
Chili Pepper AN2 (CaAN2): A Visible Selection Marker for Nondestructive Monitoring of Transgenic Plants


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
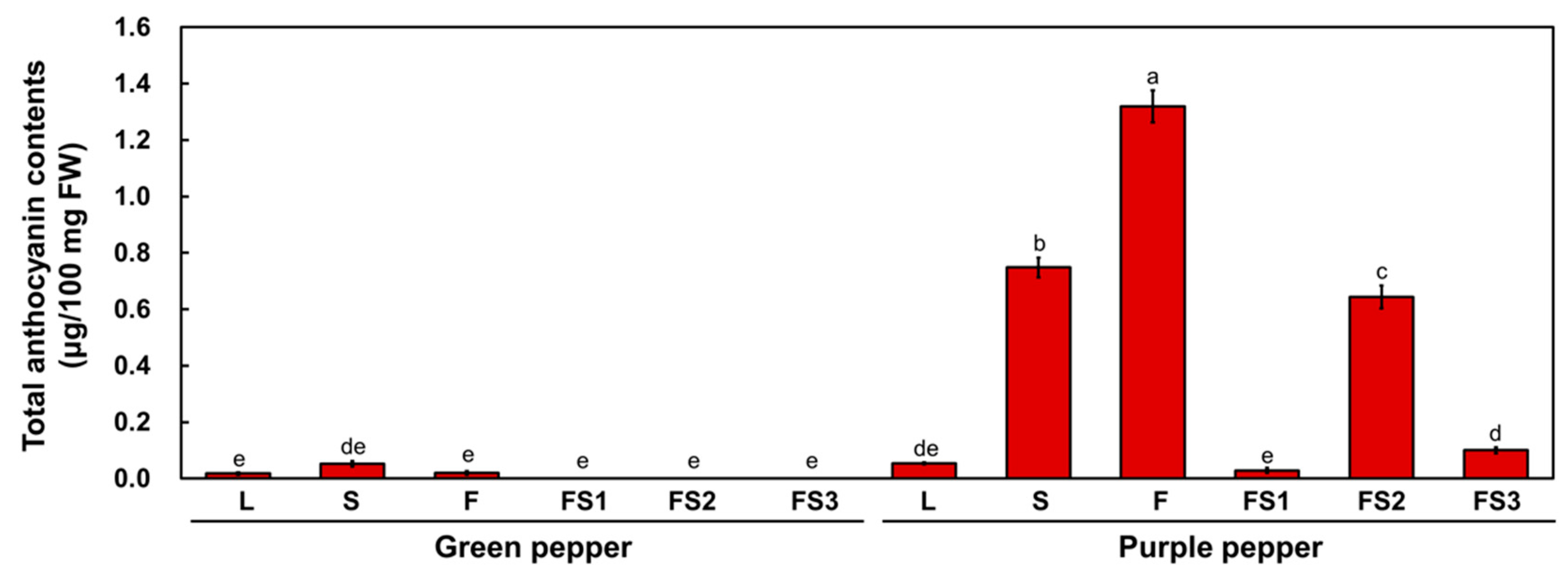
2.1. Anthocyanin Accumulation Determines the Green and Purple Coloration of Chili Pepper
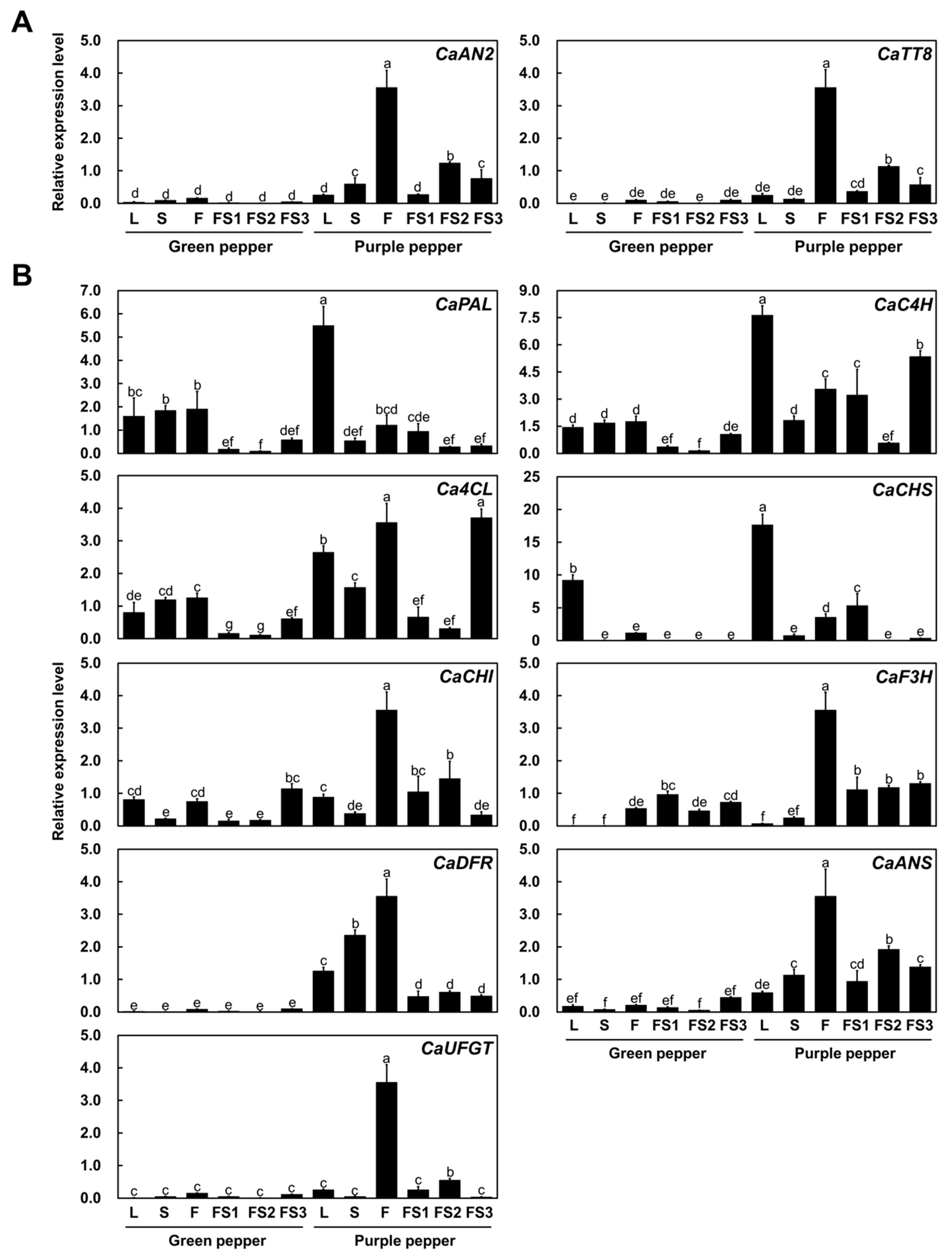
2.2. CaAN2 and CaTT8 Are Highly Expressed in Purple Chili Pepper Fruit

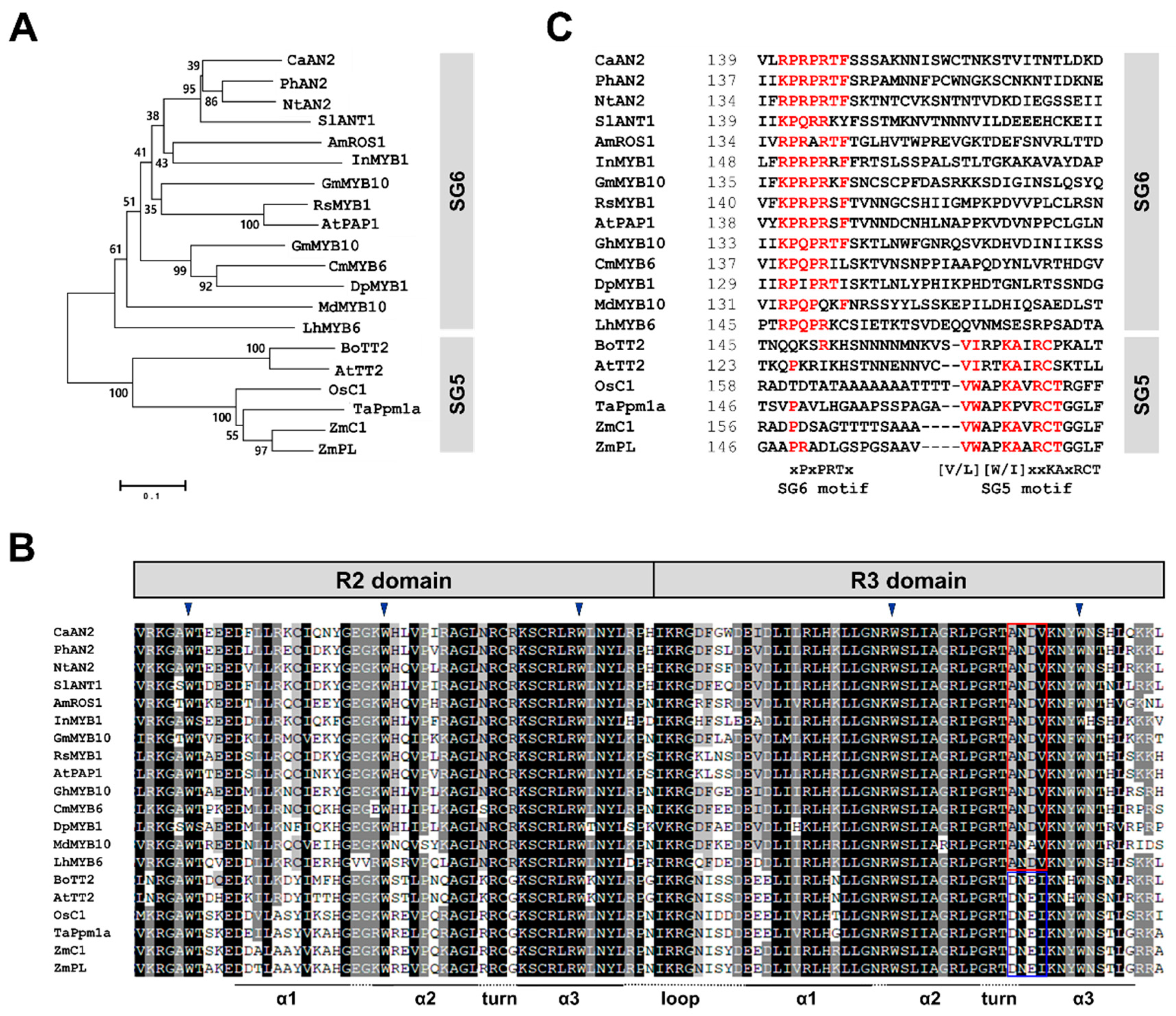
2.3. Isolation of CaAN2 cDNA and Phylogenetic Analysis
2.4. CaAN2 Is an Active Regulator of Anthocyanin Biosynthesis
2.5. CaAN2 Is a Good Candidate Visible Selectable Marker Gene
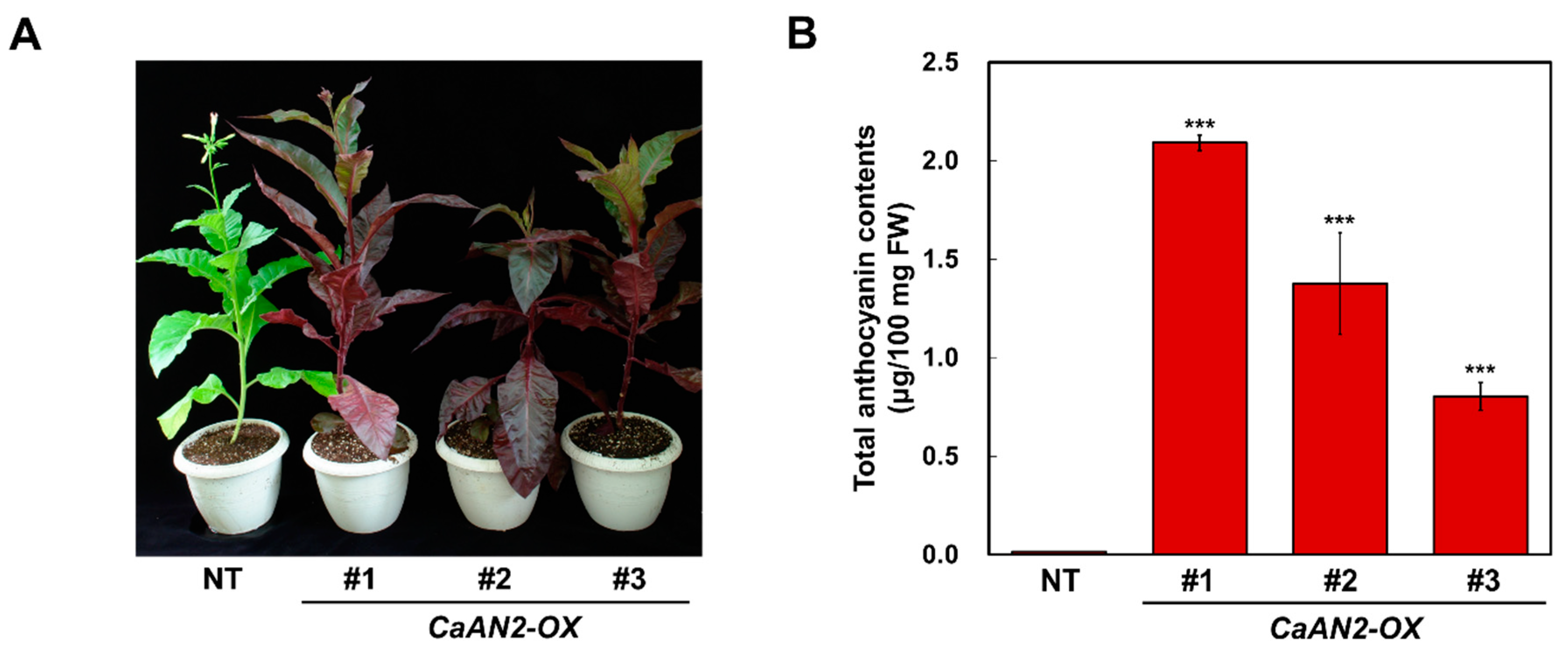
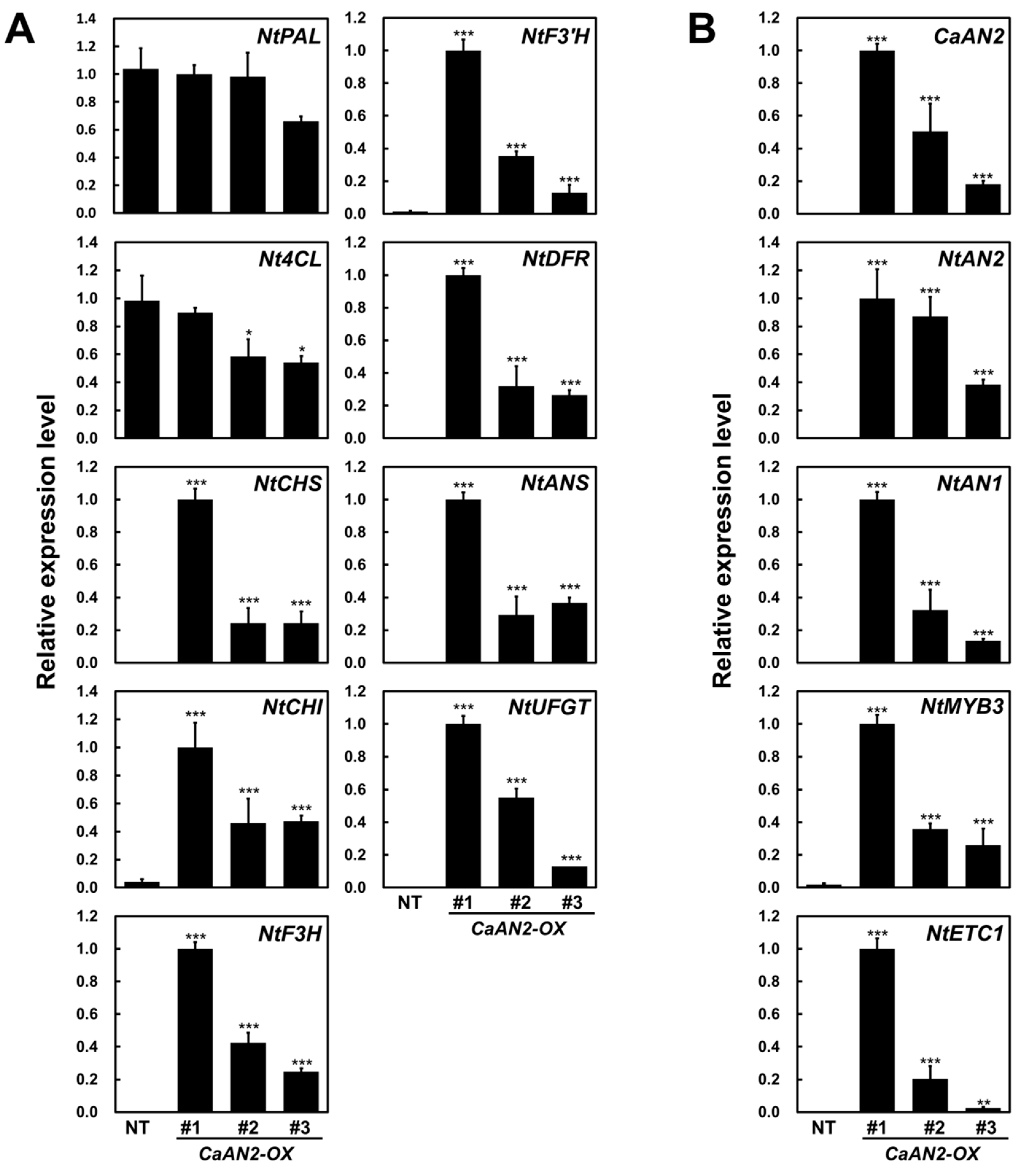
2.6. Ectopic Expression of CaAN2 Strongly Promotes Anthocyanin Biosynthesis
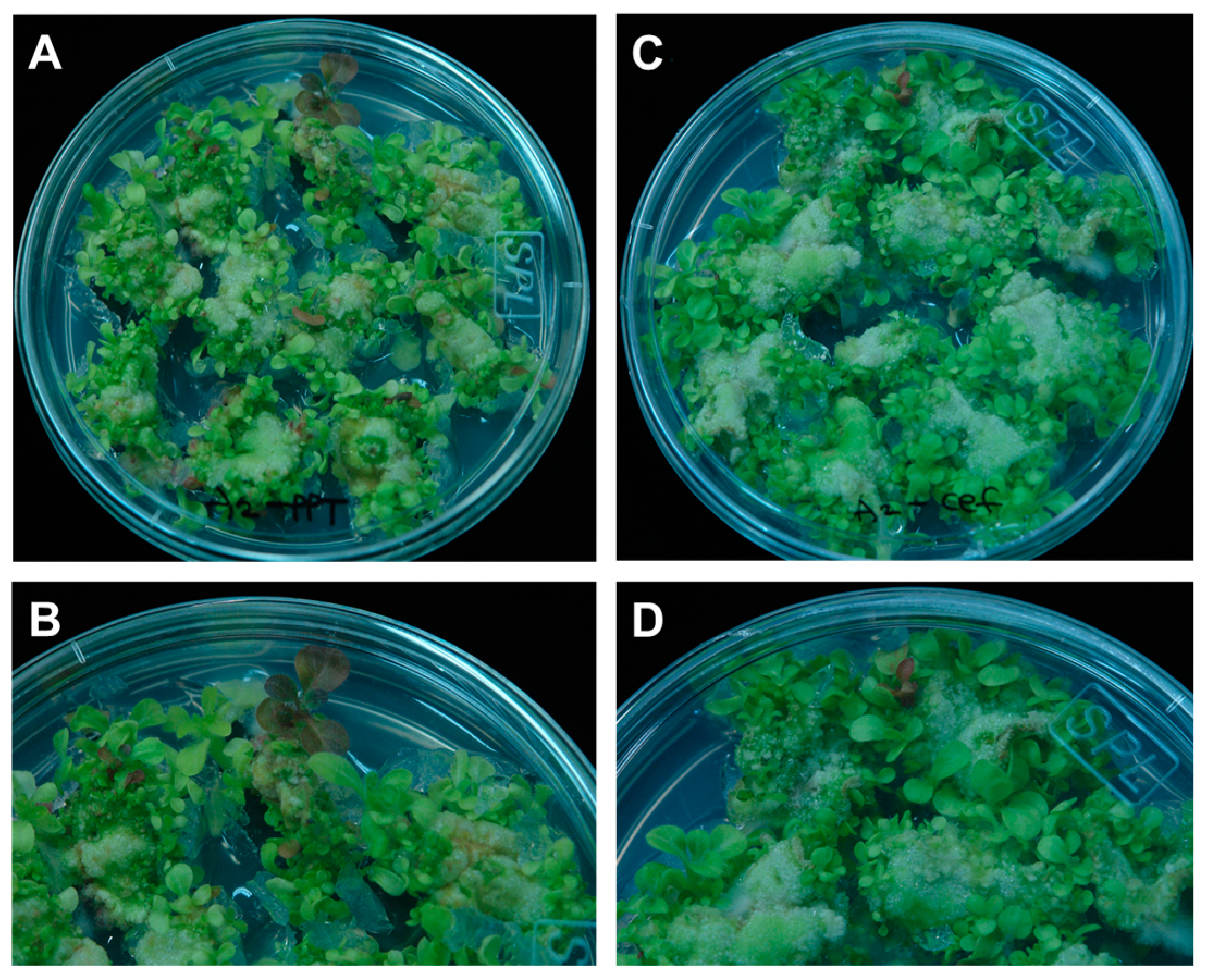
2.7. Plant Coloration Facilitates Detection of and Selection against Chimerism
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. RNA Extraction, cDNA Synthesis and Isolation of Genomic DNA
