
Seed Morphology in Species from the Silene mollissima Aggregate (Caryophyllaceae) by Comparison with Geometric Models




Abstract
:1. Introduction
2. Results
2.1. General Morphology
2.1.1. Lateral Views of Seeds
2.1.2. Dorsal View of Seeds
2.2. Comparison of the Average Silhouette of Seeds with the Geometric Models
2.2.1. Lateral View of Seeds
2.2.2. Dorsal View of Seeds
2.3. Shape Quantification with Models: The Mean J Index in the Seeds of Each Species
2.3.1. Shape Quantification in the Lateral Views with Models LM1, LM4 and LM5
2.3.2. Shape Quantification in the Dorsal Views with Models DM1, DM5 and DM6
3. Discussion
4. Materials and Methods
4.1. Seeds of Silene Analyzed
4.2. Seed Images
4.3. General Morphological Description
4.4. Obtention of an Average Silhouette
4.5. Geometric Models Used in the Comparisons
4.6. Comparison with Geometric Models: Calculation of the J Index
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A











References
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabler, R.K.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabler, R.K.; Oxelman, B. Notes on phylogeny and infrageneric classification of Silene, L. (Caryophyllaceae). Available online: https://doi.org/10.13140/RG.2.2.25580.39040 (accessed on 30 June 2020).
- Jeanmonod, D. Révision de la section Siphonomorpha Otth du genre Silene L. en Mediterranée occidentale III: Aggrégat italica et espèces affines. Candollea 1984, 39, 549–639. [Google Scholar]
- Jeanmonod, D. Révision de la section Siphonomorpha Otth du genre Silene L. (Caryophyllaceae) en Méditerranée occidentale II: Le groupe de S. mollissima. Candollea 1984, 39, 195–259. [Google Scholar]
- Jeanmonod, D. Révision de la section Siphonomorpha Otth du genre Silene L. en Mediterranée occidentale. V: Synthèse. Candollea 1985, 40, 35–56. [Google Scholar]
- Murru, V.; Santo, A.; Piazza, C.; Hugot, L.; Bacchetta, G. Seed germination, salt stress tolerance, and the effect of nitrate on three Tyrrhenian coastal species of the Silene mollissima aggregate (Caryophyllaceae). Botany 2015, 93, 881–892. [Google Scholar] [CrossRef]
- Murru, V.; Santo, A.; Gallo, M.; Cardona, C.; Boi, M.; Bacchetta, G. Comparative germination ecology and seedling growth of two Ibero-Levantine endemic species belonging to the Silene mollissima aggregate (Caryophyllaceae). Flora 2017, 227, 10–17. [Google Scholar] [CrossRef]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Brullo, S. Silene oenotriae (Caryophyllaceae): A new species from S Italy. Nord. J. Bot. 1997, 17, 649–652. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2005; p. 302. [Google Scholar]
