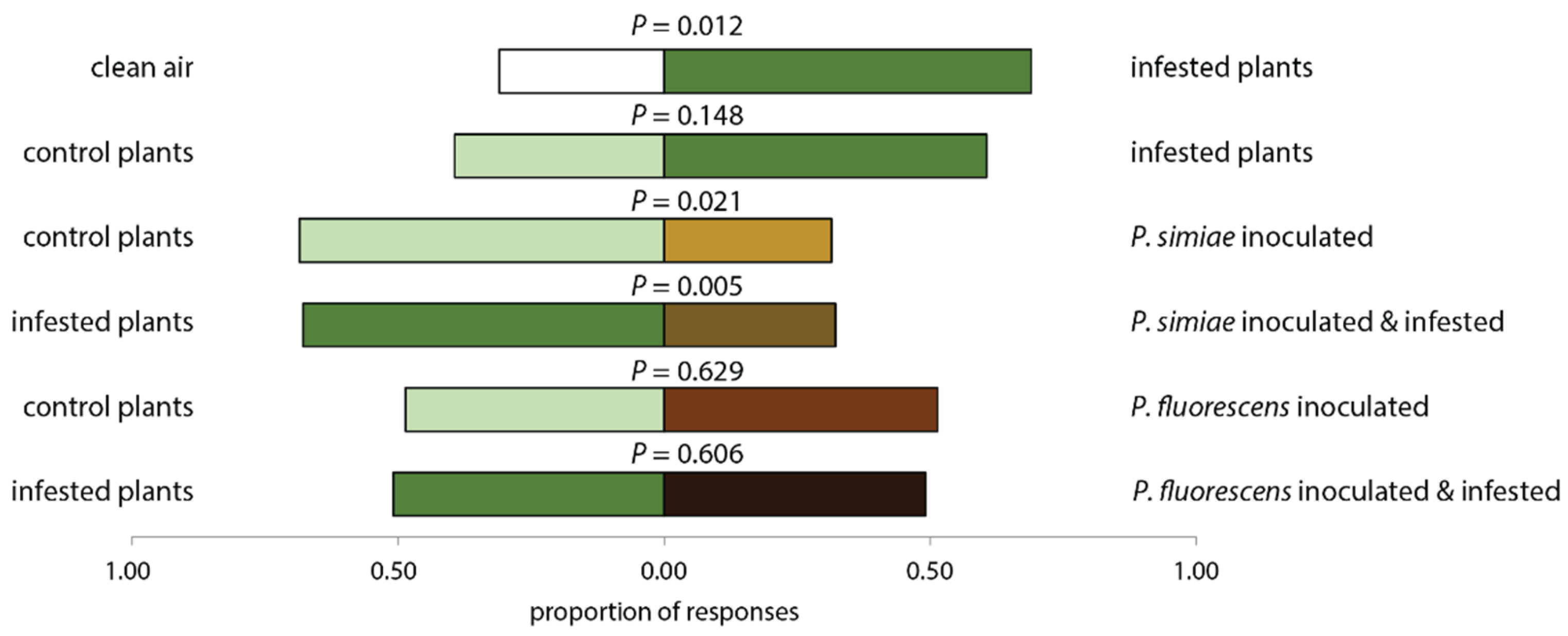
2.2. Inoculation with P. simiae Renders Tomato Plants Less Attractive to the Predator P. maculiventris
3. Discussion
4. Materials and Methods
4.1. Study System
4.2. Cultivation of Study Organisms
4.3. Plant Treatments
4.4. Rhizobacteria Effects on Plant Biomass and Direct Resistance
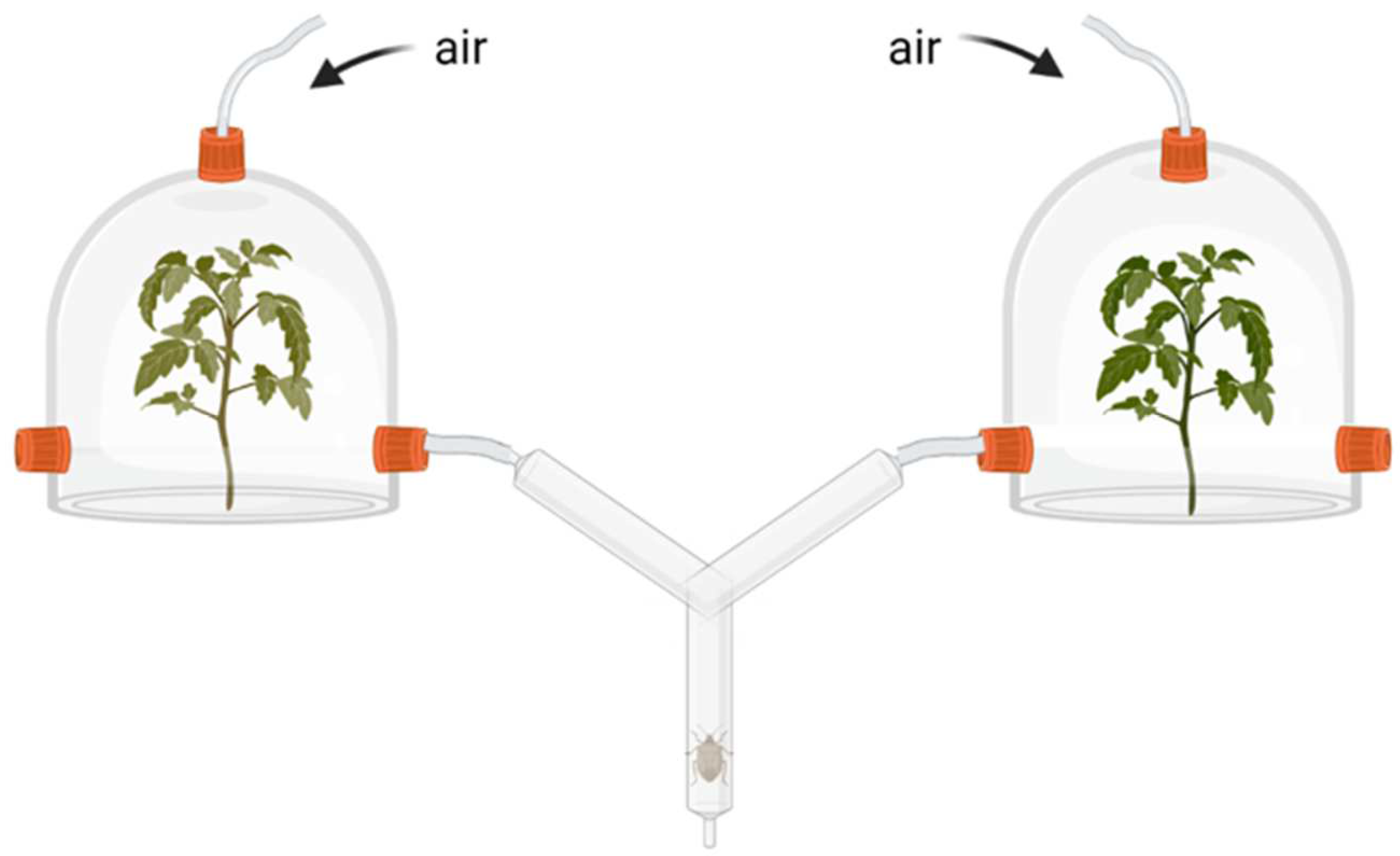
4.5. Rhizobacteria Effects on Plant Indirect Resistance
4.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Wees, S.C.M.; Van der Ent, S.; Pieterse, C.M.J. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Arif, I.; Batool, M.; Schenk, P.M. Plant Microbiome Engineering: Expected Benefits for Improved Crop Growth and Resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.M.; Mescher, M.C.; De Moraes, C.M. Plant dependence on rhizobia for nitrogen influences induced plant defenses and herbivore performance. Int. J. Mol. Sci. 2014, 15, 1466–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, J.M.; Mescher, M.C.; De Moraes, C.M. Plant-rhizobia mutualism influences aphid abundance on soybean. Plant Soil. 2009, 323, 187–196. [Google Scholar] [CrossRef]
- Pangesti, N.; Pineda, A.; Pieterse, C.M.; Dicke, M.; van Loon, J.J. Two-way plant mediated interactions between root-associated microbes and insects: From ecology to mechanisms. Front. Plant Sci. 2013, 4, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanakumar, D.; Lavanya, N.; Muthumeena, B.; Raguchander, T.; Suresh, S.; Samiyappan, R. Pseudomonas fluorescens enhances resistance and natural enemy population in rice plants against leaffolder pest. J. Appl. Entomol. 2008, 132, 469–479. [Google Scholar] [CrossRef]
- Babikova, Z.; Gilbert, L.; Bruce, T.J.; Birkett, M.; Caulfield, J.C.; Woodcock, C.; Pickett, J.A.; Johnson, D. Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack. Ecol. Lett. 2013, 16, 835–843. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, H.; Singh, V.K.; Singh, P.P.; Singh, S.K.; Ansari, W.A.; Yadav, A.; Singh, P.K.; Pandey, K.D. Role of Pseudomonas sp. in Sustainable Agriculture and Disease Management. In Agriculturally Important Microbes for Sustainable Agriculture—Volume 2: Applications in Crop Production and Protection; Meena, V.S., Mishra, P.K., Bisht, J.K., Pattanayak, A., Eds.; Springer: Singapore, 2017; Volume 2, pp. 195–215. [Google Scholar] [CrossRef]