4.3. Measurement of Total Anthocyanin Contents
4.4. Quantitative Reverse-Transcription PCR (qRT-PCR) Analysis
4.5. Gene Isolation and Sequence Analysis
4.6. In Planta Assays of the Anthocyanin Biosynthesis Activity of CaAN2
4.7. Plant Regeneration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Long, D.; Wu, X.; Yang, Z.; Lenk, I.; Nielsen, K.K.; Gao, C. Comparison of three selectable marker genes for transformation of tall fescue (Festuca arundinacea Schreb.) plants by particle bombardment. In Vitro Cell. Dev. Biol. Plant 2011, 47, 658–666. [Google Scholar] [CrossRef]
- Izawati, A.M.; Masani, M.Y.; Ismanizan, I.; Parveez, G.K. Evaluation on the effectiveness of 2-deoxyglucose-6-phosphate phosphatase (DOGR1) gene as a selectable marker for oil palm (Elaeis guineensis Jacq.) embryogenic calli transformation mediated by Agrobacterium tumefaciens. Front. Plant Sci. 2015, 6, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucht, J.M. Public Acceptance of Plant Biotechnology and GM Crops. Viruses 2015, 7, 4254–4281. [Google Scholar] [CrossRef] [PubMed]
- Lal, M.; Bhardwaj, E.; Chahar, N.; Dangwal, M.; Das, S. (Trans) Gene Flow: Mechanisms, Biosafety Concerns and Mitigation for Containment. In Reproductive Ecology of Flowering Plants: Patterns and Processes; Springer: Berlin/Heidelberg, Germany, 2020; pp. 335–394. [Google Scholar]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.H.; Gresshoff, P.M.; Indrasumunar, A.; Ferguson, B.J. pHairyRed: A Novel Binary Vector Containing the DsRed2 Reporter Gene for Visual Selection of Transgenic Hairy Roots. Mol. Plant 2011, 4, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Saigo, T.; Wang, T.; Watanabe, M.; Tohge, T. Diversity of anthocyanin and proanthocyanin biosynthesis in land plants. Curr. Opin. Plant Biol. 2020, 55, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot. 2011, 62, 2465–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Kortstee, A.J.; Khan, S.A.; Helderman, C.; Trindade, L.M.; Wu, Y.; Visser, R.G.; Brendolise, C.; Allan, A.; Schouten, H.J.; Jacobsen, E. Anthocyanin