
Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments

Jiangsu Engineering Research Center for Taxodium Rich, Germplasm Innovation and Propagation, Institute of Botany, Jiangsu Province and Chinese Academy of Sciences, Nanjing Botanical Garden, Memorial Sun Yat-Sen, Nanjing 210014, China
*
Author to whom correspondence should be addressed.
Plants 2022, 11(7), 894; https://doi.org/10.3390/plants11070894
Submission received: 14 February 2022
/
Revised: 23 March 2022
/
Accepted: 25 March 2022
/
Published: 28 March 2022
(This article belongs to the Special Issue Water and Nitrogen Management in Soil-Crop System)
Abstract
:To understand the characteristics of net NH4+ and NO3− fluxes and their relation with net H+ fluxes in Taxodium, net fluxes of NH4+, NO3− and H+ were detected by a scanning ion-selective electrode technique under different forms of fixed nitrogen (N) and experimental conditions. The results showed that higher net NH4+ and NO3− fluxes occurred at 2.1–3.0 mm from the root apex in T. ascendens and T. distichum. Compared to NH4+ or NO3− alone, more stable net NH4+ and NO3− fluxes were found under NH4NO3 supply conditions, of which net NH4+ flux was promoted at least 1.71 times by NO3−, whereas net NO3− flux was reduced more than 81.66% by NH4+ in all plants, which indicated that NH4+ is preferred by Taxodium plants. T. ascendens and T. mucronatum had the largest net NH4+ and total N influxes when NH4+:NO3− was 3:1. 15N Atom% and activities of N assimilation enzymes were improved by single N fertilization in the roots of T. distichum. In most cases, net H+ fluxes were tightly correlated with net NH4+ and NO3− fluxes. Thus, both N forms and proportions could affect N uptake of Taxodium. These findings could provide useful guidance for N management for better productivity of Taxodium plants.
1. Introduction
Nitrogen (N) plays a significant role in plant growth and development since it is a crucial component of plants’ chlorophylls, nucleic acids, proteins, and secondary metabolites [1]. Ammonium (NH4+) and nitrate (NO3−) are two primary forms of inorganic N absorbed and used by plants, and their fluxes in roots are varied with the distance from the apex. Spatial variability in the fluxes of NH4+ and NO3− has been explored along the roots in some herbaceous and woody plants [2,3,4,5]. For instance, the maximal net NH4+ influx happened at the root apex in rice (Oryza sativa L.) [2] and Populus simonii [5], whereas the highest net NH4+ influx appeared at 5 mm, 10 mm, and 5–20 mm from the root apex in lodgepole pine (Pinus contorta) [6], Populus popularis [7] and Douglas-fir (Pseudotsuga menziesii) [6], respectively. In the case of NO3−, previous studies observed that the highest net NO3− flux occurred at 0–10 mm in P. contorta [6], and at 15 mm from the apex in P. simonii [5] and P. popularis [7]. In rice, net NO3− influx increased to a maximum at 21 mm from the apex and then gradually declined [2]. Obviously, different plant species have distinct patterns of NH4+ and NO3− flux rates along the fine roots.
Apart from the spatial variation along the roots, NH4+ and NO3− fluxes are also affected by environmental factors such as N levels. A previous study in tea (Camellia sinensis) roots demonstrated increased net influxes of NH4+ and NO3− when the solution concentration increased from 0.2 mM to 1.2 mM under KNO3 and NH4Cl [1]. However, research on Picea glauca revealed a converse result in most cases; the roots presented net NH4+ and NO3− influxes in 50 μM with net effluxes in 1500 μM solutions [8]. When roots were treated in 10, 100, and 1000 μM NH4NO3 solutions, net NH4+ influxes increased gradually in P. popularis but decreased by degrees in Populus alba × Populus glandulosa [4]. In contrast, the hybrid presented higher net NO3− influxes than P. popularis in most cases [4]. This phenomenon revealed that the N concentrations in soils have prominent effects on the uptake of NH4+ and NO3−, and they are significantly related to the plant species. Additionally, previous studies revealed that interactions between NH4+ and NO3− exist on fluxes of both ions [7,9]. The interactions between NH4+ and NO3− are complicated among plants [10], and the underlying mechanisms remain unclear [7]. It is documented that the presence of NH4+ and NO3− negatively affect the uptake of each other, but NH4+ is preferred in C. sinensis [1]. However, net NH4+ influx was induced by the simultaneous provision of NO3−, and net NO3− influx was inhibited in the presence of NH4+ in roots of P. popularis and Populus asperata [7,11]. Moreover, a previous study on Douglas-fir and lodgepole pine showed that net NH4+ uptake remained unchanged in the presence or absence of NO3− [6]. Overall, interactions between NH4+ and NO3− and their preferences may result in changes of NH4+ and NO3− fluxes under different proportions of NH4+ and NO3− supply. Nonetheless, little information is available on the fluxes of NH4+ and NO3− in plant roots under fluctuating proportions of both inorganic N forms.
On the other hand, fluxes of NH4+ and NO3− are correlated with the plasma membrane PM-H+-ATPase activity that extrudes H+ from the cytosol to the outside at the expense of adenosine triphosphate (ATP) [12]. Previous research found that NO3− is transported across the plasma membrane via NO3−/H+ symporters with the involvement of PM-H+-ATPase [13]. The concentration of NH4+ can increase the activity of PM-H+-ATPase [14]. Furthermore, the expression of genes encoding PM-H+-ATPase was positively associated with fluxes of NH4+ and NO3− [8]. The significant correlations between NH4+, NO3− fluxes, and H+ uptake rate have been observed in many plants [12,15,16,17].
Taxodium species including T. ascendens, T. distichum, and T. mucronatum have been introduced from southeastern America to many countries owing to their economic and ecological benefits [18]. For instance, they can be used as woody bioenergy crops [19]. Taxodium oil showed adequate bioassay for insecticidal activity [20]. Compounds isolated from the bark can exhibit cytotoxic substances, thus treating against cancer cells [21]. Moreover, Taxodium plants have been selected as suitable species for afforestation in many challenging areas [22,23]. Although N is crucial for Taxodium growth and development, less information is available on the fluxes of NH4+ and NO3− as well as their correlation with H+ flux in fine roots. In this study, a non-invasive micro-electrodes technique was employed to investigate NH4+, NO3− and H+ fluxes in fine roots of T. ascendens, T. distichum, and T. mucronatum under different N forms and their proportions. Our objectives were (i) to determine the distance from the root apex of Taxodium plants where there are greater net NH4+ and NO3− fluxes; (ii) to illustrate the characteristics of NH4+, NO3− and H+ fluxes and their interactions under different N forms and proportions.
2. Materials and Methods
2.1. Plant Cultivation
Semi-lignified cuttings (10 cm in length, 0.3 cm in diameter) of T. ascendens, T. distichum, and T. mucronatum were selected. After being soaked in 3‰ 3-indoleacetic acid (IAA) solution for 2 min, they were repotted into a pot containing 1:1 volume of peat: perlite in a ventilated greenhouse at the Institute of Botany, Jiangsu Province and Chinese Academy of Sciences (35° 03′ N, 118° 49′ E), under normal growth conditions (approximately 25 °C) in a photoperiod of 14/10 h of light/dark. Two months later, cuttings with uniform size and development were selected for the NMT experiments.
Seeds were collected from a healthy T. distichum grown in the Institute of Botany, Jiangsu Province and Chinese Academy of Sciences. They were planted in black plastic pots (5 × 5 × 15 cm) filled with 1:1 volume of peat:perlite in a climate chamber (23–25 °C/15–18 °C, day/night; light per day, 14 h; photosynthetic photon flux, 160 μmol m−2 s−1; relative air humidity, 50–60%). After 19 weeks of growth, the plants were transferred into black plastic boxes (25 × 15 × 14.5 cm, 4 plants per box) containing 4 L of modified 1/4 Hoagland’s nutrient solution [24]. All nutrient solutions were continuously aerated with an air pump, and each solution was refreshed every other day. After 16 d, plants were used to explore 15N Atom% and enzymatic activities.
2.2. Experimental Design
To determine the positions along the root where the maximal influxes of NH4+ and NO3− occur, a preliminary experiment was carried out at 14 positions, in turns, 0, 0.3, 0.6, 0.9, 1.2, 1.5, 1.8, 2.1, 2.5, 3.0, 5.0, 8.0, 15.0 and 30.0 mm away from the root apex. The measuring solution was 0.5 mM MES (2-(N-Morpholino) ethanesulfonic acid hydrate buffer.), pH 6.0, to which either 1.0 mM NH4Cl for NH4+ or 1.0 mM KNO3 for NO3− was added. After that, the position where the greater net uptake of NH4+ and NO3− occurred was detected to carry out the following experiments.
