
Utilization of Biomasses from Landscape Conservation Growths Dominated by Common Ragwort (Jacobaea vulgaris Gaertn.) for Biomethanization


Abstract
:1. Introduction
- (I)
- What are the substrate characteristics of a biomass from common ragwort at the recommended stage of cutting?
- (II)
- Can substrates dominated by common ragwort readily be ensiled without the addition of a carbohydrate-rich biomass?
- (III)
- To what extent does ensiling contribute to the reduction of pyrrolizidine alkaloids?
- (IV)
- Which specific methane yield can be attained by the common wet fermentation technique?
- (V)
- Which stage of ragwort development is best for controlling spreading of the plant, yet maintains the best option for efficient biomethanization?
2. Results
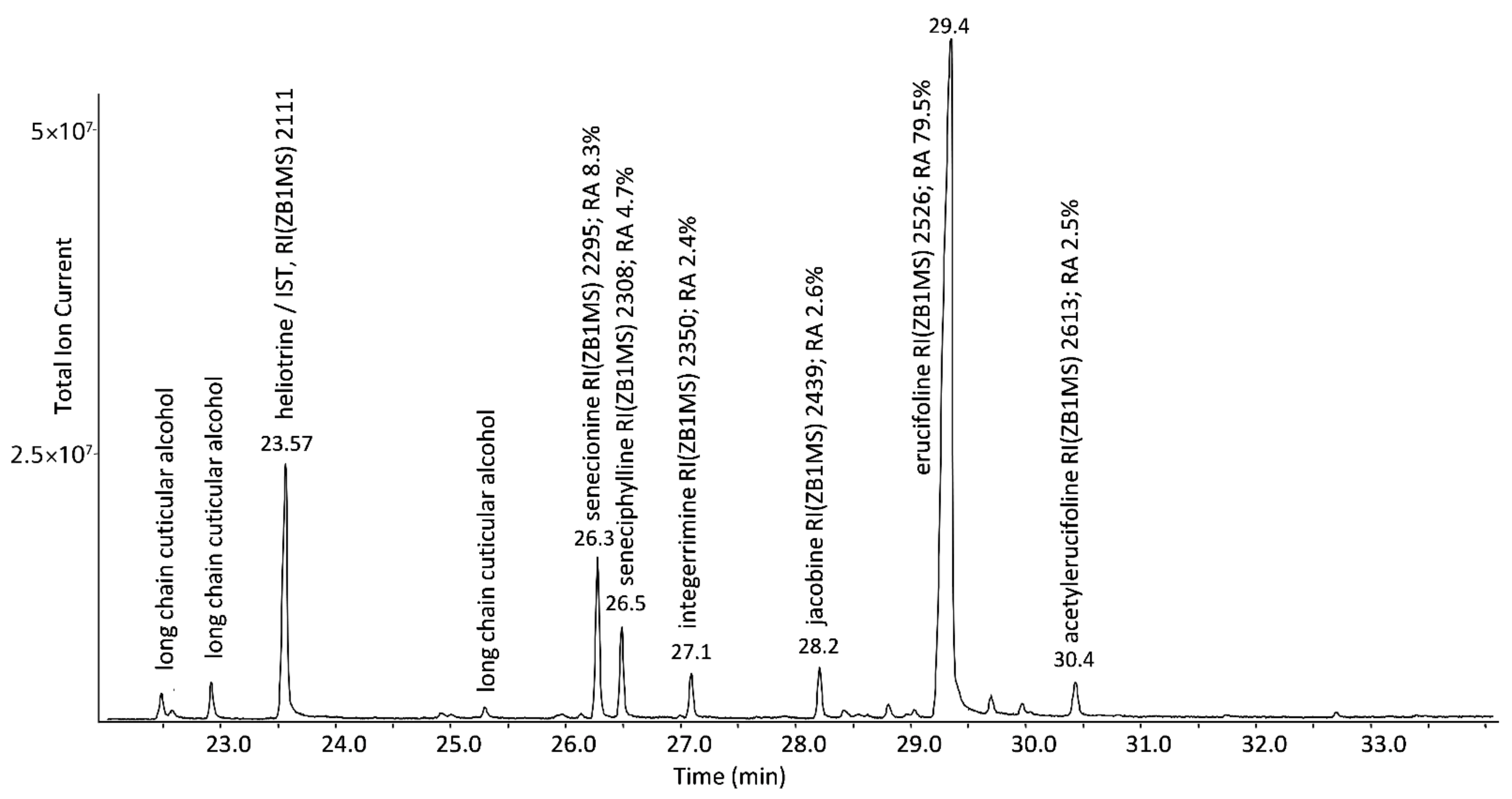
2.1. Characterization of the PA Pattern
2.2. Further Biochemical Properties of the Ragwort’s Biomass
2.2.1. Collection I—Fermentation
2.2.2. Collection II—Growth Stages
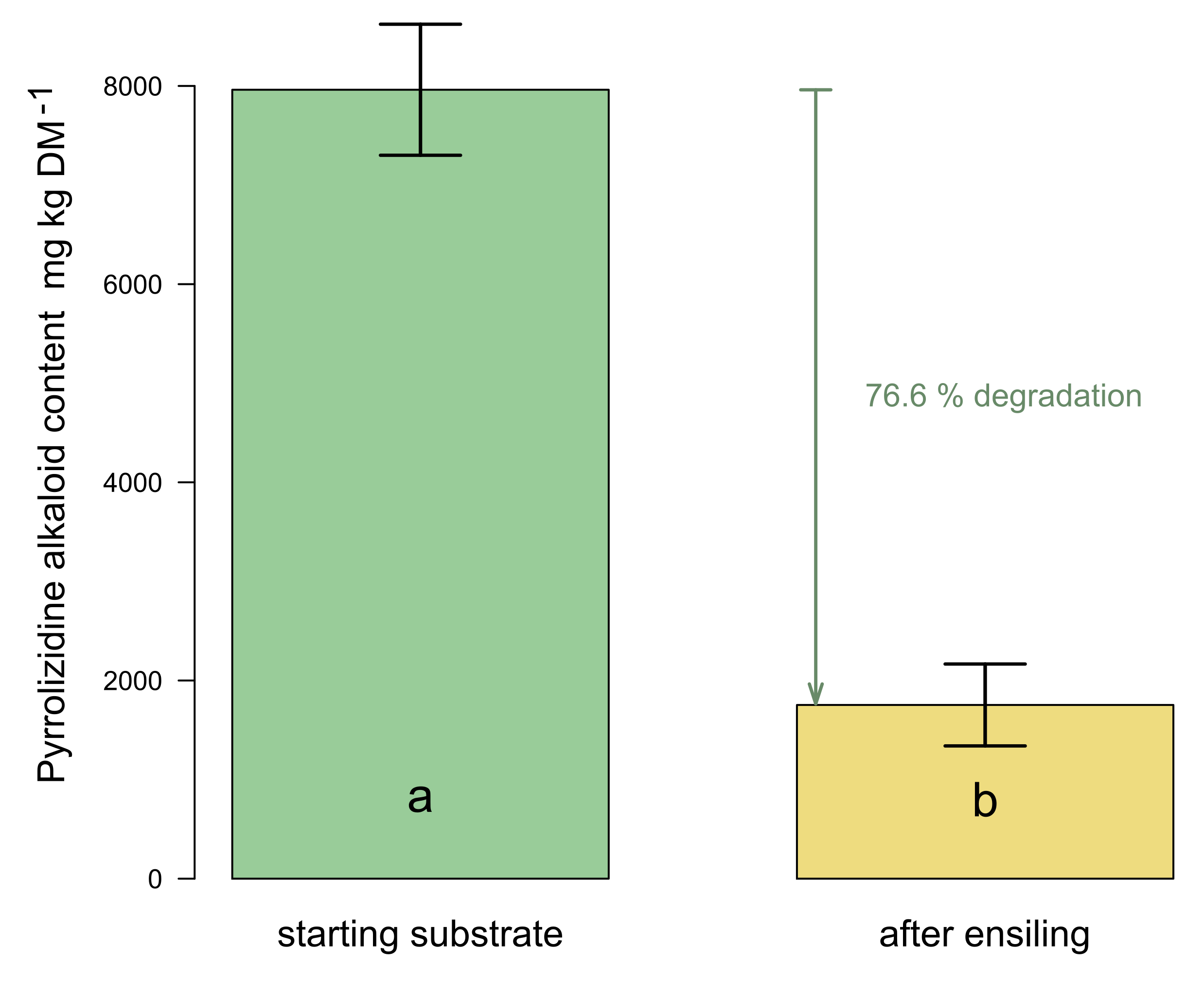
