
Effects of Four Cropping Patterns of Lilium brownii on Rhizosphere Microbiome Structure and Replant Disease


Abstract
:1. Introduction
2. Results
2.1. Changes in Soil pH under Four Planting Patterns
2.2. Overview of 16S rDNA Pyrosequencing and ITS rDNA Sequence Pyrosequencing
2.3. Bacterial and Fundal α-Diversity Analysis
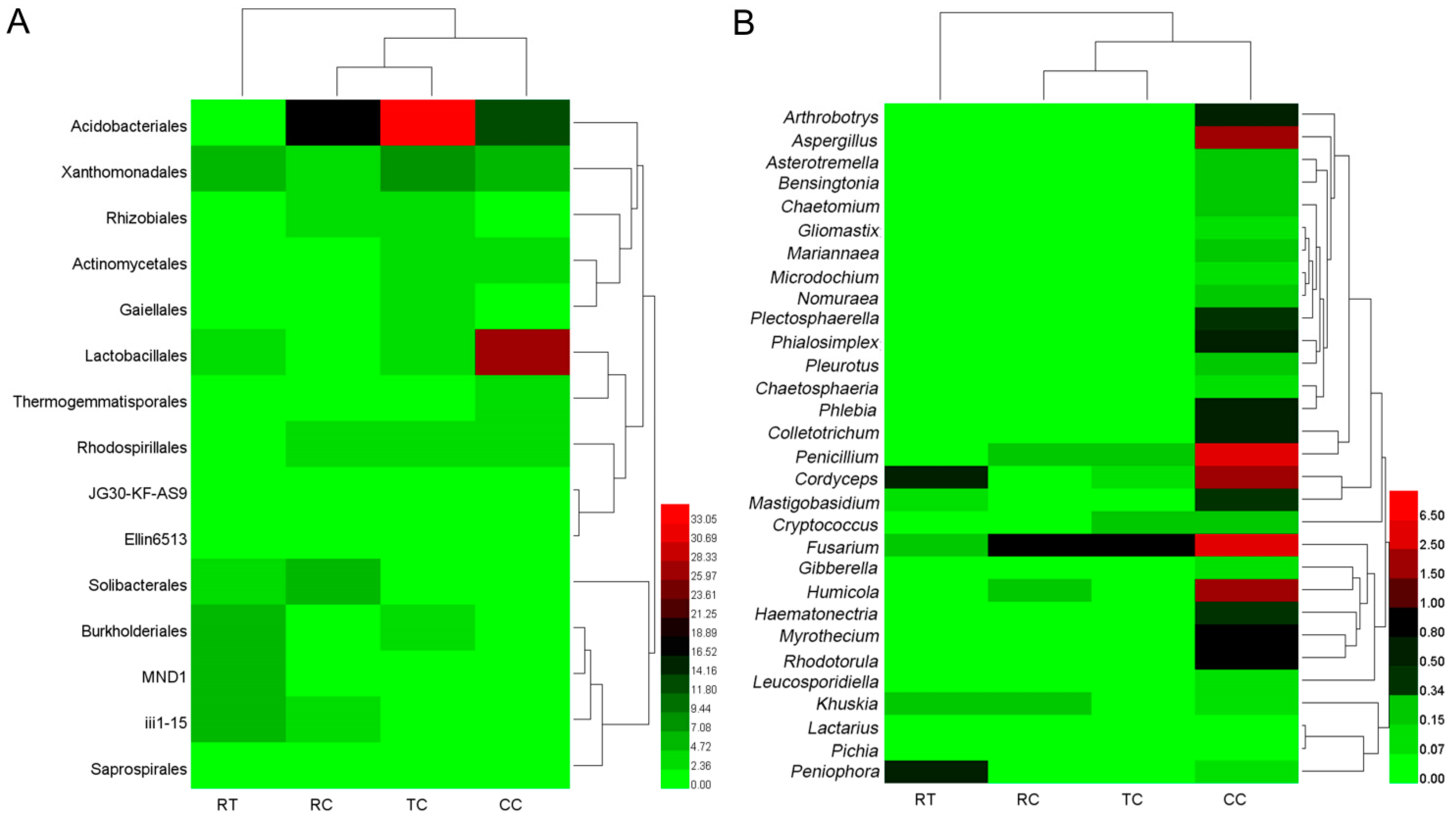
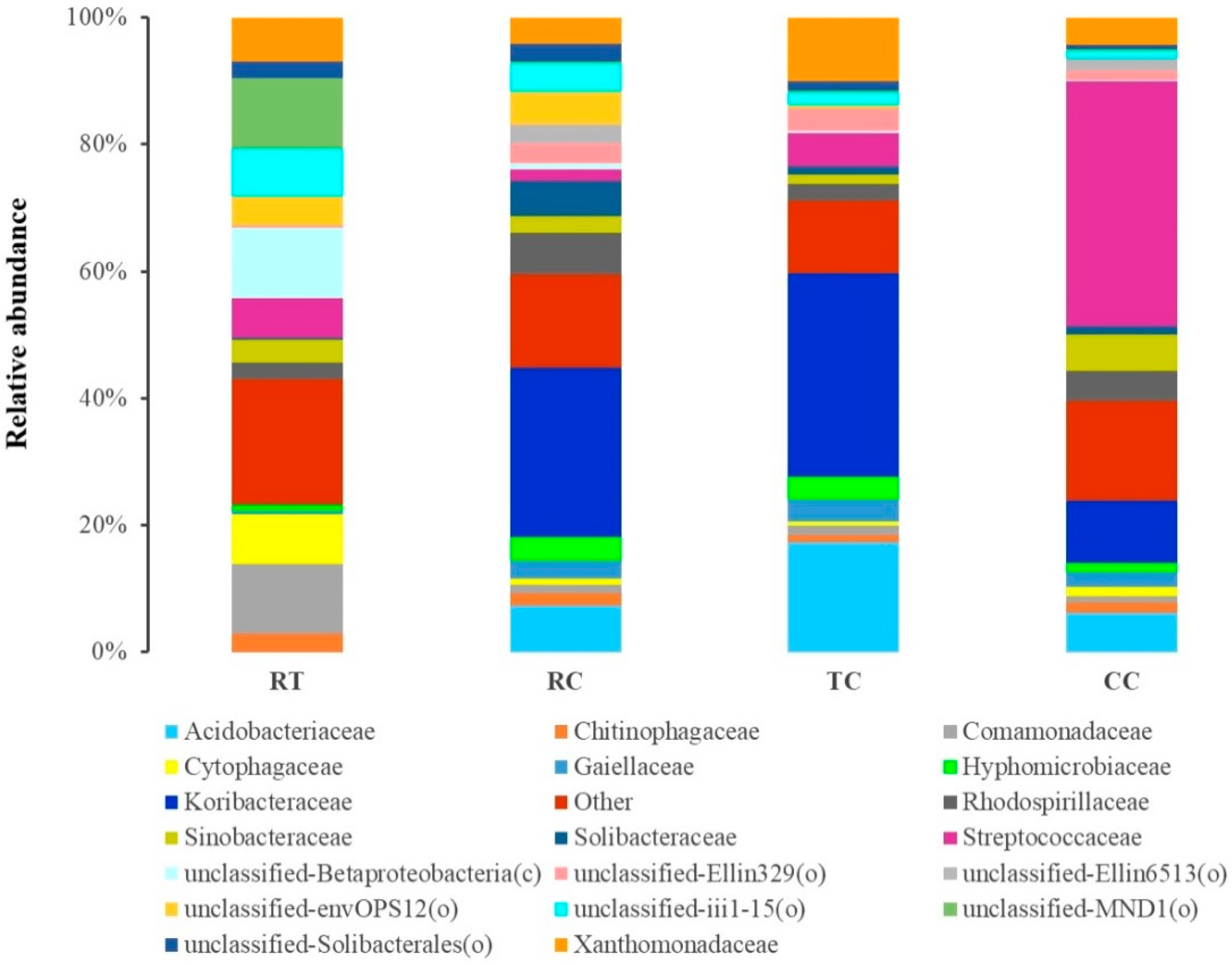
2.4. Shift in Rhizosphere Bacterial and Fungal Populations under Different Planting Patterns
3. Discussion
3.1. The Correlation between Soil pH and Rhizosphere Bacterial Diversity
3.2. Changes in Microbial Composition under Four Plant Treatments
3.3. Effects of Paddy-Upland Rotation
4. Materials and Methods
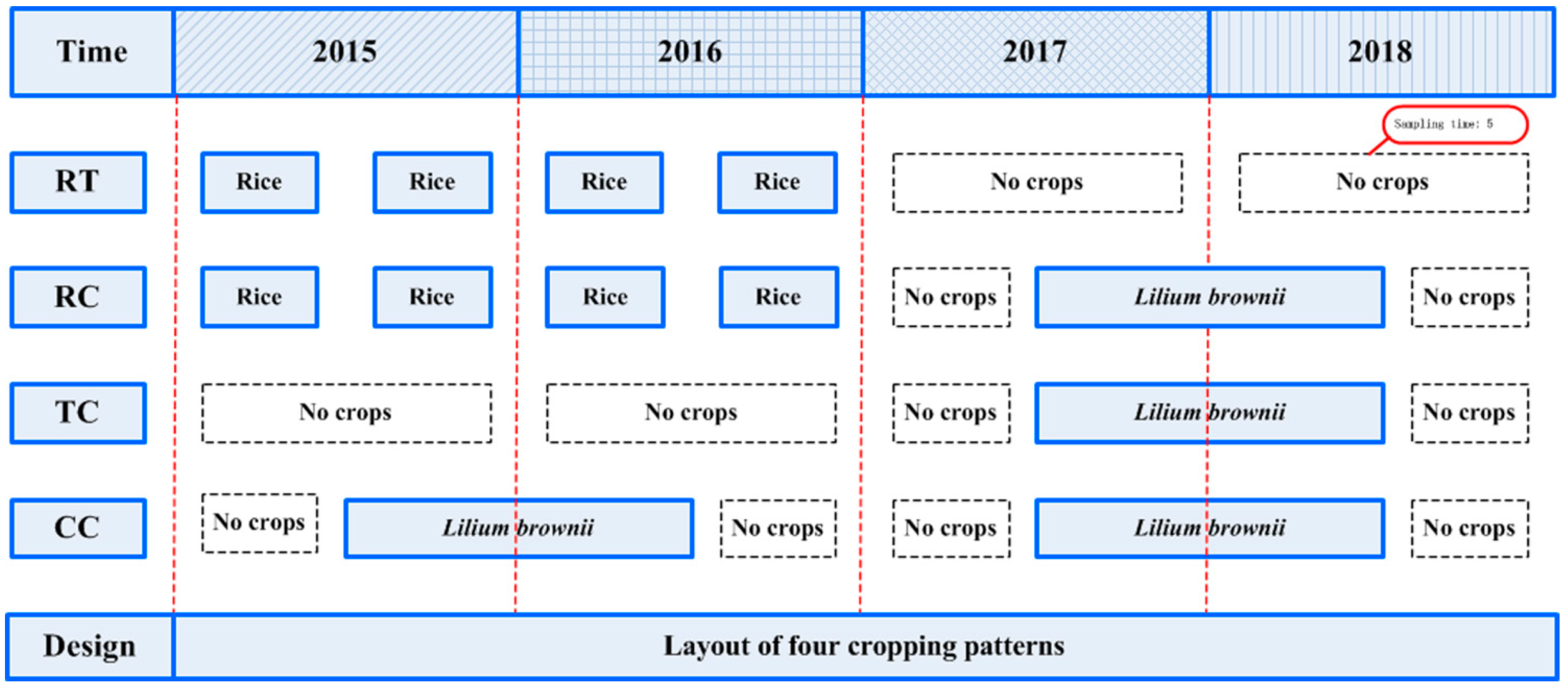
4.1. Overview of the Experimental Site
4.2. Experimental Design
4.3. Sample Preparation and Determination of Soil pH
4.4. Extraction and Pyrosequencing of Soil Total DNA
4.5. Analysis of Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Louis, B.P.; Maron, P.A.; Viaud, V.; Leterme, P.; Menasseri-Aubry, S. Soil C and N models that integrate microbial diversity. Environ. Chem. Lett. 2016, 14, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, C.R. Role of microbial diversity for soil, health and plant nutrition. In Molecular Mechanisms of Plant and Microbe Coexistence; Springer: Berlin/Heidelberg, Germany, 2008; Volume 15, pp. 53–74. [Google Scholar]
- Chourasiya, D.; Sharma, M.P.; Maheshwari, H.S.; Ramesh, A.; Sharma, S.K.; Adhya, T.K. Microbial Diversity and Soil Health in Tropical Agroecosystems. Microorg. Sustain. 2018, 10, 978–981. [Google Scholar]
- Ryszkowski, L.; Szajdak, L.; Karg, J. Effects of continuous cropping of rye on soil biota and biochemistry. Crit. Rev. Plant Sci. 1998, 17, 225–244. [Google Scholar] [CrossRef]