To investigate the net fluxes of NH4+ and NO3− under different N forms and proportions, the measuring solutions were designed as (1) NH4+ (NH4Cl): 0.1 and 1.0 mM, (2) NO3− (KNO3): 0.1 and 1.0 mM and (3) NH4+:NO3−: 1:3, 1:1, and 3:1 (total N = 2 mM), containing 0.5 mM MES, pH 6.0.
To explore the biomass, 15N Atom% and enzymatic activities under single N fertilization, 24 seedlings with similar performance (ca. 15 cm in height) were selected and divided into three groups (8 plants in each group). Three N treatments: 0 mM 15NH4Cl and K15NO3 (serving as control, CK), 1 mM 15NH4Cl and 1 mM K15NO3 in 1/4 modified Hoagland’s nutrient solution [24] were applied. Dicyandiamide (7 μM, C2H4N4) was added into the nutrient solution to inhibit nitrification [19]. After 3 d, 4 plants from each treatment were harvested and used for measurements of 15N Atom%, and the remaining 4 plants in each treatment were used for enzymatic activities.
2.3. Measurement of NH4+, NO3− and H+ Fluxes
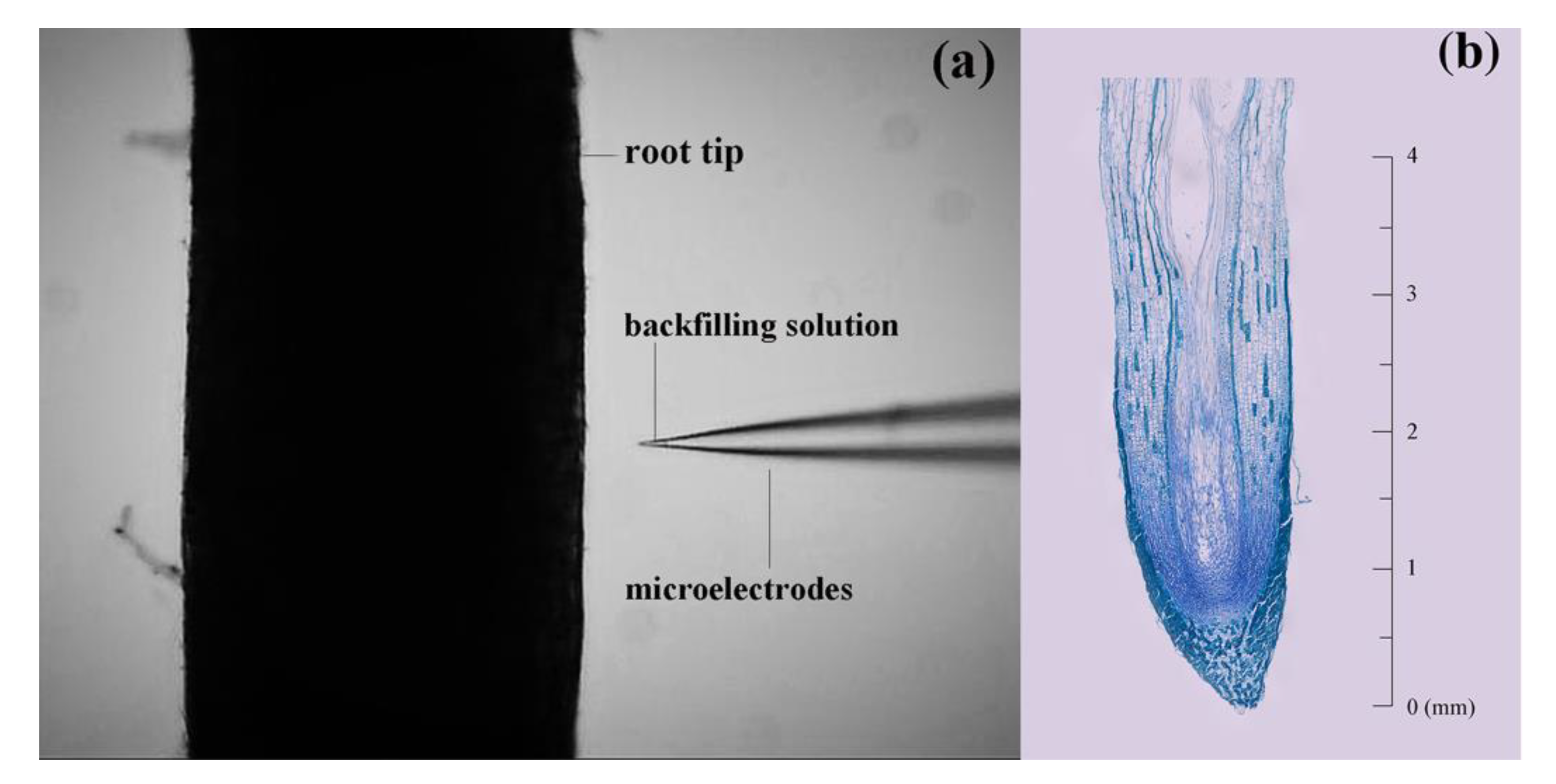
To understand the real-time NH4+, NO3− and H+ uptake by the fine roots under different treatments, ions flux alterations on the root surface were measured by using a non-invasive micro-test technology (NMT) system (youngerusa.com; xuyue.net) (Figure 1a).
The measurement procedures were described by Zhao et al. [5]. Firstly, ion-selective microelectrodes designed with 2–4 μm apertures were manufactured and silanized. Secondly, for the NH4+ electrode, in sequence, 100 mM NH4Cl was used as a backfilling solution, followed by an NH4+ selective liquid ion exchange cocktail (#09879, Sigma, St. Louis, MI, USA). Similarly, for the NO3− electrode, 10 mM KNO3 was used as the backfilling solution, followed by a NO3− selective liquid ion exchange cocktail (#72549, Sigma). For the H+ electrode, 15 mM NaCl and 40 mM KH2PO4 were used as backfilling solution, followed by an H+ selective liquid ion exchange cocktail (#95293, Sigma). Prior to the flux measurements, the microelectrodes were calibrated. For NH4+ calibration, 0.05/0.5 mM NH4Cl in addition to other compounds (0.5 mM MES, pH 6.4/5.4) were used in the measuring solution; for NO3− calibration, 0.05/0.5 mM KNO3 in addition to the compounds (0.5 mM MES, pH 6.4/5.4) were used in the measuring solution; for H+ calibration, pH 6.4/5.4 in addition to 0.5 mM MES were used in the measuring solution. Only electrodes with Nernstian slopes higher than 55 mV per tenfold concentration difference were used.
After that, fine white roots, 15–35 mm from the apex, were selected. They were fixed at the bottom of the petri dish filled with 10–20 mL measuring solutions for 20 min. After being equilibrated, the samples were transferred to another petri dish containing 5 mL fresh solution and then were put under the microscope. The tips of the microelectrodes were aligned and kept 30 μm away from the target point, which is a specific distance from the apex of the root. Net fluxes of NH4+/NO3− were recorded at each measurement point for 5 min. Not only eight biological repetitions (eight fine roots from four plants) but also 50 measurement time points in each repetition were considered.
2.4. Determination of 15N Uptake and Enzyme Activities
The roots were harvested and rinsed three times in distilled water. The 15N Atom% (15N AT%) and the amount of plant N derived from 15N-labeled fertilizer (Ndff%) were detected [25].
Activities of nitrate reductase (NR, EC 1.7.99.4), nitrite reductase (NiR, EC 1.7.2.1), glutamine synthetase (GS, EC 6.3.1.2), glutamate synthetase (GOGAT, EC 1.4.7.1) and glutamate dehydrogenase (GDH, EC 1.4.1.2) in the roots were assayed [4].
2.5. Statistical Analysis
In order to determine the NH4+, NO3− and H+ fluxes along the root tip, imFluxes V2.0 (xuyue.net) was used to obtain the data at each measuring point. The positive values represent net influxes, and the net negative values represent net effluxes. To analyze data for the ion fluxes, biomass, 15N AT% and enzyme activities, one-way ANOVA (Duncan’s multiple range tests at 5% level) was performed with the SPSS 25.0 (Statistical Product and Service Solutions, IBM, New York, NY, USA). GraphPad Prism version 9.1 was used to draw figures.