- Pineda, A.; Kaplan, I.; Bezemer, T.M. Steering Soil Microbiomes to Suppress Aboveground Insect Pests. Trends Plant Sci. 2017, 22, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Pulido, H.; Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Combined effects of mutualistic rhizobacteria counteract virus-induced suppression of indirect plant defences in soya bean. Proc. R. Soc. B 2019, 286, 20190211. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.J.; Bloemberg, G.V. Life in the Rhizosphere. In Pseudomonas; Ramos, J.-L., Ed.; Springer: Boston, MA, USA, 2004; Volume 1, pp. 403–430. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.C.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockwell, V.O.; Stack, J.P. Using Pseudomonas spp. for Integrated Biological Control. Phytopathology 2007, 97, 244–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Mortel, J.E.; de Vos, R.C.; Dekkers, E.; Pineda, A.; Guillod, L.; Bouwmeester, K.; van Loon, J.J.; Dicke, M.; Raaijmakers, J.M. Metabolic and transcriptomic changes induced in Arabidopsis by the rhizobacterium Pseudomonas fluorescens SS101. Plant Physiol. 2012, 160, 2173–2188. [Google Scholar] [CrossRef] [Green Version]
- Pangesti, N.; Vandenbrande, S.; Pineda, A.; Dicke, M.; Raaijmakers, J.M.; van Loon, J.J. Antagonism between two root-associated beneficial Pseudomonas strains does not affect plant growth promotion and induced resistance against a leaf-chewing herbivore. FEMS Microbiol. Ecol. 2017, 93, fix038. [Google Scholar] [CrossRef] [Green Version]
- Pangesti, N.; Weldegergis, B.T.; Langendorf, B.; van Loon, J.J.; Dicke, M.; Pineda, A. Rhizobacterial colonization of roots modulates plant volatile emission and enhances the attraction of a parasitoid wasp to host-infested plants. Oecologia 2015, 178, 1169–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineda, A.; Soler, R.; Weldegergis, B.T.; Shimwela, M.M.; van Loon, J.J.; Dicke, M. Non-pathogenic rhizobacteria interfere with the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid signalling. Plant Cell Environ. 2013, 36, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Kappers, I.F.; Aharoni, A.; van Herpen, T.W.J.M.; Luckerhoff, L.L.P.; Dicke, M.; Bouwmeester, H.J. Genetic Engineering of Terpenoid Metabolism Attracts Bodyguards to Arabidopsis. Science 2005, 309, 2070–2072. [Google Scholar] [CrossRef] [PubMed]
- Nemec, S.; Lund, E. Leaf Volatiles of Mycorrhizal and Nonmycorrhizal Citrus Jambhiri Lush. J. Essent. Oil Res. 1990, 2, 287–297. [Google Scholar] [CrossRef]
- Babikova, Z.; Gilbert, L.; Randall, K.C.; Bruce, T.J.; Pickett, J.A.; Johnson, D. Increasing phosphorus supply is not the mechanism by which arbuscular mycorrhiza increase attractiveness of bean (Vicia faba) to aphids. J. Exp. Bot. 2014, 65, 5231–5241. [Google Scholar] [CrossRef] [Green Version]