- She, S.Y.; Niu, J.J.; Zhang, C.; Xiao, Y.H.; Chen, W.; Dai, L.J.; Liu, X.D.; Yin, H.Q. Significant relationship between soil bacterial community structure and incidence of bacterial wilt disease under continuous cropping system. Arch. Microbiol. 2017, 199, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.X.; Luo, J.G.; Guo, C.; Kong, L.Y. New steroidal saponins from the bulbs of L. brownii. Carbohydr. Res. 2012, 361, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Wang, Z.; Zhang, Y.M.; Luo, J.G.; Kong, L.Y. Bioassay-Guided Isolation of Anti-Inflammatory Components from the Bulbs of L.brownii and Identifying the Underlying Mechanism through Acting on the NF-kappa B/MAPKs Pathway. Molecules 2017, 22, 506. [Google Scholar] [CrossRef] [Green Version]
- Tong, Q.Z. Studies on the Germplasm Resources Evaluation and Utilization on Medical Lily in Hunan. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2009. [Google Scholar]
- Zeng, L. Current Production Situation of Longya Lilium in Shaoyang and Studies on Its In Vitro Buld Induction as well as Detoxification. Master’s Thesis, Hunan Agricultural University, Changsha, China, 2010. [Google Scholar]
- Song, J.T.; Lin, Z.J.; Zhang, J.X.; Meng, J.; Chen, Z.B. Geochemical Characteristics of Selenium in Soil and Its Influencing Factors in Longhui County, Hunan Province: A Case Study of Shimen-Tantou Township. South China Geol. 2021, 1, 83–92. [Google Scholar]
- Li, Y.F.; Ming, J.; Liu, X.Y.; Wang, L.G.; Yuan, S.X.; Liu, C.; Wang, Y.; Xu, L.F.; Yuan, Y.Y. Comparative Study on Edibility of 15 Lily Species and Varieties. Acta Hortic. Sin. 2013, 40(S), 2693. [Google Scholar]
- Gao, Z.; Wang, X.L.; Wang, B.; Li, R.G. Effects of fertilization on the non-virus bulb cultivation of Lilium brownii var. viridulum. China Cucurbits Veg. 2020, 11, 56–60. [Google Scholar]
- Benitez, M.S.; Osborne, S.L.; Lehman, R.M. Previous crop and rotation history effects on maize seedling health and associated rhizosphere microbiome. Sci. Rep. 2017, 7, 15709. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Gao, P.; Wang, Y.; Li, W.; Cui, X.W.; Zhou, J.M.; Peng, F.Y.; Dai, L.Y. Earthworm activity optimized the rhizosphere bacteal community structure and further alleviated the yield loss in continuous cropping lily (Lilium lancifolium Thunb.). Sci. Rep. 2021, 11, 20840. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.; Liu, Y.J.; Peng, S.G.; Yin, H.Q.; Meng, D.L.; Tao, J.M.; Gu, Y.B.; Li, J.; Yang, S.; Xiao, N.W.; et al. Soil potentials to resist continuous cropping obstacle: Three field cases. Environ. Res. 2021, 200, 111319. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Xue, L.; Ren, H.D.; Li, S.; Leng, X.H.; Yao, H.X. Soil bacterial community structure and co-occurrence patternduring vegetation restoration in Karst Rocky desertification area. Front. Microbiol. 2017, 8, 2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, D.S.; Teixeira, P.J. Root-exuded coumarin shapes the root microbiome. Proc. Natl. Acad. Sci. USA 2018, 115, 5629–5631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acuna, J.J.; Jorquera, M.A. Diversity, interaction, and bioprospecting of plant-associated microbiomes diversity. Diverity 2020, 10, 390. [Google Scholar]
- Du, Q.; Lu, D.; Ma, K. Effect of potato continuous cropping on soil microbial community structure and function. Ecol. Environ. Sci. 2012, 21, 1252–1256. [Google Scholar]
- Yang, Q.X.; Wang, R.F.; Xu, Y.Y.; Kang, C.X.; Miao, Y.; Li, M.J. Dynamic change of the rhizosphere microbial community in response to growth stages of consecutively monocultured Rehmanniae glutinosa. Biologia 2016, 12, 1320–1329. [Google Scholar] [CrossRef]
- Wu, J.P.; Jiao, Z.B.; Zhou, J.; Guo, F.L.; Ding, Z.L.; Qiu, Z.M. Analysis of bacterial communities in rhizosphere soil of continuously cropped healthy and diseased konjac. World J. Microbiol. Biotechnol. 2017, 33, 134. [Google Scholar] [CrossRef]
- Wei, Z.; Yu, D. Analysis of the succession of structure of the bacteria community in soil from long-term continuous cotton cropping in Xinjiang using high-throughput sequencing. Arch. Microbiol. 2018, 200, 653–662. [Google Scholar] [CrossRef]