3. Results
3.1. Net Fluxes of NH4+ and NO3− along the Root Tip
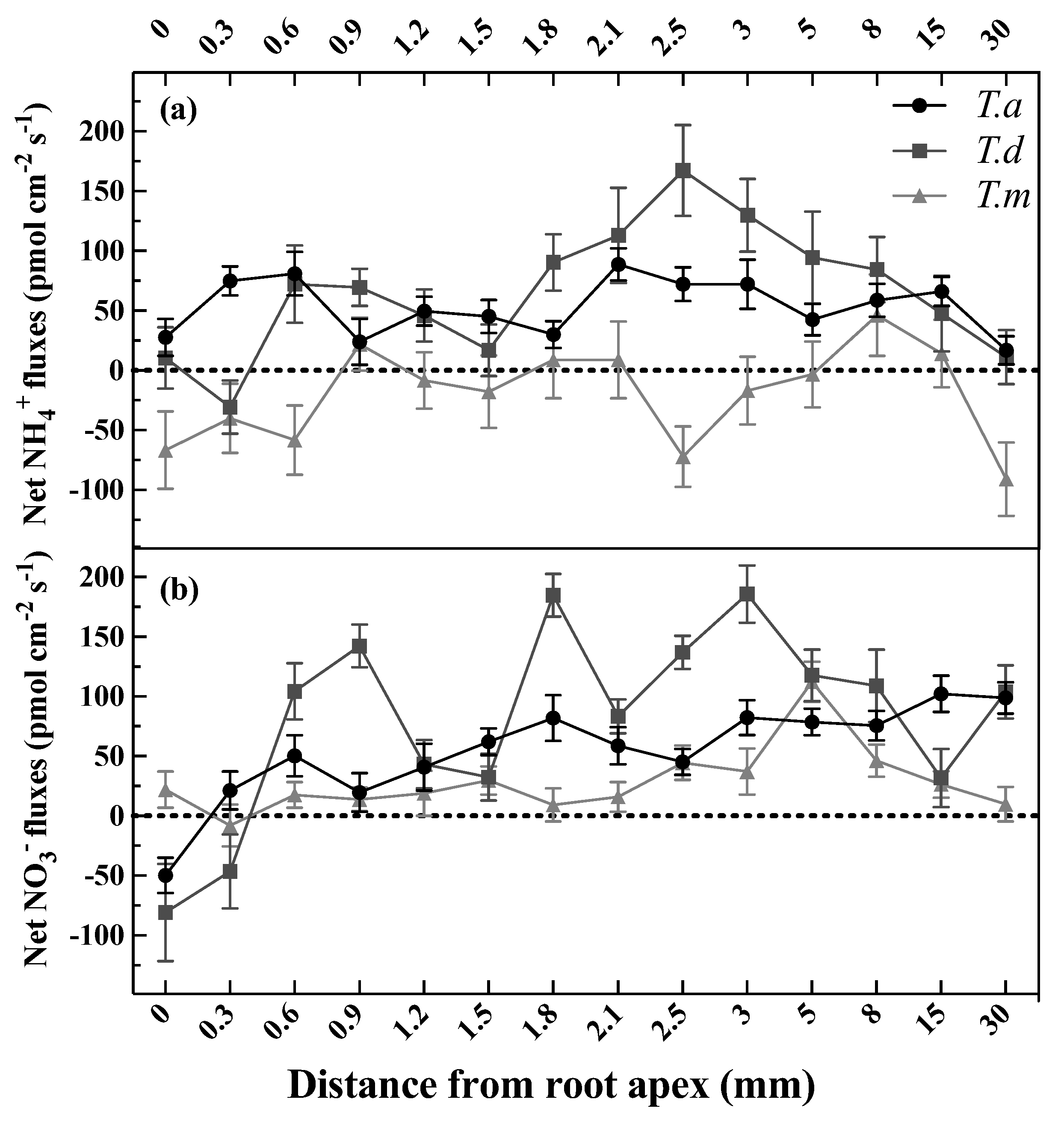
Net fluxes of NH4+ and NO3− were determined along the root tip up to 30.0 mm from the apex, and their fluxes were widely varied at different locations (Figure 2a,b). The NH4+ fluxes ranged from 17.02 (net influx) to 88.89 (net influx) pmol cm−2 s−1 in T. ascendens, and varied dramatically from −30.52 to 167.15 pmol cm−2 s−1 in T. distichum, and fluctuated between −90.94 and 46.31 pmol cm−2 s−1 in T. mucronatum when supplied as 1 mM NH4Cl (Figure 2a). Intriguingly, both T. ascendens and T. distichum showed strong NH4+ uptake rates from 2.1 to 3.0 mm along the root tip (Figure 2a).
Net NO3− fluxes ranged from −49.68 to 102.15 pmol cm−2 s−1 in T. ascendens, from −80.61 to 185.98 pmol cm−2 s−1 in T. distichum, and from −7.88 to 112.27 pmol cm−2 s−1 in T. mucronatum when fed with 1 mM KNO3 (Figure 2b). The maximal net NO3− influxes of T. ascendens, T. distichum, and T. mucronatum were detected at 15.0, 3.0, and 5.0 mm from the root apex, respectively (Figure 2b). As a result, the following experiments selected 2.5 mm from the root apex as the specific position to investigate the net fluxes of NH4+ and NO3−. Moreover, 2.5 mm from the apex belongs to the elongation zone of the root tip in Taxodium plants (Figure 1b).
3.2. Net Fluxes of NH4+ and NO3− under Different N Forms
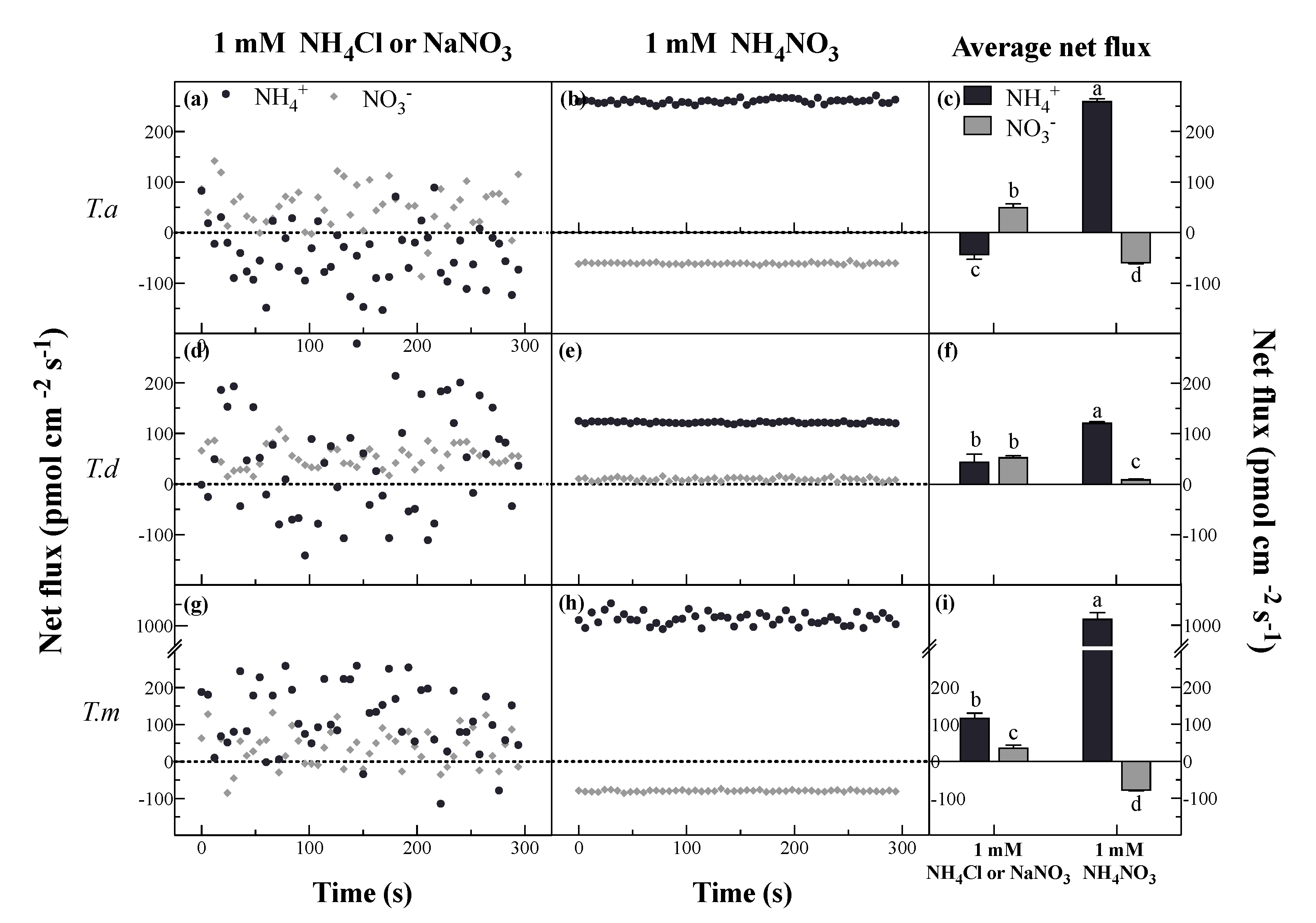
As NH4Cl and KNO3 were added separately, the NH4+ and NO3− fluxes fluctuated widely for all tested plants at 2.5 mm from the root apex during a 5-min period (Figure 3). Both NH4+ and NO3− fluxes of T. distichum and T. mucronatum showed a tendency towards net influx. T. ascendens, however, tended to show net efflux of NH4+ and net influx of NO3− (Figure 3a,d,g). When supplied with mixed N (NH4NO3), stable fluxes of NH4+ and NO3− were observed, and distinctly, NH4+ fluxes were much greater than NO3− fluxes in all Taxodium plants (Figure 3b,e,h). Compared to 1 mM NH4Cl, average net fluxes of NH4+ were stimulated by 688%, 171%, and 762% under 1 mM NH4NO3 in roots of T. ascendens, T. distichum, and T. mucronatum, respectively (Figure 3c,f,i). Thus, the increase of NH4+ fluxes was as follows: T. mucronatum > T. ascendens > T. distichum (Figure 3c,f,i). The same order was observed for the decreases in net NO3− fluxes, which were decreased by 314%, 220%, and 81.66% under 1 mM NH4NO3 compared with that under 1 mM KNO3, respectively (Figure 3c,f,i).