- Arroyo Marín, J. Plant diversity in the region of the strait of Gibraltar, a multilevel approach. Lagascalia 1997, 19, 393–404. [Google Scholar]
- Galán de Mera, A.; Morales Alonso, R.; Vicente Orellana, J.A.; Cortés, J.E. Silene gazulensis sp. nov. (Caryophyllaceae) un nuevo endemismo del entorno del estrecho de Gibraltar. Acta Bot. Malac. 1999, 24, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Porceddu, M.; Santo, A.; Orrù, M.; Meloni, F.; Ucchesu, M.; Picciau, R.; Sarigu, M.; Cuena Lombrana, A.; Podda, L.; Sau, S.; et al. Seed conservation actions for the preservation of plant diversity: The case of the Sardinian Germplasm Bank (BG-SAR). Plant Sociol. 2017, 54, 111–117. [Google Scholar]
- Bacchetta, G.; Carta, A.; Paradis, G.; Piazza, C.; Peruzzi, L. Further insights into the taxonomy of the Silene nocturna species complex (Caryophyllaceae): A systematic survey of the taxa from Sardinia and Corsica. Phytotaxa 2014, 175, 037–044. [Google Scholar] [CrossRef] [Green Version]
- Camelia, I. Aspects regarding seeds morphology and germination peculiarities in sometaxa from Silene L. genera. J. Plant Dev. 2011, 18, 5–10. [Google Scholar]
- Fawzi, N.; Fawzy, A.; Mohamed, A. Seed morphological studies of some species of Silene L. (Caryophyllaceae). Int. J. Bot. 2010, 6, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Keshavarzi, M.; Mahdavinejad, M.; Sheidai, M.; Gholipour, A. Seed and pollen morphology of some Silene species (Caryophyllaceae) in Iran. Phytol. Balc. Int. J. Balkan Flora Veg. 2015, 21, 7–12. [Google Scholar]
- Yildiz, K. A morphological investigation of Silene L. (Caryophyllaceae) species distributed in West Anatolia and North Cyprus. Pak. J. Bot. 2006, 38, 67–83. [Google Scholar]
- Yildiz, K. Morphological and palynological investigation of Silene gigantea L. var. gigantea and Silene behen L. (Caryophyllaceae). Turk. J. Bot 2006, 30, 105–119. [Google Scholar]
- Mebatsion, H.K.; Paliwal, J.; Jayas, D.S. Evaluation of variations in the shape of grain types using principal components analysis of the elliptic Fourier descriptors. Comput. Electron. Agric. 2012, 80, 63–70. [Google Scholar] [CrossRef]
- Smykalova, I.; Grillo, O.; Bjelkova, M.; Pavelek, M.; Venora, G. Phenotypic evaluation of flax seeds by image analysis. Ind. Crops Prod. 2013, 47, 232–238. [Google Scholar]
- Lo Bianco, M.; Grillo, O.; Cañadas, E.; Venora, G.; Bacchetta, G. Inter- and intraspecific diversity in Cistus L. (Cistaceae) seeds, analysed with computer vision techniques. Plant Biol. 2017, 19, 183–190. [Google Scholar] [CrossRef]