- Li, H.; Wang, J.Q.; Liu, Q.; Zhou, Z.F.; Chen, F.L.; Xiang, D. Effects of consecutive monoculture of sweet potato on soil bacterial community as determined by pyrosequencing. J. Basic Microbiol. 2019, 2, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.L.; Xu, Y.; Yin, Y.L.; Ye, X.L. Effects of different years continuous cropping and grafting on the biological activities of eggplant soil. Chin. J. Appl. Ecol. 2010, 29, 290–294. [Google Scholar]
- Lin, M.Z.; Wang, H.B.; Lin, H.F. Effects of Pseudostellariae heterophylla continuous cropping on rhizosphere soil microorganisms. Chin. J. Appl. Ecol. 2012, 31, 106–111. [Google Scholar]
- Rousk, J.; Baath, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 10, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Delgado-Baquerizo, M.; Trivedi, C.; Hamonts, K.; Anderson, I.C.; Singh, B.K. Keystone microbial taxa regulate the invasion of a fungal pathogen in agro-ecosystems. Soil Biol. Biochem. 2017, 111, 10–14. [Google Scholar] [CrossRef]
- Cha, J.Y.; Han, S.; Hong, H.J.; Cho, H.; Kim, D.; Kwon, Y.; Kwon, S.K.; Crusemann, M.; Lee, Y.B.; Kim, J.F.; et al. Microbial and biochemical basis of a Fusarium wilt-suppressive soil. ISME J. 2016, 1, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogen: Looking back over 30 years. Phytopathology 2007, 2, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Trivedi, P.; Kumar, B.; Palni, L.; Man, S. Characterization of a phosphate solubilizing and antagonistic strain of Pseudomonas putida (B0) isolated from a sub-alpine location in the Indian Central Himalaya. Curr. Microbiol. 2006, 2, 102–107. [Google Scholar] [CrossRef]
- Jiang, J.H.; Song, Z.; Yang, X.T.; Mao, Z.Q.; Nie, X.H.; Guo, H.; Peng, X.W. Microbial community analysis of apple rhizosphere around Bohai Gulf. Sci. Rep. 2017, 7, 8918. [Google Scholar] [CrossRef]
- Wu, L.K.; Chen, J.; Xiao, Z.G.; Zhu, X.C.; Wang, J.Y.; Wu, H.M.; Wu, Y.H.; Zhang, Z.Y.; Lin, W.X. Barcoded Pyrosequencing Reveals a Shift in the Bacterial Community in the Rhizosphere and Rhizoplane of Rehmannia glutinosa under Consecutive Monoculture. Int. J. Mol. Sci. 2018, 19, 850. [Google Scholar] [CrossRef] [Green Version]