- Duijff, B.J.; Pouhair, D.; Olivain, C.; Alabouvette, C.; Lemanceau, P. Implication of Systemic Induced Resistance in the Suppression of Fusarium Wilt of Tomato by Pseudomonas fluorescens WCS417r and by Nonpathogenic Fusarium oxysporum Fo47. Eur. J. Plant Pathol. 1998, 104, 903–910. [Google Scholar] [CrossRef]
- Yan, Z.; Reddy, M.S.; Ryu, C.M.; McInroy, J.A.; Wilson, M.; Kloepper, J.W. Induced systemic protection against tomato late blight elicited by plant growth-promoting rhizobacteria. Phytopathology 2002, 92, 1329–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrieri, E.; Lingua, G.; Digilio, M.C.; Massa, N.; Berta, G. Do interactions between plant roots and the rhizosphere affect parasitoid behaviour? Ecol. Entomol. 2004, 29, 753–756. [Google Scholar] [CrossRef]
- Pappas, M.L.; Liapoura, M.; Papantoniou, D.; Avramidou, M.; Kavroulakis, N.; Weinhold, A.; Broufas, G.D.; Papadopoulou, K.K. The Beneficial Endophytic Fungus Fusarium solani Strain K Alters Tomato Responses Against Spider Mites to the Benefit of the Plant. Front. Plant Sci. 2018, 9, 1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schausberger, P.; Peneder, S.; Jürschik, S.; Hoffmann, D. Mycorrhiza changes plant volatiles to attract spider mite enemies. Funct. Ecol. 2012, 26, 441–449. [Google Scholar] [CrossRef]
- Battaglia, D.; Bossi, S.; Cascone, P.; Digilio, M.C.; Prieto, J.D.; Fanti, P.; Guerrieri, E.; Iodice, L.; Lingua, G.; Lorito, M.; et al. Tomato below ground-above ground interactions: Trichoderma longibrachiatum affects the performance of Macrosiphum euphorbiae and its natural antagonists. Mol. Plant Microbe Interact. 2013, 26, 1249–1256. [Google Scholar] [CrossRef] [Green Version]
- Cappellari, L.D.R.; Chiappero, J.; Santoro, M.V.; Giordano, W.; Banchio, E. Inducing phenolic production and volatile organic compounds emission by inoculating Mentha piperita with plant growth-promoting rhizobacteria. Sci. Hortic. 2017, 220, 193–198. [Google Scholar] [CrossRef]
- Cappellari, L.D.R.; Santoro, M.V.; Schmidt, A.; Gershenzon, J.; Banchio, E. Improving Phenolic Total Content and Monoterpene in Mentha x piperita by Using Salicylic Acid or Methyl Jasmonate Combined with Rhizobacteria Inoculation. Int. J. Mol. Sci. 2019, 21, 50. [Google Scholar] [CrossRef] [Green Version]
- Sant’ana, J.; Da Silva, R.F.P.; Dickens, J.C. Olfactory Reception of Conspecific Aggregation Pheromone and Plant Odors by Nymphs of the Predator, Podisus maculiventris. J. Chem. Ecol. 1999, 25, 1813–1826. [Google Scholar] [CrossRef]