- Lo Bianco, M.; Grillo, O.; Escobar Garcia, P.; Mascia, F.; Venora, G.; Bacchetta, G. Morpho-colorimetric characterization of Malva alliance taxa by seed image analysis. Plant Biol. 2017, 19, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Sarigu, M.; Porceddu, M.; Schmitt, E.; Camarda, I.; Bacchetta, G. Taxonomic discrimination of the Paeonia mascula group in the Tyrrhenian Islands by seed image analysis. Syst. Biodivers. 2019, 17, 801–810. [Google Scholar] [CrossRef]
- Farris, E.; Orrù, M.; Ucchesu, M.; Amadori, A.; Porceddu, M.; Bacchetta, G. Morpho-colorimetric characterization of the Sardinian endemic taxa of the genus Anchusa L. by seed image analysis. Plants 2020, 9, 1321. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, E.; Martín, J.J.; Ardanuy, R.; de Diego, J.G.; Tocino, Á. Modeling the Arabidopsis seed shape by a cardioid: Efficacy of the adjustment with a scale change with factor equal to the Golden Ratio and analysis of seed shape in ethylene mutants. J. Plant Physiol. 2010, 67, 408–410. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Tocino, Á.; Ardanuy, R.; de Diego, J.G.; Cervantes, E. Dynamic analysis of Arabidopsis seed shape reveals differences in cellulose mutants. Acta Physiol. Plant. 2014, 36, 1585–1592. [Google Scholar] [CrossRef]
- Cervantes, E.; Martín, J.J.; Chan, P.K.; Gresshoff, P.M.; Tocino, Á. Seed shape in model legumes: Approximation by a cardioid reveals differences in ethylene insensitive mutants of Lotus japonicus and Medicago truncatula. J. Plant Physiol. 2012, 169, 1359–1365. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, E.; Saadaoui, E.; Tocino, Á.; Martín-Gómez, J.J. Seed shape quantification in the model legumes: Methods and applications. In The Model Legume Medicago truncatula; de Brujin, F., Ed.; Wiley: Hoboken, NJ, USA, 2020; Chapter 3-1-6; pp. 92–98. [Google Scholar] [CrossRef]
- Cervantes, E.; Martín-Gómez, J.J. Seed Shape Description and Quantification by Comparison with Geometric Models. Horticulturae 2019, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, E.; Martín-Gómez, J.J.; Saadaoui, E. Updated Methods for Seed Shape Analysis. Scientifica 2016, 2016, 5691825. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, E.; Martín Gómez, J.J.; Gutiérrez del Pozo, D.; Silva Dias, L. An Angiosperm species dataset reveals relationships between seed size and two-dimensional shape. Horticulturae 2019, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Martín-Gómez, J.J.; Rewicz, A.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. Seed morphology in Silene based on geometric models. Plants 2020, 9, 1787. [Google Scholar] [CrossRef]