- Campos, S.B.; Lisboa, B.B.; Camargo, F.; Bayer, C.; Sczyrba, A.; Dirksen, P.; Albersmeier, A.; Kalinowski, J.; Beneduzi, A.; Costa, P.B. Soil suppressiveness and its relations with the microbial community in a Brazilian subtropical agroecosystem under different management systems. Soil Biol. Biochem. 2016, 96, 191–197. [Google Scholar] [CrossRef]
- Latz, E.; Eisenhauer, N.; Rall, B.C.; Allan, E.; Roscher, C.; Scheu, S.; Jousset, A. Plant diversity improves protection against soil-borne pathogens by fostering antagonistic bacterial communities. J. Ecol. 2012, 3, 597–604. [Google Scholar] [CrossRef]
- Li, X.G.; de Boer, W.; Zhang, Y.A.; Ding, C.F.; Zhang, T.L.; Wang, X.X. Suppression of soil-borne Fusariwn pathogens of peanut by intercropping with the medicinal herb Atractylodes lancea. Soil Biol. Biochem. 2018, 116, 120–130. [Google Scholar] [CrossRef]
- Peralta, A.L.; Sun, Y.M.; McDaniel, M.D.; Lennon, J.T. Crop rotational diversity increases disease suppressive capacity of soil microbiomes. Ecosphere 2018, 5, e02235. [Google Scholar] [CrossRef]
- Zhou, X.G.; Liu, J.; Wu, F.Z. Soil microbial communities in cucumber monoculture and rotation systems and their feedback effects on cucumber seedling growth. Plant Soil 2017, 415, 507–520. [Google Scholar] [CrossRef]
- Xi, H.; Zhang, X.K.; Qu, Z. Effects of cotton–maize rotation on soil microbiome structure. Mol. Plant Pathol. 2021, 6, 673–682. [Google Scholar] [CrossRef]
- Lopes, A.R.; Manaia, C.M.; Nunes, O.C. Bacterial community variations in an alfalfa- rice rotation system revealed by 16S rRNA gene 454-pyrosequencing. FEMS Microbiol. Ecol. 2014, 3, 650–663. [Google Scholar] [CrossRef] [Green Version]
- Hou, P.F.; Chien, C.H.; Chiang-Hsieh, Y.F.; Tseng, K.C.; Chow, C.N.; Huang, H.J.; Chang, W.C. Paddy-upland rotation for sustainable agriculture with regards to diverse soil microbial community. Sci. Rep. 2018, 8, 7966. [Google Scholar] [CrossRef]