- Sant’ana, J.; Dickens, J.C. Comparative Electrophysiological Studies of Olfaction in Predaceous Bugs, Podisus maculiventris and P. nigrispinus. J. Chem. Ecol. 1998, 24, 965–984. [Google Scholar] [CrossRef]
- Dickens, J.C. Predator–prey interactions: Olfactory adaptations of generalist and specialist predators. Agric. For. Entomol. 1999, 1, 47–54. [Google Scholar] [CrossRef]
- Vuorinen, T.; Nerg, A.-M.; Ibrahim, M.A.; Reddy, G.V.P.; Holopainen, J.K. Emission of Plutella xylostella Induced Compounds from Cabbages Grown at Elevated CO2 and Orientation Behavior of the Natural Enemies. Plant Physiol. 2004, 135, 1984–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldrich, J.R.; Kochansky, J.P.; Abrams, C.B. Attractant for a Beneficial Insect and Its Parasitoids: Pheromone of the Predatory Spined Soldier Bug, Podisus maculiventris (Hemiptera: Pentatomidae). Environ. Entomol. 1984, 13, 1031–1036. [Google Scholar] [CrossRef]
- Ruberson, J.R.; Tauber, M.J.; Tauber, C.A. Plant Feeding by Podisus maculiventris (Heteroptera: Pentatomidae): Effect on Survival, Development, and Preoviposition Period. Environ. Entomol. 1986, 15, 894–897. [Google Scholar] [CrossRef]
- Gouinguene, S.P.; Turlings, T.C. The effects of abiotic factors on induced volatile emissions in corn plants. Plant Physiol. 2002, 129, 1296–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmelz, E.A.; Alborn, H.T.; Engelberth, J.; Tumlinson, J.H. Nitrogen deficiency increases volicitin-induced volatile emission, jasmonic acid accumulation, and ethylene sensitivity in maize. Plant Physiol. 2003, 133, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.N.; Hasanuzzaman, A.T.M.; Zhang, Z.-F.; Zhang, Y.; Liu, T.-X. High Level of Nitrogen Makes Tomato Plants Releasing Less Volatiles and Attracting More Bemisia tabaci (Hemiptera: Aleyrodidae). Front. Plant Sci. 2017, 8, 466. [Google Scholar] [CrossRef] [Green Version]
- Shavit, R.; Ofek-Lalzar, M.; Burdman, S.; Morin, S. Inoculation of tomato plants with rhizobacteria enhances the performance of the phloem-feeding insect Bemisia tabaci. Front. Plant Sci. 2013, 4, 306. [Google Scholar] [CrossRef] [Green Version]
- Löser, T.B.; Mescher, M.C.; De Moraes, C.M.; Maurhofer, M. Effects of Root-Colonizing Fluorescent Pseudomonas Strains on Arabidopsis Resistance to a Pathogen and an Herbivore. Appl. Environ. Microbiol. 2021, 87, e0283120. [Google Scholar] [CrossRef]