3.3. Net NH4+, NO3− and H+ Fluxes under Different N Concentrations
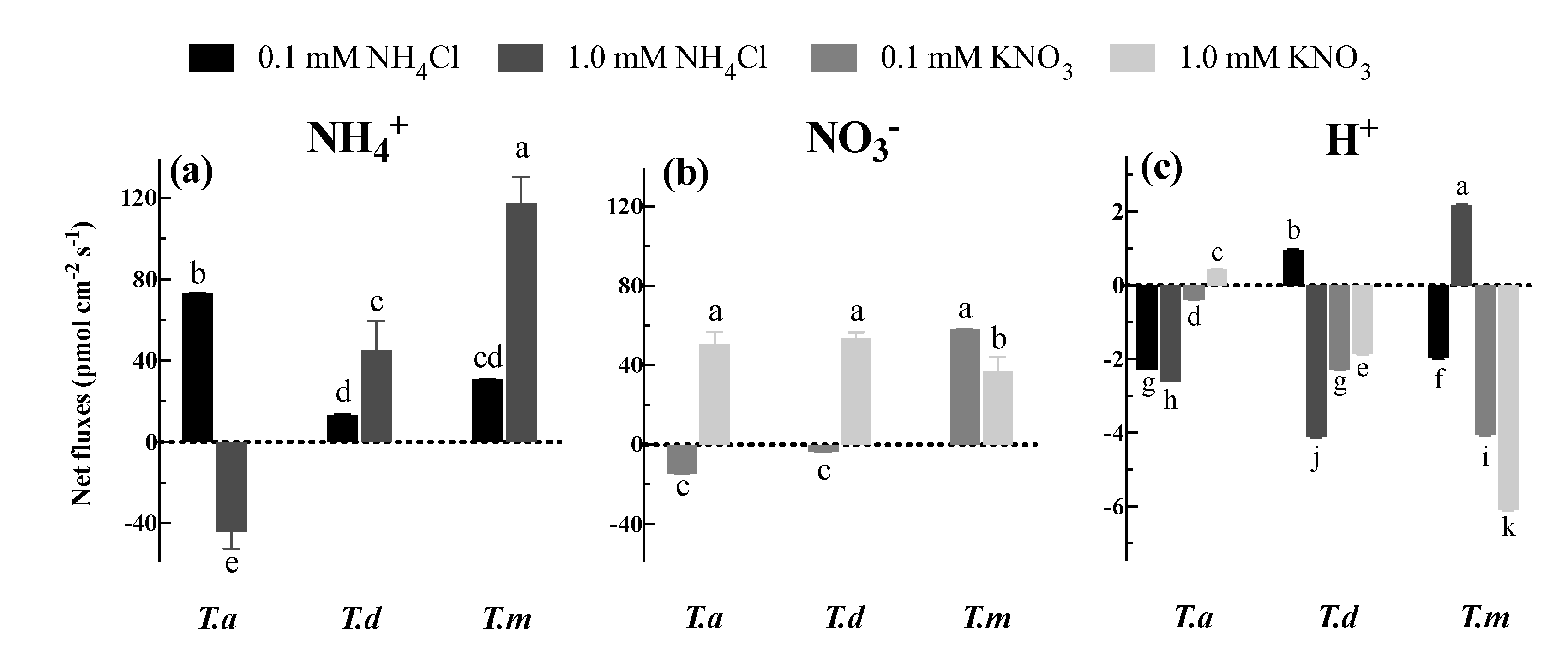
Except for T. ascendens exposed to 1.0 mM NH4Cl, all three species showed a tendency for net NH4+ influx when fed with 0.1 or 1.0 mM NH4Cl (Figure 4a). Additionally, the net influx of NH4+ in T. ascendens was significantly greater (p < 0.05) than those in the other two species under 0.1 mM NH4Cl (Figure 4a). Compared to 0.1 mM NH4Cl, net influx of NH4+ was promoted by 2.40 and 2.84 times under 1.0 mM NH4Cl treatment in T. distichum and T. mucronatum, respectively (Figure 4a). Apart from T. ascendens and T. distichum treated with 0.1 mM KNO3, all the plants displayed a tendency for net NO3− influx when supplied with 0.1 or 1.0 mM KNO3 (Figure 4b). Moreover, the fluxes of NO3− were significantly lower (p < 0.05) in 1.0 mM than in 0.1 mM KNO3 in T. mucronatum (Figure 4b).
At the same time, net H+ fluxes were determined in this study (Figure 4c). Here, we found that H+ presented net effluxes under all treatments except for T. ascendens under 1.0 mM KNO3, T. distichum under 0.1 mM NH4Cl, and T. mucronatum under 1.0 mM NH4Cl (Figure 4c). Other than T. distichum exposed to 0.1 mM NH4Cl, the change tendency of net H+ fluxes was similar to the variations of net NH4+ and NO3− fluxes when the solution concentration was increased from 0.1 mM to 1.0 mM (Figure 4c).