- Juan, A.; Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. New techniques for seed shape description in Silene species. Taxonomy 2022, 2, 1. [Google Scholar] [CrossRef]
- Rodríguez-Lorenzo, J.L.; Martín-Gómez, J.J.; Juan, A.; Tocino, Á.; Janoušek, B.; Cervantes, E. New geometric models for shape quantification of the dorsal view in seeds of Silene species. Plants 2022. submitted. [Google Scholar]
- Murru, V.; Grillo, O.; Santo, A.; Ucchesu, M.; Piazza, C.; Gaio, A.; Carta, A.; Bacchetta, G. Seed morpho-colorimetric analysis on some Tyrrhenian species of the Silene mollissima aggregate (Caryophyllaceae). Flora 2019, 258, 151445. [Google Scholar] [CrossRef]
- Thomson, P.A. Variations in Seed Size Within Populations of Silene dioica (L.) Clairv. in Relation to Habitat. Ann. Bot. 1981, 47, 623–634. [Google Scholar] [CrossRef]
- Murawska-Wlodarczyk, K.; Korzeniak, U.; Babst-Kostecka, A. Metalliferous habitats and seed microbes affect the seed morphology and reproductive strategy of Arabidopsis helleri. Plant Soil 2022, 470, 175–192. [Google Scholar] [CrossRef]
- Cavers, P.B.; Steel, M.G. Patterns of Change in Seed Weight Over Time on Individual Plants. Am. Nat. 1984, 124, 324–335. [Google Scholar] [CrossRef]
- Naciri, Y.; Du Pasquier, P.-E.; Lundberg, M.; Jeanmonod, D.; Oxelman, B. A phylogenetic circumscription of Silene sect. Siphonomorpha (Caryophyllaceae) in the Mediterranean Basin. Taxon 2017, 66, 91–108. [Google Scholar] [CrossRef]
- Bacchetta, G.; Fenu, G.; Mattana, E.; Piotto, B.; Virevaire, M. Manuale per la Raccolta, Studio, Conservazione e Gestione Ex Situ del Germoplasma; APAT: Rome, Italy, 2006; Volume 37. [Google Scholar]
- Bacchetta, G.; Bueno Sanchez, A.; Fenu, G.; Jiménez Alfaro, B.; Mattana, E.; Piotto, B.; Virevaire, M. Conservación Ex Situ de Plantas Silvestres; Obra Social La Caixa y Gobierno del Principado de Asturias; Principado de Asturias: Oviedo, Spain, 2008.
- Rasband, W.S.; ImageJ U.S. National Institutes of Health: Bethesda, MA, USA. 2018. Available online: http://imagej.nih.gov/ij/ (accessed on 21 February 2022).
- Zdilla, M.J.; Hatfield, S.A.; McLean, K.A.; Cyrus, L.M.; Laslo, J.M.; Lambert, H.W. Circularity, solidity, axes of a best fit ellipse, aspect ratio, and roundness of the foramen ovale: A morphometric analysis with neurosurgical considerations. J. Craniofacial Surg. 2016, 27, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, E.; Martín-Gómez, J.J.; Espinosa-Roldán, F.E.; Muñoz-Organero, G.; Tocino, Á.; Cabello-Sáenz de Santamaría, F. Seed Morphology in Key Spanish Grapevine Cultivars. Agronomy 2021, 11, 734. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Campbell, G.; Skillings, J.H. Nonparametric Stepwise Multiple Comparison Procedures. J. Am. Stat. Assoc. 1985, 80, 998–1003. [Google Scholar] [CrossRef]