- Maarastawi, S.A.; Frindte, K.; Linnartz, M.; Knief, C. Crop Rotation and Straw Application Impact Microbial Communities in Italian and Philippine Soils and the Rhizosphere of Zea mays. Front. Microbiol. 2018, 9, 1295. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.B.; He, Y.S.; Qu, W.H.; Pan, Q.P.; Luo, Y.L. Analysis of Lily planting base in Longhui County of Hunan province. J. Hunan Univ. Chin. Med. 2003, 3, 57–58. [Google Scholar]
- Lu, R.K. Analytical Methods of Soil Agricultural Chemistry; China Agricultural Science Press: Beijing, China, 1999. [Google Scholar]
- Zhou, J.; Bruns, M.A.; Tiedje, J.M. DNA recovery from soils of diverse composition. Appl. Environ. Microbiol. 1996, 62, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Li, M.; Wei, G.; Tian, R.; Li, C.; Wang, B.; Lin, R.; Shi, C.; Chi, X.; Zhou, B.; et al. The occurrence of potato common scab correlates with the community composition and function of the geocaulosphere soil microbiome. Microbiome 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reazin, C.; Morris, S.; Smith, J.E.; Cowan, A.D.; Jumpponen, A. Fires of differing intensities rapidly select distinct soil fungal communities in a Northwest US ponderosa pine forest ecosystem. For. Ecol. Manag. 2016, 377, 118–127. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 16, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.Z.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 13, 1658–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 7, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatment | pH | ||
|---|---|---|---|
| Seedling Emergence Period | Flourishing Period | (Bulblet) Expanding Stage | |
| RT | 6.69 ± 0.03 a | 6.47 ± 0.05 b | 6.53 ± 0.02 b |
| RC | 6.50 ± 0.16 b | 5.91 ± 0.05 c | 5.65 ± 0.11 d |
| TC | 5.25 ± 0.11 e | 4.65 ± 0.07 g | 4.61 ± 0.07 g |
| CC | 4.91 ± 0.04 f | 4.40 ± 0.03 h | 4.33 ± 0.07 h |
| Sources of Variation | Sum of Squares | Degrees of Freedom | Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| A between factors | 8.8856 | 3 | 2.9619 | 491.111 | 0.0001 |
| B between factors | 0.0055 | 2 | 0.0028 | 0.457 | 0.6386 |