3.4. Net NH4+ and NO3− Fluxes under Different N Proportions
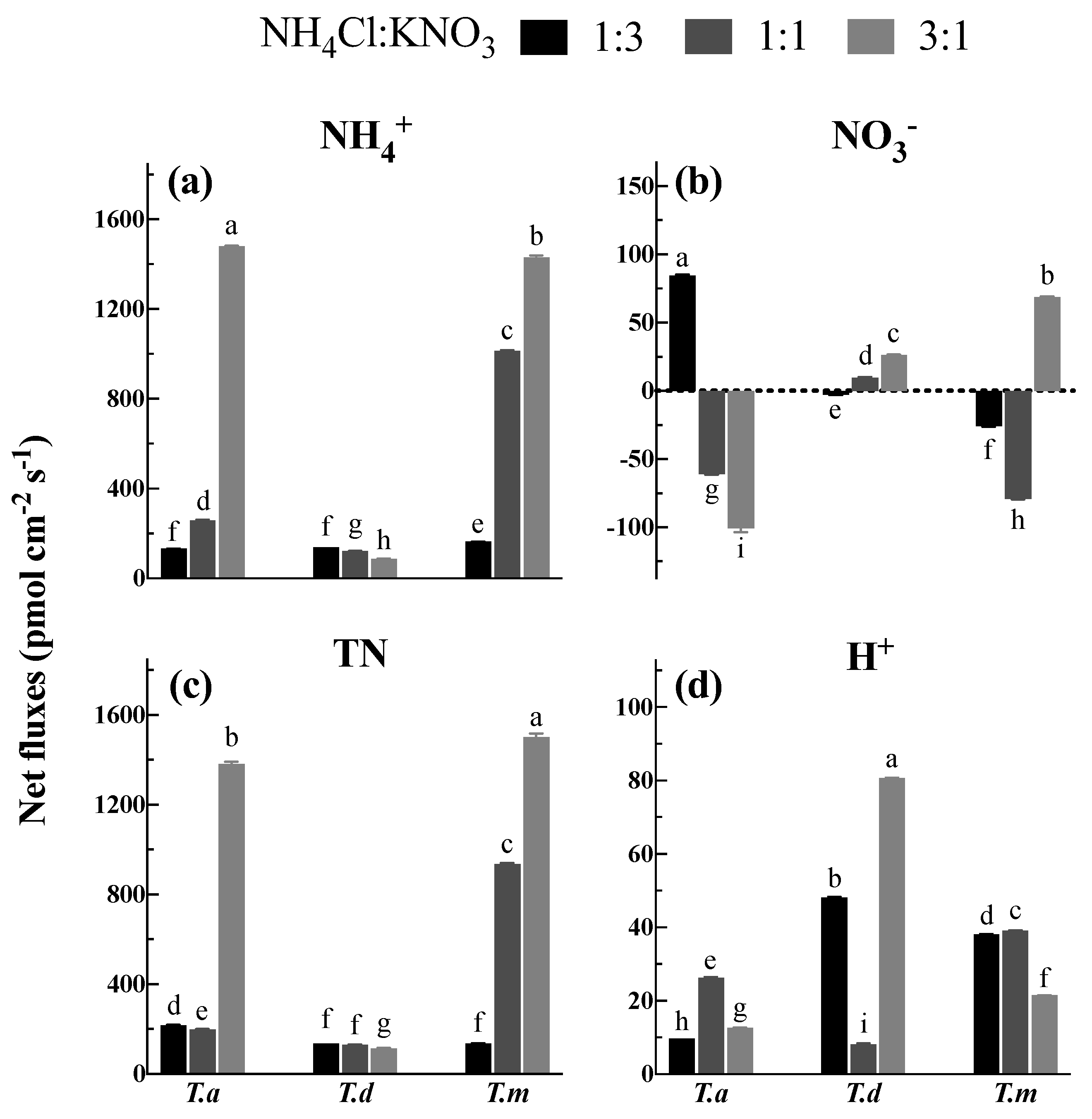
Under 2.0 mM TN (total nitrogen) consisting of various proportions of NH4Cl and KNO3 (1:3, 1:1, and 3:1), T. ascendens, T. distichum, and T. mucronatum showed great diversities in the fluxes of NH4+, NO3− and TN (Figure 5). The net influx of NH4+ ranged from 88.00 to 1480.80 pmol cm−2 s−1 across the three tested species (Figure 5a). It was 0.96 and 10.15 times greater under NH4: NO3 at 1:1 and 3:1 than at 1:3 in T. ascendens, respectively (Figure 5a). Similarly, the net NH4+ influxes were 5.21 and 7.76 times higher than 1:3 when treated with 1:1 and 3:1, respectively, in T. mucronatum (Figure 5a). A decreasing trend of net NH4+ influx was observed for T. distichum with an increase in the NH4+ proportion (Figure 5a).
Compared with the net NH4+ influx, the net flux of NO3− was much lower, ranging from −100.53 to 84.85 pmol cm−2 s−1 under different proportions of NH4Cl and KNO3 (Figure 5b). It is surprising that the net influx of NO3− observed under the 1:3 solution was replaced by net efflux when the proportion changed to 1:1 and 3:1 in T. ascendens (Figure 5b). T. distichum, however, presented a totally converse trend whereby the net flux of NO3− significantly (p < 0.05) increased by 1.68 times when the NH4+ proportion was raised from 1:1 to 3:1 (Figure 5b). In the case of T. mucronatum, there was net efflux under 1:3 and 1:1 and net influx under 3:1 (Figure 5b).
The trend of TN fluxes ranging from 114.23 to 1500.48 pmol cm−2 s−1 was similar to the fluxes of NH4+ (Figure 5c), and the highest net influx of TN was observed for T. mucronatum and T. ascendens when the proportion of NH4Cl: KNO3 was 3:1 (Figure 5c). Among the three species, T. distichum displayed the lowest net NH4+, NO3− and TN fluxes in all measuring solutions (Figure 5c).
3.5. 15N AT%, Ndff% and Enzyme Activities in the Roots of T. distichum
Compared with CK, 15N AT% was elevated by 70.27% or 29.73% in 1 mM 15NH4+-treated or 1 mM 15NO3−-treated T.distichum roots (Table 1). Similar results were also observed in Ndff% (Table 1). Compared to CK, however, no significant difference was found in the root biomass of T. distichum supplied with 1 mM 15NH4+ or 15NO3− during the 3 d experiment period (Table 1).
15NH4+ or 15NO3− fertilization also has positive impacts on the activities of N assimilation enzymes (Table 2). When compared with CK, NR, NiR, GS, GDH and GOGAT activities were enhanced by 50.63%, 33.74%, 39.40%, 48.25% and 59.36%, respectively in 1 mM 15NH4+-supplied T. distichum roots (Table 2). Similarly, the activities of NR and NiR were increased by 221.03% and 11.93%, respectively in 1 mM 15NO3−-fed T. distichum roots (Table 2).
4. Discussion
4.1. Spatial Variability of Net NH4+ and NO3− Fluxes along the Fine Roots
Fine roots consist of four distinct regions, including root cap, meristematic, elongation, and maturation zones, characterized by different anatomical and functional features [7]. These anatomical and functional diversities could bring about distinct absorbing abilities for NH4+ and NO3− in different root zones [26,27,28]. Spatial variability of net NH4+ and/or NO3− flux has been observed in fine roots of various plant species [29]. For example, maximal net NH4+ influx occurred at the root apex in rice [2] and P. simonii [5], and at 5 mm, 10 mm, and 5–20 mm from the root apex in P. contorta [6] and P. popularis [7], and Douglas-fir [6] respectively. Such spatial variation of net NH4+ and NO3− influxes along the root axis was also observed in our research. The largest net influxes of NH4+ and NO3− were detected at m from the apex of T. ascendens and T. distichum, which belongs to the elongation zone. Such differences are possibly because of cytosolic concentrations of NH4+ and NO3− in the elongation zone being lower than the thresholds needed for N assimilation to support the fast growth [30,31]. Similar results were observed in studies of Arabidopsis, where larger net NH4+ fluxes were shown in the elongation zones [28,32]. Moreover, Phyllostachys edulis showed relatively higher net influxes of NH4+ and NO3− at 2–5 mm from the root apex [30]. The net NH4+ or NO3− fluxes were found to be higher in segment I (0–35 mm) than segment II (35–70 mm) in Populus × canescens [31]. In addition, we found that T. distichum had the greatest NH4+ and NO3− uptake rates among the three Taxodium species.
4.2. Net NH4+ and NO3− Fluxes under Single N Treatments
Generally, environmental N levels have a significant impact on the NH4+ and NO3− fluxes of fine roots [4]. For instance, gradual increases in the fluxes of NH4+ and/or NO3− were determined when supplied N was elevated in P. popularis and P. alba × P. glandulosa [4], and C. sinensis [1]. However, the opposite results were observed in P. glauca [8], wheat [24], and corn (Zea mays L.) [33]. In our study, except for T. ascendens under NH4Cl and T. mucronatum under KNO3 solutions, most outcomes showed elevated NH4+ or NO3− uptake rates resulting from increasing NH4Cl or KNO3 supply. Consistently, higher 15N AT% and Ndff% were induced by 15NH4+ or 15NO3− treatment in the roots of T. distichum. Moreover, 15NH4+ or 15NO3− fertilization also brought about higher activities of NR, NiR, GS, GDH and GOGAT in the roots of T. distichum. These results suggest that N fertilization could be applied to stimulate NH4+ and NO3− absorption and assimilation capacities for Taxodium plants in practice.
Although an increasing N supply is likely to enhance N uptake in most cases, the provision of just NH4+ could lead to soil acidification [34]. In most cases, to maintain ion homeostasis, roots release H+ while absorbing NH4+, decreasing pH in the growth medium [35,36]. Eventually, this may lead to physiological and morphological disturbance of plants and then bring about toxicity and low production [37]. For example, acidification can significantly induce aluminum absorption, which is harmful to the development of plants [38]. In contrast, after absorption of NO3−, OH− could be released, contributing to the increase of pH [39]. Thus, a balanced supply of NH4+ and NO3− is expected to improve the N uptake of plants and the soil environment.