- Sokal, R.R.; Braumann, C.A. Significance Tests for Coefficients of Variation and Variability Profiles. Syst. Zool. 1980, 29, 50. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
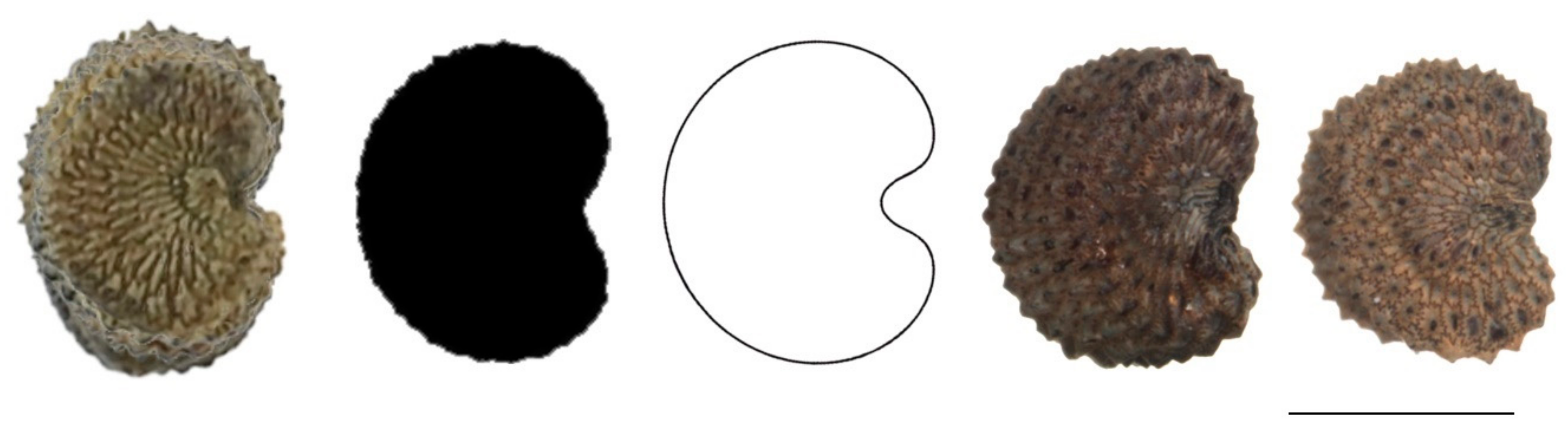{kind=link}
{kind=link}
| Species | N | A (mm2) | P (mm) | L (mm) | W (mm) | AR | C | R |
|---|---|---|---|---|---|---|---|---|
| S. andryalifolia | 30 | 1.50 e (9.28) | 4.98 f (5.02) | 1.55 e (5.29) | 1.23 d (5.22) | 1.27 bcd (5.06) | 0.76 d (3.60) | 0.79 bcd (4.67) |
| S. auricolifolia | 15 | 1.44 de (18.80) | 4.89 f (10.98) | 1.45 d (9.31) | 1.26 d (9.87) | 1.15 a (5.17) | 0.75 cd (6.27) | 0.87 e (4.97) |
| S. badaroi | 130 | 0.86 a (16.03) | 3.77 a (8.50) | 1.17 a (8.34) | 0.93 a (8.40) | 1.25 b (4.88) | 0.76 d (4.00) | 0.80 cd (4.82) |
| S. gazulensis | 48 | 0.90 a (11.74) | 4.01 c (6.57) | 1.22 b (6.63) | 0.94 a (7.01) | 1.28 cde (7.40) | 0.69 a (4.24) | 0.77 abc (7.08) |
| S. hicesiae | 130 | 0.87 a (14.91) | 3.90 b (7.45) | 1.21 b (8.25) | 0.92 a (8.50) | 1.32 e (8.30) | 0.72 b (4.12) | 0.76 a (8.11) |
| S. hifacensis | 282 | 1.19 bc (18.18) | 4.49 d (10.37) | 1.38 c (9.46) | 1.09 b (9.20) | 1.26 bc (4.11 | 0.74 c (5.99) | 0.80 cd (4.05) |
| S. ichnusae | 66 | 1.23 bc (17.21) | 4.56 de (9.79) | 1.41 cd (10.38) | 1.11 b (8.74) | 1.25 bc (7.97) | 0.75 c (5.39) | 0.79 cd (7.61) |
| S. mollissima | 40 | 1.16 bc (13.57) | 4.58 de (6.36) | 1.36 c (6.78) | 1.08 b (7.09) | 1.26 bc (3.33) | 0.69 a (3.71) | 0.80 cd (3.35) |
| S. oenotriae | 61 | 1.23 c (14.59) | 4.80 f (7.23) | 1.43 d (8.87) | 1.09 b (9.72) | 1.32 de (11.26) | 0.68 a (5.50) | 0.77 ab (10.73) |
| S. tomentosa | 44 | 1.15 b (10.38) | 4.62 e (6.28) | 1.34 c (5.17) | 1.09 b (6.38) | 1.23 b (4.54) | 0.68 a (4.87) | 0.81 d (4.61) |
| S. velutina | 135 | 1.38 d (15.01) | 4.83 f (7.91) | 1.53 d (8.43) | 1.15 c (9.70) | 1.33 e (9.82) | 0.74 c (3.98) | 0.76 a (9.80) |
| Species | N | A (mm2) | P (mm) | L (mm) | W (mm) | AR | C | R |
|---|---|---|---|---|---|---|---|---|
| S. andryalifolia | 24 | 1.33 d (8.90) | 4.88 d (5.91) | 1.60 e (6.53) | 1.06 e (4.49) | 1.52 bc (6.92) | 0.70 e (7.01) | 0.66 cd (6.81) |
| S. auricolifolia | 14 | 1.51 e (17.78) | 4.96 d (9.68) | 1.43 bc (8.04) | 1.33 f (10.27) | 1.08 a (4.46) | 0.77 f (7.64) | 0.93 e (4.46) |