| A × B | 18.4177 | 6 | 3.0696 | 508.978 | 0.0001 |
| Error | 0.1447 | 24 | 0.006 | ||
| Total variation | 27.4535 | 35 |
| Treatment | Bacteria | Fungi | ||
|---|---|---|---|---|
| Chao1 | Shonnon | Chao1 | Shonnon | |
| RT | 4867.00 ± 4.58 b | 9.95 ± 0.03 a | 230.50 ± 0.79 b | 4.53 ± 0.04 b |
| RC | 4892.00 ± 2.65 a | 9.90 ± 0.08 a | 296.47 ± 3.61 a | 3.83 ± 0.21 c |
| TC | 4018.33 ± 2.52 c | 8.53 ± 0.04 b | 155.90 ± 3.32 c | 1.96 ± 0.13 d |
| CC | 3475.00 ± 1.73 d | 7.65 ± 0.03 c | 291.73 ± 3.05 a | 6.02 ± 0.23 a |
| Genera | RT | RC | TC | CC |
|---|---|---|---|---|
| Arthrobotrys | 0 | 0.01 ± 0.002 e | 0 | 0.65 ± 0.030 g |
| Aspergillus | 0 | 0 | 0.01 ± 0.002 d | 1.91 ± 0.080 d |
| Colletotrichum | 0 | 0 | 0.02 ± 0.005 d | 0.54 ± 0.020 h |
| Cordyceps | 0.78 ± 0.040 a | 0.03 ± 0.005 d | 0.13 ± 0.026 c | 2.34 ± 0.078 c |
| Fusarium | 0.16 ± 0.020 c | 0.87 ± 0.044 a | 0.87 ± 0.020 a | 2.76 ± 0.095 b |
| Humicola | 0 | 0.22 ± 0.035 b | 0 | 1.80 ± 0.044 d |
| Myrothecium | 0.02 ± 0.002 d | 0.01 ± 0.001 e | 0 | 0.84 ± 0.046 f |
| Penicillium | 0 | 0.15 ± 0.020 c | 0.27 ± 0.055 b | 6.11 ± 0.078 a |
| Peniophora | 0.53 ± 0.030 b | 0 | 0 | 0.12 ± 0.017 i |
| Phialosimplex | 0 | 0 | 0 | 0.64 ± 0.017 g |
| Phlebia | 0 | 0 | 0 | 0.56 ± 0.017 h |
| Rhodotorula | 0 | 0.03 ± 0.001 d | 0 | 0.93 ± 0.010 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, W.; Liao, X.; Wang, C.; Zhang, Y. Effects of Four Cropping Patterns of Lilium brownii on Rhizosphere Microbiome Structure and Replant Disease. Plants 2022, 11, 824. https://doi.org/10.3390/plants11060824
Ma W, Liao X, Wang C, Zhang Y. Effects of Four Cropping Patterns of Lilium brownii on Rhizosphere Microbiome Structure and Replant Disease. Plants. 2022; 11(6):824. https://doi.org/10.3390/plants11060824
Chicago/Turabian StyleMa, Wenyue, Xiaolan Liao, Chong Wang, and Ya Zhang. 2022. "Effects of Four Cropping Patterns of Lilium brownii on Rhizosphere Microbiome Structure and Replant Disease" Plants 11, no. 6: 824. https://doi.org/10.3390/plants11060824