4.3. Net NH4+ and NO3− Fluxes under Mixed N Treatments
Many studies had demonstrated that the uptake of NH4+ and NO3− was affected by each other when both N forms were provided [1,7]. In this study, the presence of NO3− stimulated the uptake of NH4+, whereas the net fluxes of NO3− were inhibited by NH4+ in Taxodium plants. Similar results were found in the roots of corn, tea, wheat, rice and Brassica campestris [1,2,9,24,33], which indicated that NH4+ and NO3− might interact with each other under coexistence N forms. These results might be related to cytosolic NH4+/NO3− thresholds [30]. In detail, more NH4+ may be required for plant development when NO3− was provided, while pre-existing NH4+ may reduce the thresholds of NO3− in the plant [1]. Considering that a higher net NH4+ influx than NO3− was observed, it can be concluded that Taxodium plants show a preference for NH4+. It is noted that when NH4Cl or KNO3 was solely supplied, the fluxes of NH4+ or NO3− in the three species were erratic. However, stable net NH4+ and NO3− fluxes were observed when NH4+ and NO3− were both present in the solution, indicating a better balance in the mixed solution. In addition, this interesting phenomenon was reported in the study of C. sinensi, which might be the result of the competition between NH4+ and NO3−, and the underlying mechanism needs to be further studied [1].
Because of the greater N uptake in mixed treatments than in single N conditions, strong net uptake of NH4+ in fine roots of Taxodium species was expected to occur when NH4+ and NO3− were supplied in different proportions [37]. In the case of tea, the maximum net NH4+ influx was observed when NH4+:NO3− was 1:1, and the highest net NO3− influx occurred when NH4+:NO3− was 1.2:1 [1]. In blueberry (Vaccinium corymbosum L.), the mRNA levels of ammonium transporter 3 (VcAMT3) involved in NH4+ uptake as well as nitrate transporter 1.5 (VcNRT1.5) and VcNRT2 involved in NO3− uptake was highest when the NH4+:NO3− ratio was 2:1 [40]. The highest growth rate, which is positively correlated with N uptake, of T. aestivum L., Brachiaria brizantha, and Pseudostellaria heterophylla was found when the NH4+ and NO3− were supplied equivalently [1,24,41,42]. In this study, the best uptake rates of N were found when NH4+:NO3− was 3:1, 1:3, and 3:1 for T. ascendens, T. distichum, and T. mucronatum, respectively, which could provide an applicable proportion of NH4+ and NO3− when producing special N fertilizer for the productivity of Taxodium plants. Additionally, the optimal equilibrium between NH4+ and NO3− supply largely differed between the three Taxodium species, implying that the induction of N transport systems require distinct NH4+ and NO3− ratios among these plants.
In the present study, we found that with the change in the proportion of NH4+: NO3− (total N concentration: 2 mM), the NH4+ influxes were improved more than NO3−. This observation indicates a preference for NH4+ over NO3−, which is in good agreement with our previous outcomes. In most plant species, NH4+ is first absorbed into cells and then directly converted to amino acids, whereas cytosolic NO3− is assimilated at a higher energy cost. It is reduced to NO2− with the help of nitrate reductase (NR) and is further converted into NH4+ in plastids by nitrite reductase (NiR), which requires more energy than NH4+ for both transportation and further reduction [43]. On the other hand, the flux discrepancies between NH4+ and NO3− might result from the lower activity of NO3− transport systems affected by NH4+, which reduces the expression of the NO3−-related genes [1]. In blueberry plants, the expression of AMTs and NRTs was largely affected by the different ratios of NH4+: NO3− [40]. A previous study has indicated that different AMTs determined the uptake of NH4+ to a certain extent, which was mediated by the external concentration [44]. Furthermore, various AMTs and NRTs have different substrate affinities appropriate to different N concentrations [5,8,10]. Therefore, the complicated fluxes of NH4+ and NO3− when supplied at different proportions might be related to the distinct energetic and biochemical characteristics of uptake and assimilation pathway between NH4+ and NO3− in plant roots [4,5,24].
4.4. Net NH4+ and NO3− Fluxes Associated with H+
In this study, the alteration of H+ fluxes was tightly associated with the variation in NH4+ or NO3−. Previous studies revealed that H+ fluxes might be correlated with the transport of NH4+ and NO3−, since NH4+ is transported into root cells through a symporter (co-transport with H+) and/or a uniporter, and NO3− is co-transported with H+ via a symporter into the cytosol [12,13,14]. Additionally, by maintaining a proton gradient, plasma membrane PM-H+-ATPase facilitates transport by pumping H+ into the apoplast during the uptake of NH4+ or NO3− in some parts of the roots [4,43,45]. The activities of PM-H+-ATPase are determined by the transcript levels of corresponding mRNAs [4]. Although inconsistent results of H+ fluxes under different N treatments were observed in the present study, H+ still plays an essential role in plant uptake of NH4+ and NO3−. Similar to a previous study in fine roots of P. popularis [7], our data indicated a tendency for net H+ uptake when two forms of N were supplied simultaneously. Intriguingly, fluxes of H+ fluctuated under both single N sources. Through our findings, we suspect that there may be an interaction between net H+ flux and net NH4+/NO3− flux in roots of Taxodium species. Similar results were observed by Garnett et al. [15]. The specifics of the proposed interaction remain unclear. It is challenging to find out the specific mechanism underlying the correlation between H+ and NH4+/NO3− in Taxodium roots.
5. Conclusions
In summary, spatial variability of NH4+ and NO3− fluxes was observed along fine roots of Taxodium plants, and T. ascendens and T. distichum had higher fluxes of NH4+ and NO3− at 2.1–3.0 mm from the root apex. In most cases, net fluxes of NH4+ and NO3− increased with the elevated single N levels. NH4+ and NO3− affected each other when they were both supplied, and Taxodium plants preferred NH4+. Higher net N influxes were found when NH4+ and NO3− were simultaneously supplied than sole N treatments, especially in T. ascendens and T. mucronatum at 3:1 of NH4+:NO3−. Additionally, H+ fluxes were tightly correlated with net NH4+ and NO3− fluxes. These findings are valuable for understanding the characteristics of NH4+ and NO3− fluxes in the fine roots of Taxodium plants in the context of single and various ratios of N supply, and could provide a scientific basis for N management for silvicultural practice and better productivity of Taxodium plants.
Author Contributions
Conceptualization, J.H.; Formal analysis, S.W. and Y.L.; Investigation, S.W., Y.L. and R.Z.; Methodology, J.H. and Y.Y.; Resources, J.H., R.Z. and Y.Y.; Writing—original draft preparation, S.W., J.H. and Y.L.; Writing—review and editing, S.W. and J.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 31870592 and 32101490, and Jiangsu Long-term Scientific Research Base for Taxodium Rich. Breeding and Cultivation, grant number LYKJ(2021)05.
Institutional Review Board Statement
The study did not require ethical approval, for studies not involving humans or animals.