| S. badaroi | 74 | 0.75 a (16.11) | 3.69 a (7.35) | 1.22 a (7.14) | 0.78 b (10.04) | 1.56 c (7.00) | 0.69 de (5.80) | 0.64 c (6.76) |
| S. gazulensis | 23 | 0.72 a (14.61) | 3.78 ab (9.12) | 1.24 a (7.95) | 0.73 a (7.18) | 1.71 def (5.29) | 0.63 b (5.67) | 0.59 ab (5.40) |
| S. hicesiae | 43 | 0.71 a (17.35) | 3.87 b (9.38) | 1.24 a (8.66) | 0.73 a (9.65) | 1.68 de (6.35) | 0.60 a (7.30) | 0.59 b (6.12) |
| S. hifacensis | 128 | 1.00 b (17.60) | 4.35 c (9.68) | 1.46 bc (9.56) | 0.87 c (8.79) | 1.68 d (5.76) | 0.66 c (5.08) | 0.60 b (5.69) |
| S. ichnusae | 20 | 0.96 b (16.82) | 4.26 c (7.67) | 1.39 b (9.49) | 0.87 c (8.75) | 1.60 c (8.44) | 0.66 c (8.77) | 0.63 c (8.19) |
| S. mollissima | 20 | 1.04 b (11.35) | 4.45 c (5.82) | 1.51 cd (5.20) | 0.87 c (7.20) | 1.74 ef (4.59) | 0.66 c (4.11) | 0.58 a (4.73) |
| S. oenotriae | 20 | 1.19 c (11.87) | 4.93 de (6.26) | 1.53 d (4.97) | 0.99 d (8.09) | 1.55 bc (7.40) | 0.61 a (7.18) | 0.65 cd (6.83) |
| S. tomentosa | 20 | 1.03 b (11.23) | 4.37 c (4.25) | 1.39 b (5.05) | 0.94 d (6.92) | 1.49 b (4.22) | 0.68 cd (5.06) | 0.67 d (4.26) |
| S. velutina | 39 | 1.23 c (13.57) | 5.12 e (7.37) | 1.66 f (6.15) | 0.94 d (9.94) | 1.78 f (9.90) | 0.59 a (9.43) | 0.57 a (9.60) |
| Species | N | LM1 | LM2 | LM3 | LM4 | LM5 | LM6 | LM7 | LM8 |
|---|---|---|---|---|---|---|---|---|---|
| S. andryalifolia | 24 | 92.0 | 91.8 | 88.3 | 91.9 | 91.9 | 84.2 | 88.5 | 89.0 |
| S. auricolifolia | 8 | 92.9 | 91.1 | 85.2 | 87.5 | 88.7 | 80.2 | 89.4 | 89.6 |
| S. badaroi | 70 | 91.0 | 91.0 | 90.4 | 93.4 | 91.7 | 86.3 | 89.3 | 90.3 |
| S. gazulensis | 32 | 90.6 | 88.4 | 90.4 | 90.8 | 93.3 | 87.7 | 91.4 | 90.0 |
| S. hicesiae | 48 | 88.1 | 87.3 | 89.2 | 91.7 | 90.1 | 88.7 | 87.5 | 86.8 |
| S. hifacensis | 128 | 91.9 | 91.0 | 90.4 | 93.8 | 92.0 | 86.3 | 89.1 | 90.5 |
| S. ichnusae | 20 | 92.9 | 91.7 | 88.3 | 90.5 | 91.9 | 82.9 | 89.4 | 91.8 |
| S. mollissima | 20 | 91.7 | 91.0 | 91.3 | 92.8 | 93.4 | 88.3 | 90.3 | 89.2 |
| S. oenotriae | 24 | 89.9 | 87.1 | 90.5 | 92.0 | 91.3 | 89.2 | 89.7 | 87.0 |
| S. tomentosa | 20 | 92.8 | 92.2 | 90.1 | 92.5 | 93.2 | 86.5 | 91.4 | 91.0 |
| S. velutina | 45 | 87.1 | 85.8 | 88.5 | 87.9 | 88.7 | 87.7 | 89.1 | 88.8 |
| Species | N | DM1 | DM5 | DM6 |
|---|---|---|---|---|
| S. andryalifolia | 24 | 90.5 | 93.7 | 90.1 |
| S. auricolifolia | 8 | 79.2 | 79.2 | 68.0 |
| S. badaroi | 70 | 91.9 | 94.2 | 90.9 |
| S. gazulensis | 32 | 92.0 | 91.7 | 93.1 |
| S. hicesiae | 48 | 89.2 | 91.0 | 91.4 |
| S. hifacensis | 128 | 91.4 | 92.5 | 93.4 |
| S. ichnusae | 20 | 92.0 | 92.8 | 92.9 |
| S. mollissima | 20 | 91.4 | 90.9 | 93.2 |