- Salama, H.S.; Dimetry, N.Z.; Salem, S.A. On the Host Preference and Biology of the Cotton Leaf Worm Spodoptera littoralis Bois. Z. Angew. Entomol. 1971, 67, 261–266. [Google Scholar] [CrossRef]
- De Clercq, P.; Degheele, D. A meat-based diet for rearing the predatory stinkbugs Podisus maculiventris and Podisus sagitta [Het.: Pentatomidae]. Entomophaga 1992, 37, 149–157. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Leeman, M.; Oorschot, M.; Sluis, I.; Schippers, B.; Bakker, P. Dose-response relationships in biological control of Fusarium wilt of radish by Pseudomonas spp. Phytopathology 1995, 85, 1075–1081. [Google Scholar] [CrossRef]
- Christensen, S.A.; Santana, E.A.; Alborn, H.T.; Block, A.K.; Chamberlain, C.A. Metabolomics by UHPLC-HRMS reveals the impact of heat stress on pathogen-elicited immunity in maize. Metabolomics 2021, 17, 6. [Google Scholar] [CrossRef] [PubMed]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Infection of host plants by Cucumber mosaic virus increases the susceptibility of Myzus persicae aphids to the parasitoid Aphidius colemani. Sci. Rep. 2015, 5, 10963. [Google Scholar] [CrossRef] [Green Version]
- Haber, A.I.; Sims, J.W.; Mescher, M.C.; De Moraes, C.M.; Carr, D.E. A key floral scent component (β-trans-bergamotene) drives pollinator preferences independently of pollen rewards in seep monkeyflower. Funct. Ecol. 2019, 33, 218–228. [Google Scholar] [CrossRef]
- Tikunov, Y.; Laptenok, S.; Hall, R.; Bovy, A.; de Vos, R.C. MSClust: A tool for unsupervised mass spectra extraction of chromatography-mass spectrometry ion-wise aligned data. Metabolomics 2011, 8, 714–718. [Google Scholar] [CrossRef] [Green Version]
- Lommen, A. MetAlign: Interface-driven, versatile metabolomics tool for hyphenated full-scan mass spectrometry data preprocessing. Anal. Chem. 2009, 81, 3079–3086. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Löser, T.B.; Lucas-Barbosa, D.; Maurhofer, M.; Mescher, M.C.; De Moraes, C.M. Negative Effects of Rhizobacteria Association on Plant Recruitment of Generalist Predators. Plants 2022, 11, 920. https://doi.org/10.3390/plants11070920
Löser TB, Lucas-Barbosa D, Maurhofer M, Mescher MC, De Moraes CM. Negative Effects of Rhizobacteria Association on Plant Recruitment of Generalist Predators. Plants. 2022; 11(7):920. https://doi.org/10.3390/plants11070920
Chicago/Turabian StyleLöser, Tobias B., Dani Lucas-Barbosa, Monika Maurhofer, Mark C. Mescher, and Consuelo M. De Moraes. 2022. "Negative Effects of Rhizobacteria Association on Plant Recruitment of Generalist Predators" Plants 11, no. 7: 920. https://doi.org/10.3390/plants11070920