Informed Consent Statement
Studies not involving humans.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ruan, L.; Wei, K.; Wang, L.; Cheng, H.; Zhang, F.; Wu, L.; Bai, P.; Zhang, C. Characteristics of NH4+ and NO3− fluxes in tea (Camellia sinensis) roots measured by scanning ion-selective electrode technique. Sci. Rep. 2016, 6, 38370. [Google Scholar] [CrossRef] [PubMed]
- Colmer, T.D.; Bloom, A.J. A comparison of NH4+ and NO3– net fluxes along roots of rice and maize. Plant Cell Environ. 2010, 21, 240–246. [Google Scholar] [CrossRef]
- Hawkins, B.J.; Robbins, S.; Porter, R.B. Nitrogen uptake over entire root systems of tree seedlings. Tree Physiol. 2014, 34, 334–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.B. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Li, Y.; Meng, S.; Zhang, C. Net NH4+ and NO3− fluxes, and expression of NH4+ and NO3− transporter genes in roots of Populus simonii after acclimation to moderate salinity. Trees 2014, 28, 1813–1821. [Google Scholar] [CrossRef]
- Hawkins, B.J.; Boukcim, H.; Plassard, C. A comparison of ammonium, nitrate and proton net fluxes along seedling roots of Douglas-fir and lodgepole pine grown and measured with different inorganic nitrogen sources. Plant Cell Environ. 2008, 31, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Qin, J.; He, F.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.B. Net fluxes of ammonium and nitrate in association with H+ fluxes in fine roots of Populus popularis. Planta 2013, 237, 919–931. [Google Scholar] [CrossRef]
- Alber, A.; Ehlting, B.; Ehlting, J.; Hawkins, B.; Rennenberg, H. Net NH4+ and NO3− flux, and expression of NH4+ and NO3− transporters in roots of Picea glauca. Trees 2012, 26, 1403–1411. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, X.; Hao, Y.; Su, W.; Liu, H.; Sun, G.; Chen, R.; Song, S. Ammonium transporter (BcAMT1.2) mediates the interaction of ammonium and nitrate in Brassica campestris. Front. Plant Sci. 2019, 10, 1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Qi, B.; Hao, Y.; Liu, H.; Sun, G.; Chen, R.; Song, S. Appropriate NH4+/NO3− ratio triggers plant growth and nutrient uptake of optimizing the pH value of nutrient solution. Front. Plant Sci. 2021, 12, 656144. [Google Scholar] [CrossRef]
- Tang, B.; Yin, C.; Yang, H.; Sun, Y.; Liu, Q. The coupling effects of water deficit and nitrogen supply on photosynthesis, WUE, and stable isotope composition in Picea asperata. Acta Physiol. Plant 2017, 39, 148. [Google Scholar] [CrossRef]
- Zhang, M.; Ding, M.; Xu, F.; Afzal, M.R.; Chen, X.; Zeng, H.; Yan, F.; Zhu, Y. Involvement of plasma membrane H+-ATPase in the ammonium-nutrition response of barley roots. J. Plant. Nutr. Soil Sci. 2018, 181, 878–885. [Google Scholar] [CrossRef]
- McClure, P.R.; Kochian, L.V.; Spanswick, R.M.; Shaff, J.E. Evidence for cotransport of nitrate and protons in maize roots: II. Measurement of NO3− and H+ fluxes with ion-selective microelectrodes. Plant Physiol. 1990, 93, 290–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.; Di, T.; Zhu, Y.; Subbarao, G.V. Transcriptional response of plasma membrane H+-ATPase genes to ammonium nutrition and its functional link to the release of biological nitrification inhibitors from sorghum roots. Plant Soil 2016, 398, 301–312. [Google Scholar] [CrossRef]
- Garnett, T.P.; Shabala, S.N.; Smethurst, P.J.; Newman, I.A. Kinetics of ammonium and nitrate uptake by eucalypt roots and associated proton fluxes measured using ion selective microelectrodes. Funct. Plant Biol. 2003, 30, 1165–1176. [Google Scholar] [CrossRef]
- Santi, S.; Locci, G.; Pinton, R.; Cesco, S.; Varanini, Z. Plasma membrane H+-ATPase in maize roots induced for NO3− uptake. Plant Physiol. 1995, 109, 1277–1283. [Google Scholar] [CrossRef] [Green Version]
- Sorgona, A.; Lupini, A.; Mercati, F.; Di Dio, L.; Sunseri, F.; Abenavoli, M.R. Nitrate uptake along the maize primary root: An integrated physiological and molecular approach. Plant Cell Environ. 2011, 34, 1127–1140. [Google Scholar] [CrossRef]
- Stalter, R. Some ecological observations of Taxodium distichum (L.) Richard, in Delaware. Castanea 1981, 46, 154–161. [Google Scholar] [CrossRef]
- Rockwood, D.; Naidu, C.; Carter, D.; Rahmani, M.; Spriggs, T.; Lin, C.; Alker, G.; Isebrands, J.; Segrest, S. Short-rotation woody crops and phytoremediation: Opportunities for agroforestry? In New Vistas in Agroforestry; Springer: Berlin/Heidelberg, Germany, 2004; Volume 61, pp. 51–63. [Google Scholar]
- Abdelsalam, N.R.; Salem, M.Z.; Ali, H.M.; Mackled, M.I.; Mervat, E.-H.; Elshikh, M.S.; Hatamleh, A.A. Morphological, biochemical, molecular, and oil toxicity properties of Taxodium trees from different locations. Ind. Crops Prod. 2019, 139, 111515. [Google Scholar] [CrossRef]
- Zaher, A.M.; Lin, J.; Arai, M. Cytotoxic activity of abietane-type diterpenes isolated from Taxodium distichum against cancer cells adapted to nutrient-starved conditions. Nat. Prod. Commun. 2020, 15, 1–6. [Google Scholar] [CrossRef]
- Hua, J.; Han, L.; Wang, Z.; Gu, C.; Yin, Y. Morpho-anatomical and photosynthetic responses of Taxodium hybrid’Zhongshanshan’406 to prolonged flooding. Flora 2017, 231, 29–37. [Google Scholar] [CrossRef]
- Xuan, L.; Hua, J.; Zhang, F.; Wang, Z.; Pei, X.; Yang, Y.; Yin, Y.; Creech, D.L. Identification and functional analysis of ThADH1 and ThADH4 genes involved in tolerance to waterlogging stress in Taxodium hybrid ‘Zhongshanshan 406’. Genes 2021, 12, 225. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Yan, W.; Chen, J.; Shangguan, Z. Net ammonium and nitrate fluxes in wheat roots under different environmental conditions as assessed by scanning ion-selective electrode technique. Sci. Rep. 2014, 4, 7223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Hu, R.; Zheng, Z.; Yang, J.; Fan, H.; Deng, X.; Yao, W.; Wang, Q.; Peng, S.; Li, J. Soil bacterial community in the multiple cropping system increased grain yield within 40 cultivation years. Front. Plant Sci. 2021, 12, 804527. [Google Scholar] [CrossRef] [PubMed]
- Enstone, D.E.; Peterson, C.A.; Hallgren, S.W. Anatomy of seedling tap roots of loblolly pine (Pinus taeda L.). Trees 2001, 15, 98–111. [Google Scholar] [CrossRef]
- Fang, Y.Y.; Babourina, O.; Rengel, Z.; Yang, X.E.; Pu, P.M. Spatial distribution of ammonium and nitrate fluxes along roots of wetland plants. Plant Sci. 2007, 173, 240–246. [Google Scholar] [CrossRef]
- Li, Q.; Li, B.H.; Kronzucker, H.J.; Shi, W.M. Root growth inhibition by NH4+ in Arabidopsis is mediated by the root tip and is linked to NH4+ efflux and GMPase activity. Plant Cell Environ. 2010, 33, 1529–1542. [Google Scholar] [CrossRef]
- Tang, B.; Yin, C.; Liu, Q. Characteristics of ammonium and nitrate fluxes along the roots of Picea asperata. J. Plant Nutr. Soil Sci. 2019, 42, 772–782. [Google Scholar] [CrossRef]
- Zou, N.; Shi, W.; Hou, L.; Kronzucker, H.J.; Huang, L.; Gu, H.; Yang, Q.; Deng, G.; Yang, G. Superior growth, N uptake and NH4+ tolerance in the giant bamboo Phyllostachys edulis over the broad-leaved tree Castanopsis fargesii at elevated NH4+ may underlie community succession and favor the expansion of bamboo. Tree Physiol. 2020, 40, 1606–1622. [Google Scholar] [CrossRef]
- Lu, Y.; Deng, S.; Li, Z.; Wu, J.; Zhu, D.; Shi, W.; Zhou, J.; Fayyaz, P.; Luo, Z.B. Physiological characteristics and transcriptomic dissection in two root segments with contrasting net fluxes of ammonium and nitrate of poplar under low nitrogen availability. Plant Cell Physiol. 2022, 63, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Di, D.W.; Sun, L.; Wang, M.; Wu, J.; Kronzucker, H.J.; Fang, S.; Chu, J.; Shi, W.; Li, G. WRKY46 promotes ammonium tolerance in Arabidopsis by repressing NUDX9 and indole-3-acetic acid-conjugating genes and by inhibiting ammonium efflux in the root elongation zone. New Phytol. 2021, 232, 190–207. [Google Scholar] [CrossRef] [PubMed]
- Mackown, C.T.; Jackson, W.A.; Volk, R.J. Restricted nitrate influx and reduction in corn seedlings exposed to ammonium. Plant Physiol. 1982, 69, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.; Gerendás, J.; Härdter, R.; Sattelmacher, B. Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis) plants. Ann. Bot. 2007, 99, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinsinger, P.; Plassard, C.; Tang, C.; Jaillard, B. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: A review. Plant Soil 2003, 248, 43–59. [Google Scholar] [CrossRef]
- Tang, C.; Drevon, J.; Jaillard, B.; Souche, G.; Hinsinger, P. Proton release of two genotypes of bean (Phaseolus vulgaris L.) as affected by N nutrition and P deficiency. Plant Soil 2004, 260, 59–68. [Google Scholar] [CrossRef]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.; Ma, L.; Shi, Y.; Zhang, F. Effects of litter incorporation and nitrogen fertilization on the contents of extractable aluminium in the rhizosphere soil of tea plant (Camallia sinensis (L.) O. Kuntze). Plant Soil 2004, 263, 283–296. [Google Scholar] [CrossRef]
- Bar-Yosef, B.; Mattson, N.; Lieth, H. Effects of NH4+:NO3−:Urea ratio on cut roses yield, leaf nutrients content and proton efflux by roots in closed hydroponic system. Sci. Hortic. 2009, 122, 610–619. [Google Scholar] [CrossRef]
- Xu, J.; Fang, Y.; Tavakkoli, E.; Pan, X.; Liao, F.; Chen, W.; Guo, W. Preferential ammonium: Nitrate ratio of blueberry isregulated by nitrogen transport and reduction systems. Sci. Hortic. 2021, 288, 110345. [Google Scholar] [CrossRef]
- De Bona, F.D.; Schmidt, F.; Monteiro, F.A. Importance of the nitrogen source in the grass species Brachiaria brizantha responses to sulfur limitation. Plant Soil 2013, 373, 201–216. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.; Liang, Y.; Gao, W.; Wang, J.; Jing, S.; Zhang, Y.; Liu, H. Influence of medium salt strength and nitrogen source on biomass and metabolite accumulation in adventitious root cultures of Pseudostellaria heterophylla. Acta Physiol. Plant 2013, 35, 2623–2628. [Google Scholar] [CrossRef]
- Miller, A.; Cramer, M. Root nitrogen acquisition and assimilation. Plant Soil 2005, 274, 1–36. [Google Scholar] [CrossRef]
- Duan, F.; Giehl, R.F.H.; Geldner, N.; Salt, D.E.; von Wiren, N. Root zone-specific localization of AMTs determines ammonium transport pathways and nitrogen allocation to shoots. PLoS Biol. 2018, 16, e2006024. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. Futile cycling at the plasma membrane: A hallmark of low-affinity nutrient transport. Trends Plant Sci. 2006, 11, 529–534. [Google Scholar] [CrossRef]
Figure 1.