| S. oenotriae | 24 | 91.8 | 92.9 | 85.7 |
| S. tomentosa | 20 | 92.2 | 93.9 | 91.5 |
| S. velutina | 45 | 89.8 | 87.8 | 91.8 |
| Species | N | LM1 | LM4 | LM5 |
|---|---|---|---|---|
| S. andryalifolia | 24 | 89.2 c (1.64) | 88.8 cd (1.68) | 89.9 b (1.72) |
| S. auricolifolia | 8 | 91.1 d (1.80) | 85.2 ab (4.00) | 85.8 a (2.61) |
| S. badaroi | 70 | 89.8 cd (1.60) | 90.0 e (2.01) | 89.7 b (1.42) |
| S. gazulensis | 32 | 87.1 ab (2.80) | 88.2 bcd (2.22) | 89.3 b (2.07) |
| S. hicesiae | 48 | 86.1 a (3.18) | 88.1 bcd (2.76) | 86.8 a (2.66) |
| S. hifacensis | 128 | 90.2 cd (2.14) | 91.1 f (1.51) | 90.1 b (1.45) |
| S. ichnusae | 20 | 89.3 cd (2.41) | 88.2 bcd (2.70) | 87.8 a (2.44) |
| S. mollissima | 24 | 87.7 b (2.07) | 86.8 abc (3.27) | 86.2 a (2.59) |
| S. oenotriae | 24 | 86.2 a (3.83) | 87.8 bcd (2.25) | 86.8 a (2.84) |
| S. tomentosa | 20 | 90.1 cd (1.40) | 89.5 de (2.10) | 89.7 b (1.69) |
| S. velutina | 45 | 84.8 a (4.93) | 84.8 a (4.58) | 85.8 a (3.61) |
| Species | N | DM1 | DM5 | DM6 |
|---|---|---|---|---|
| S. andryalifolia | 24 | 87.3 cde (1.51) | 89.9 ef (1.43) | 87.8 bcde (3.43) |
| S. auricolifolia | 14 | 73.3 a (3.75) | 73.7 a (3.49) | 68.0 a (3.56) |
| S. badaroi | 69 | 88.0 de (2.27) | 90.1 f (1.56) | 88.4 bcde (2.78) |
| S. gazulensis | 24 | 85.2 bcde (2.87) | 85.4 bc (2.51) | 89.3 cde (1.54) |
| S. hicesiae | 40 | 83.9 bc (2.77) | 85.2 bc (2.98) | 88.8 bcde (1.64) |
| S. hifacensis | 120 | 85.4 bcde (2.76) | 86.3 bcd (2.67) | 90.4 de (1.42) |
| S. ichnusae | 20 | 84.8 bcd (3.28) | 86.6 cde (2.94) | 87.4 bc (3.31) |
| S. mollissima | 20 | 84.4 bc (2.26) | 84.6 bc (2.21) | 90.5 e (1.31) |
| S. oenotriae | 20 | 86.3 bcde (2.19) | 88.7 def (1.77) | 87.6 bcd (2.93) |
| S. tomentosa | 20 | 88.6 e (1.81) | 90.0 f (1.42) | 86.1 b (2.31) |
| S. velutina | 36 | 83.0 b (4.84) | 83.2 b (5.04) | 87.8 bcde (3.04) |
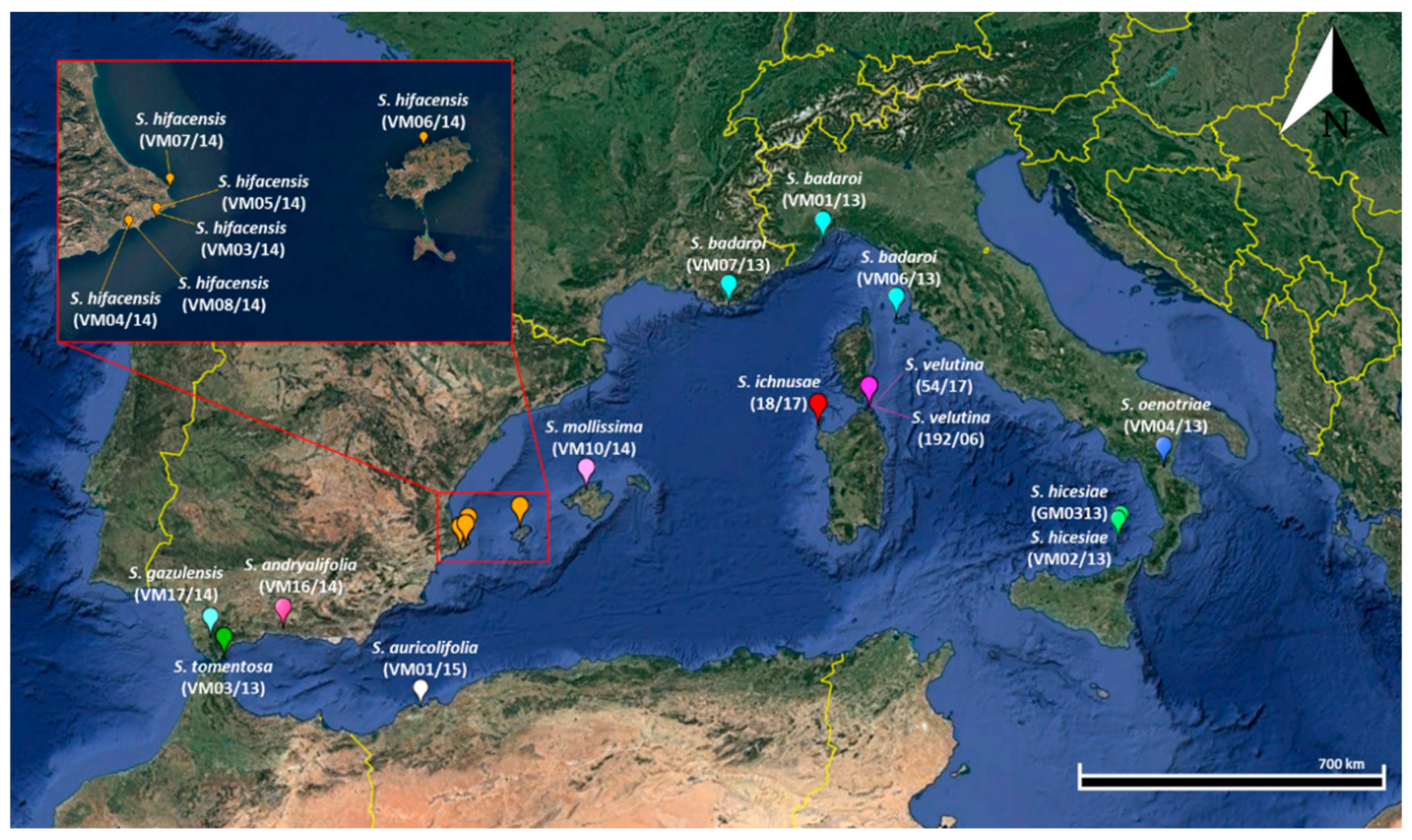
| Species | Accession Code in BG-SAR | Locality (Date of Collection) | Seed Number (in Stock) | Mean Coordinates (WGS 84) | Mean Elevation (m a.s.l.) |
|---|---|---|---|---|---|
| S. andryalifolia | VM16/14 | Canellas de Alabaida, Malaga, Spain (20 May 2014) | 30 | 36°50′ N; 3°59′ W | 500 |
| S. auricolifolia | VM01/15 | Santa Cruz, Orano, Algeria (12 May 2015) | 16 | 34°42 N; 00°40′ W | 316 |
| S. badaroi | VM01/13 | Capo Noli, Liguria, Italy (1 June 2013) | >200 | 44°11′ N; 8°25′ E | 240 |
| S. badaroi | VM06/13 | Marciana, Isola d’Elba, Tuscan Archipelago, Italy (6 July 2012) | >200 | 42°48′ N; 10°08′ E | 8 |
| S. badaroi | VM07/13 | Provence-Alpes-Côte d’Azur, Hyères, France (1 June 2013) | 41 | 42°04′ N; 6°06′ E | 10 |