production as a potential visual selection marker during plant transformation. Transgenic Res. 2011, 20, 1253–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Lee, J.; Rhee, J.; Lee, J.Y.; Lim, S.H. Loss of the R2R3 MYB Transcription Factor RsMYB1 Shapes Anthocyanin Biosynthesis and Accumulation in Raphanus sativus. Int. J. Mol. Sci. 2021, 22, 10927. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Chen, K.; Su, Y.; Jiang, S.; Xu, P.; Murray, J.D. A Root Tip-Specific Expressing Anthocyanin Marker for Direct Identification of Transgenic Tissues by the Naked Eye in Symbiotic Studies. Plants 2021, 10, 605. [Google Scholar] [CrossRef] [PubMed]
- Borovsky, Y.; Oren-Shamir, M.; Ovadia, R.; De Jong, W.; Paran, I. The A locus that controls anthocyanin accumulation in pepper encodes a MYB transcription factor homologous to Anthocyanin2 of Petunia. Theor. Appl. Genet. 2004, 109, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Venkatesh, J.; Kang, M.Y.; Kwon, J.K.; Kang, B.C. A non-LTR retrotransposon activates anthocyanin biosynthesis by regulating a MYB transcription factor in Capsicum annuum. Plant Sci. 2019, 287, 110181. [Google Scholar] [CrossRef] [PubMed]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Kim, D.H.; Yang, J.; Ha, S.H.; Kim, J.K.; Lee, J.Y.; Lim, S.H. An OsKala3, R2R3 MYB TF, Is a Common Key Player for Black Rice Pericarp as Main Partner of an OsKala4, bHLH TF. Front. Plant Sci. 2021, 12, 765049. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Brendolise, C.; Chagneì, D.; Kutty-Amma, S.; Green, S.; Volz, R.; Putterill, J.; Schouten, H.J.; Gardiner, S.E.; Hellens, R.P.; et al. Multiple repeats of a promoter segment causes transcription factor autoregulation in red apples. Plant Cell 2009, 21, 168–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borevitz, J.O.; Xia, Y.; Blount, J.; Dixon, R.A.; Lamb, C. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant Cell 2000, 12, 2383–2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagishi, M.; Shimoyamada, Y.; Nakatsuka, T.; Masuda, K. Two