Root tip in NMT experiment (a) and the vertical section in the root of T. distichum (b).

Figure 2.
Net NH4+ (a) and NO3− (b) fluxes along the root tip of T. ascendens, T. distichum, and T. mucronatum. Bars indicate standard errors (n = 8). Net influxes correspond to positive values, and negative values indicate net effluxes, respectively. The concentrations of N were set as 1.0 mM NH4Cl for NH4+ or 1.0 mM KNO3 for NO3−. T. a, T. d, and T. m represent T. ascendens, T. distichum and T. mucronatum, respectively.
Figure 2.
Net NH4+ (a) and NO3− (b) fluxes along the root tip of T. ascendens, T. distichum, and T. mucronatum. Bars indicate standard errors (n = 8). Net influxes correspond to positive values, and negative values indicate net effluxes, respectively. The concentrations of N were set as 1.0 mM NH4Cl for NH4+ or 1.0 mM KNO3 for NO3−. T. a, T. d, and T. m represent T. ascendens, T. distichum and T. mucronatum, respectively.

Figure 3.
Net fluxes of NH4+ and NO3− under single (a,d,g) and mixed (b,e,h) N forms, and the means of net fluxes of NH4+ and NO3− (c,f,i). Bars indicate standard errors (n = 8). Different letters indicate significant differences among the treatments according to Duncan’s Multiple Range Test at 5% level. T. a, T. d, and T. m represent T. ascendens, T. distichum and T. mucronatum, respectively.
Figure 3.
Net fluxes of NH4+ and NO3− under single (a,d,g) and mixed (b,e,h) N forms, and the means of net fluxes of NH4+ and NO3− (c,f,i). Bars indicate standard errors (n = 8). Different letters indicate significant differences among the treatments according to Duncan’s Multiple Range Test at 5% level. T. a, T. d, and T. m represent T. ascendens, T. distichum and T. mucronatum, respectively.

Figure 4.
Net fluxes of NH4+ (a), NO3− (b) and H+ (c) under different concentrations of NH4Cl and KNO3. Bars indicate standard errors (n = 8). Different letters indicate significant differences among the treatments according to Duncan’s Multiple Range Test at 5% level. T. a, T. d, and T. m represent T. ascendens, T. distichum and T. mucronatum, respectively.
Figure 4.
Net fluxes of NH4+ (a), NO3− (b) and H+ (c) under different concentrations of NH4Cl and KNO3. Bars indicate standard errors (n = 8). Different letters indicate significant differences among the treatments according to Duncan’s Multiple Range Test at 5% level. T. a, T. d, and T. m represent T. ascendens, T. distichum and T. mucronatum, respectively.

Figure 5.
Net fluxes of NH4+ (a), NO3− (b), total N (c) and H+ (d) under different proportions of NH4Cl and KNO3. Bars indicate standard errors (n = 8). Different letters indicate significant differences among the treatments according to Duncan’s Multiple Range Test at 5% level. T. a, T. d, and T. m represent T. ascendens, T. distichum and T. mucronatum, respectively.
Figure 5.
Net fluxes of NH4+ (a), NO3− (b), total N (c) and H+ (d) under different proportions of NH4Cl and KNO3. Bars indicate standard errors (n = 8). Different letters indicate significant differences among the treatments according to Duncan’s Multiple Range Test at 5% level. T. a, T. d, and T. m represent T. ascendens, T. distichum and T. mucronatum, respectively.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biomass, 15N AT% and Ndff% in the roots of T. distichum under three N treatments.
| Treatments | Root Biomass/g | 15N AT% | Ndff% |
|---|---|---|---|
| CK | 0.30 ± 0.01 bc | 0.37 ± 0.00 c | / |
| 1 mM 15NH4+ | 0.34 ± 0.02 ab | 0.63 ± 0.03 a | 2.65 ± 0.31 a |
| 1 mM 15NO3− | 0.26 ± 0.01 c | 0.48 ± 0.04 b | 1.16 ± 0.38 b |
Different letters behind the values in the same column indicate significant differences among the treatments according to Duncan’s Multiple Range Test at 5% level. 15N AT%: 15N Atom; Ndff%:15N from N source.
Table 2.
Activities of N assimilation enzymes in the roots of T. distichum under three N treatments.
Table 2.
Activities of N assimilation enzymes in the roots of T. distichum under three N treatments.
| Treatments | NR Activity μmol h−1 mg−1 Protein | NiR Activity μmol h−1 mg−1 Protein | GS Activity μmol h−1 mg−1 Protein | GDH Activity μmol h−1 mg−1 Protein | GOGAT Activity μmol h−1 mg−1 Protein |
|---|---|---|---|---|---|
| CK | 0.58 ± 0.02 c | 2.43 ± 0.04 c | 0.33 ± 0.00 b | 1.04 ± 0.06 b | 0.78 ± 0.03 b |
| 1 mM 15NH4+ | 0.87 ± 0.02 b | 3.25 ± 0.08 a | 0.46 ± 0.22 a | 1.54 ± 0.08 a | 1.24 ± 0.03 a |
| 1 mM 15NO3− | 1.92 ± 0.04 a | 2.72 ± 0.01 b | 0.34 ± 0.01 b | 1.01 ± 0.04 b | 0.74 ± 0.01 b |
Different letters behind the values in the same column indicate significant differences between the treatments according to Duncan’s Multiple Range Test at 5% level. NR: nitrate reductase; NiR: nitrite reductase; GS: glutamine synthetase; GDH: glutamate dehydrogenase; GOGAT: glutamate synthetase.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, S.; Hua, J.; Lu, Y.; Zhang, R.; Yin, Y. Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments. Plants 2022, 11, 894. https://doi.org/10.3390/plants11070894
AMA Style
Wu S, Hua J, Lu Y, Zhang R, Yin Y. Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments. Plants. 2022; 11(7):894. https://doi.org/10.3390/plants11070894
Chicago/Turabian StyleWu, Shuting, Jianfeng Hua, Yan Lu, Rui Zhang, and Yunlong Yin. 2022. "Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments" Plants 11, no. 7: 894. https://doi.org/10.3390/plants11070894
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.