| S. gazulensis | VM17/14 | Alcalá de los Gazules, Cadiz, Spain (15 June 2014) | 48 | 36°27′ N; 5°43′ W | 200 |
| S. hicesiae | GM0313 | Isole Eolie, Panarea, Sicily, Italy (2005) | 66 | 38°38′ N; 15°03′ E | 390 |
| S. hicesiae | VM02/13 | Isole Eolie, Panarea, Sicily, Italy (25 July 2013) | >200 | 38°38′ N; 15°03′ E | 390 |
| S. hifacensis | VM03/14 | Passebret, Cap D’Or, Alicante, Spain (1 July 2013) | >200 | 38°40′ N; 00°08′ E | 100 |
| S. hifacensis | VM04/14 | Morro de Toix, Calpe, Alicante, Spain (1 July 2013) | >200 | 38°37′ N; 00°01′ E | 56 |
| S. hifacensis | VM05/14 | Cova Cendres, Cap D’Or, Alicante, Spain (1 July 2013) | >200 | 38°41′ N; 00°09′ E | 80 |
| S. hifacensis | VM06/14 | Es Tossals, Cala Alabarca, Ibiza, Isole Baleari, Spain (16 June 2013) | 92 | 39°03′ N; 01°22′ E | 20 |
| S. hifacensis | VM07/14 | Illot de la Mona, Xabia, Alicante, Spain (1 July 2013) | 49 | 38°48′ N; 00°11′ E | 2 |
| S. hifacensis | VM08/14 | Morro de Toix, Calpe, Alicante, Spain (1 July 2014) | >200 | 38°37′ N; 00°01′ E | 56 |
| S. ichnusae | 18/17 | Capo Falcone, Stintino, Sardinia, Italy (1 July 2017) | 67 | 40°58′ N; 08°12′ E | 15 |
| S. mollissima | VM10/14 | Coma Freda, Maiorca, Isole Baleari, Spain (5 August 2014) | 96 | 39°48′ N; 02°52′ E | 650 |
| S. oenotriae | VM04/13 | Massiccio del Pollino, Basilicata, Italy (2 June 2013) | 62 | 39°49′ N; 16°19′ E | 350 |
| S. tomentosa | VM03/13 | Gibraltar Botanical Garden, Gibraltar (unknown) | 64 | 36°07′ N; 05°20′ W | 250 |
| S. velutina | 192/06 | Cala del Morto, Isola di La Maddalena, Sardinia, Italy (15 July 2006) | >200 | 41°14′ N; 09°24′ E | 3 |
| S. velutina | 54/17 | Abbattoggia, Isola di La Maddalena, Sardinia, Italy (12 August 2017) | 73 | 41°14′ N; 09°24′ E | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Gómez, J.J.; Porceddu, M.; Bacchetta, G.; Cervantes, E. Seed Morphology in Species from the Silene mollissima Aggregate (Caryophyllaceae) by Comparison with Geometric Models. Plants 2022, 11, 901. https://doi.org/10.3390/plants11070901
Martín-Gómez JJ, Porceddu M, Bacchetta G, Cervantes E. Seed Morphology in Species from the Silene mollissima Aggregate (Caryophyllaceae) by Comparison with Geometric Models. Plants. 2022; 11(7):901. https://doi.org/10.3390/plants11070901
Chicago/Turabian StyleMartín-Gómez, José Javier, Marco Porceddu, Gianluigi Bacchetta, and Emilio Cervantes. 2022. "Seed Morphology in Species from the Silene mollissima Aggregate (Caryophyllaceae) by Comparison with Geometric Models" Plants 11, no. 7: 901. https://doi.org/10.3390/plants11070901