R2R3-MYB genes, homologs of petunia AN2, regulate anthocyanin biosyntheses in flower tepals, tepal spots and leaves of Asiatic hybrid lily. Plant Cell Physiol. 2010, 51, 463–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Bhowmik, S.S.; Cheng, A.Y.; Long, H.; Tan, G.Z.H.; Hoang, T.M.L.; Karbaschi, M.R.; Williams, B.; Higgins, T.J.V.; Mundree, S.G. Robust Genetic Transformation System to Obtain Non-chimeric Transgenic Chickpea. Front. Plant Sci. 2019, 10, 524. [Google Scholar] [CrossRef] [Green Version]
- Yau, Y.Y.; Stewart, C.N., Jr. Less is more: Strategies to remove marker genes from transgenic plants. BMC Biotechnol. 2013, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.H.; Sohn, S.H.; Kim, D.H.; Kim, J.K.; Lee, J.Y.; Kim, Y.M.; Ha, S.H. Use of an anthocyanin production phenotype as a visible selection marker system in transgenic tobacco plant. Plant Biotechnol. Rep. 2012, 6, 203–211. [Google Scholar] [CrossRef]
- Palapol, Y.; Ketsa, S.; Lin-Wang, K.; Ferguson, I.B.; Allan, A.C. A MYB transcription factor regulates anthocyanin biosynthesis in mangosteen (Garcinia mangostana L.) fruit during ripening. Planta 2009, 229, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Schwinn, K.E.; Boase, M.R.; Bradley, J.M.; Lewis, D.H.; Deroles, S.C.; Martin, C.R.; Davies, K.M. MYB and bHLH transcription factor transgenes increase anthocyanin pigmentation in petunia and lisianthus plants, and the petunia phenotypes are strongly enhanced under field conditions. Front. Plant Sci. 2014, 5, 603. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhu, M.; Wu, J.; Ouyang, L.; Wang, R.; Sun, H.; Yan, L.; Wang, L.; Xu, M.; Zhan, H.; et al. Repurposing of Anthocyanin Biosynthesis for Plant Transformation and Genome Editing. Front. Genome Ed. 2020, 2, 607982. [Google Scholar] [CrossRef]
- Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An effective strategy for reliably isolating heritable and Cas9-free arabidopsis mutants generated by CRISPR/Cas9-mediated genome editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.H.; Kim, D.H.; Jung, J.A.; Lee, J.Y. Alternative Splicing of the Basic Helix–Loop–Helix Transcription Factor Gene CmbHLH2 Affects Anthocyanin Biosynthesis in Ray Florets of Chrysanthemum (Chrysanthemum morifolium). Front. Plant Sci. 2021, 12, 669315. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, S.-H.; Kim, D.-H.; Cho, M.-C.; Lee, J.-Y. Chili Pepper AN2 (CaAN2): A Visible Selection Marker for Nondestructive Monitoring of Transgenic Plants. Plants 2022, 11, 820. https://doi.org/10.3390/plants11060820
Lim S-H, Kim D-H, Cho M-C, Lee J-Y. Chili Pepper AN2 (CaAN2): A Visible Selection Marker for Nondestructive Monitoring of Transgenic Plants. Plants. 2022; 11(6):820. https://doi.org/10.3390/plants11060820
Chicago/Turabian StyleLim, Sun-Hyung, Da-Hye Kim, Myeong-Cheoul Cho, and Jong-Yeol Lee. 2022. "Chili Pepper AN2 (CaAN2): A Visible Selection Marker for Nondestructive Monitoring of Transgenic Plants" Plants 11, no. 6: 820. https://doi.org/10.3390/plants11060820