2.3. PA Degradation during Ensiling Process
2.3.1. Results of the Ensiling Experiment
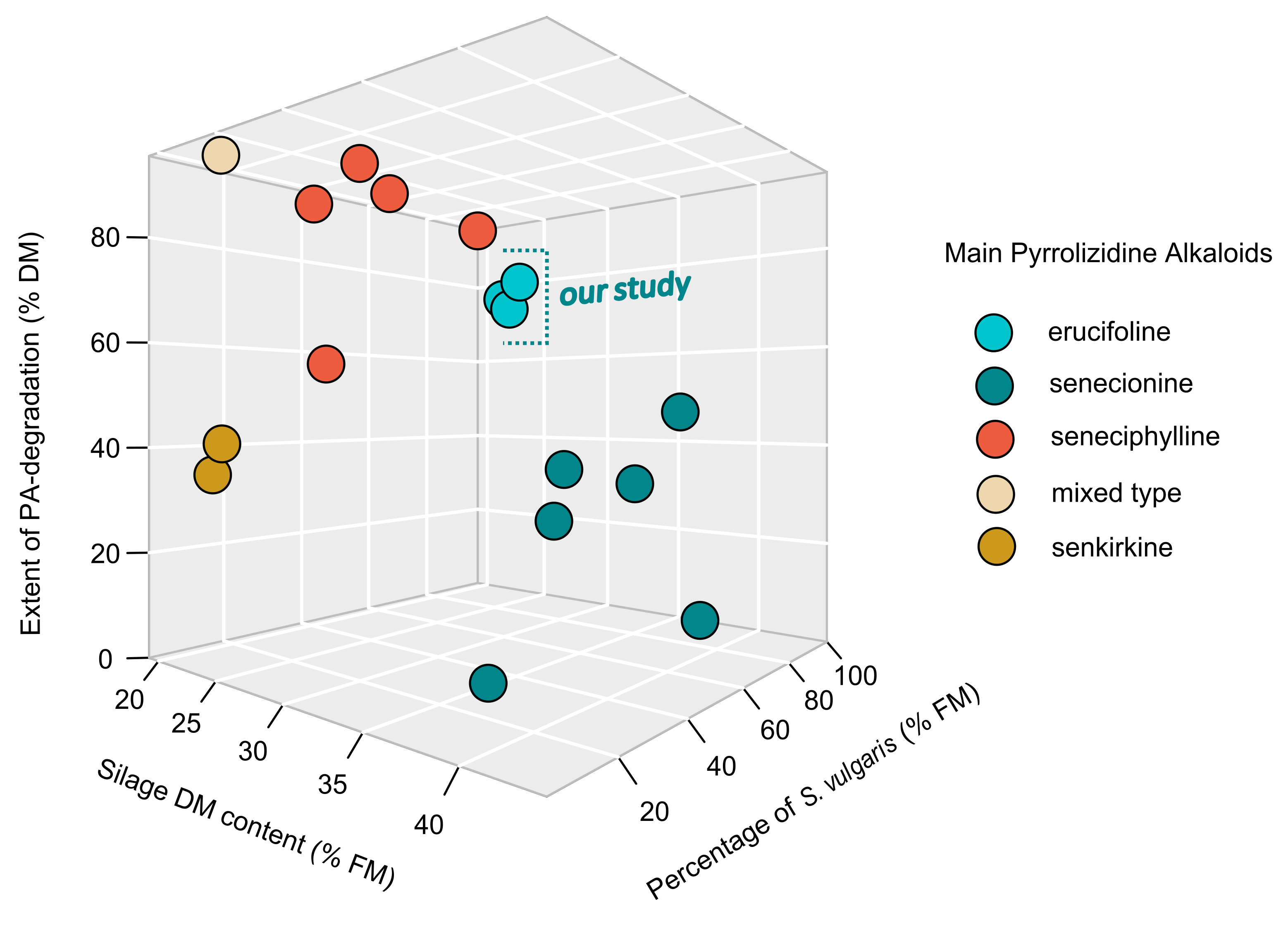
2.3.2. Extent of PA-Degradation in the Context of Similar Studies
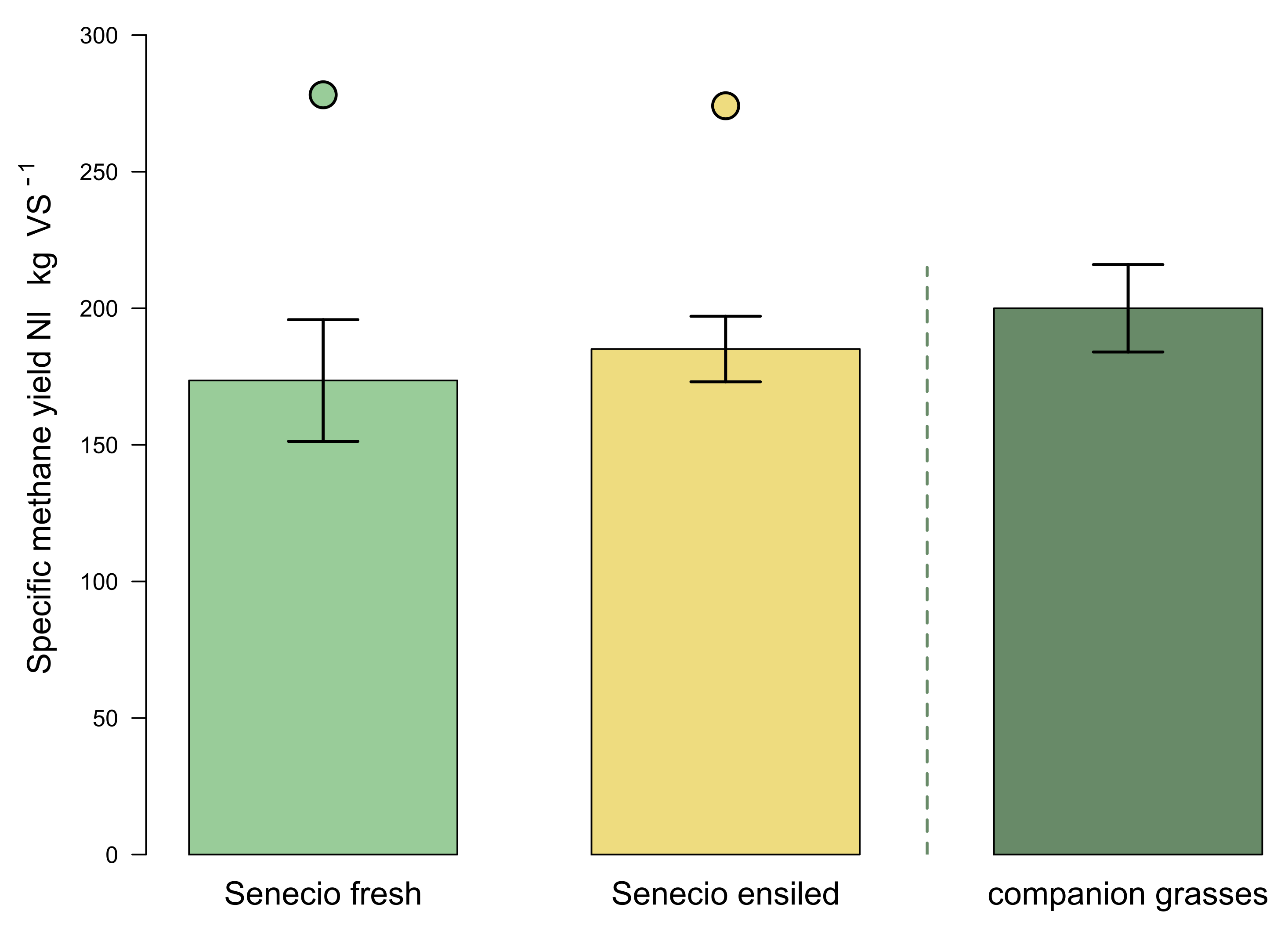
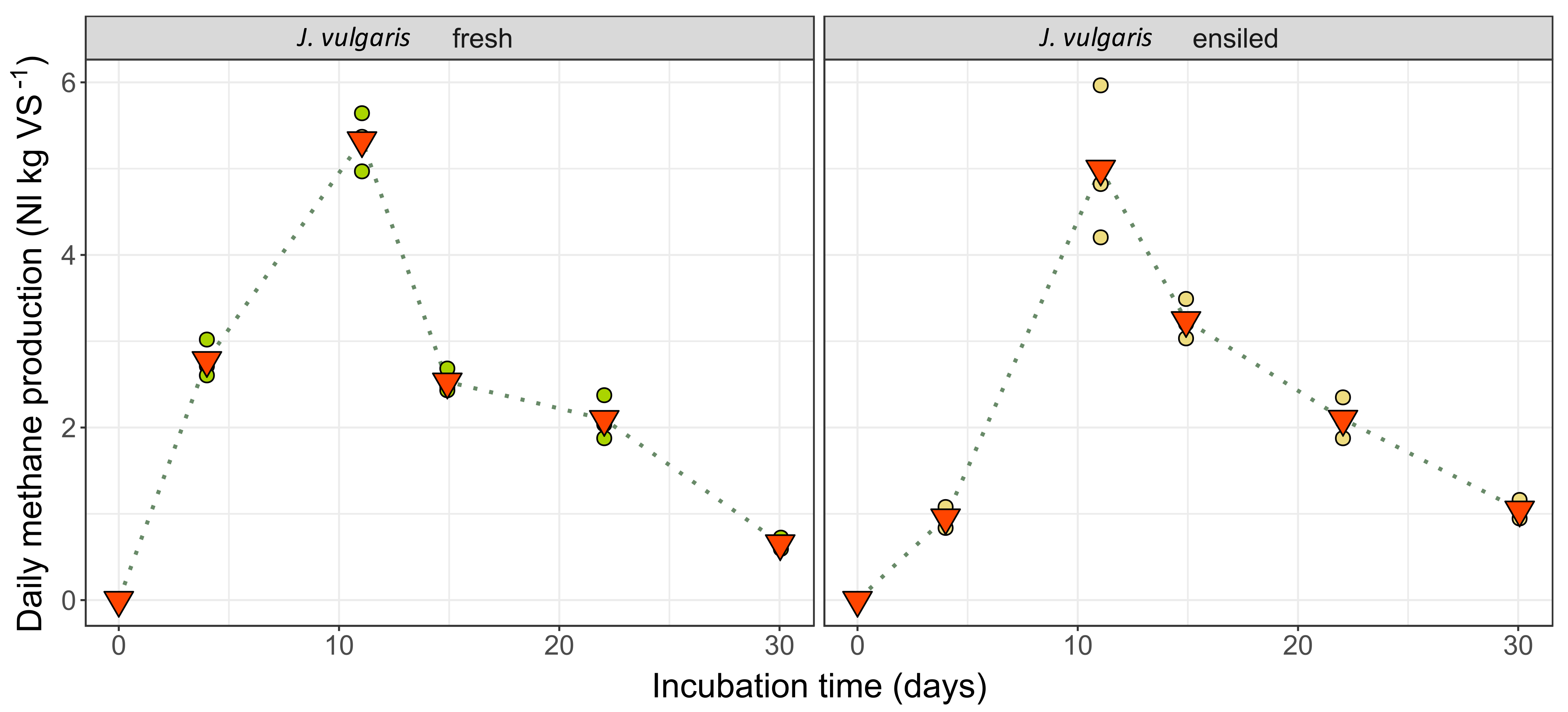
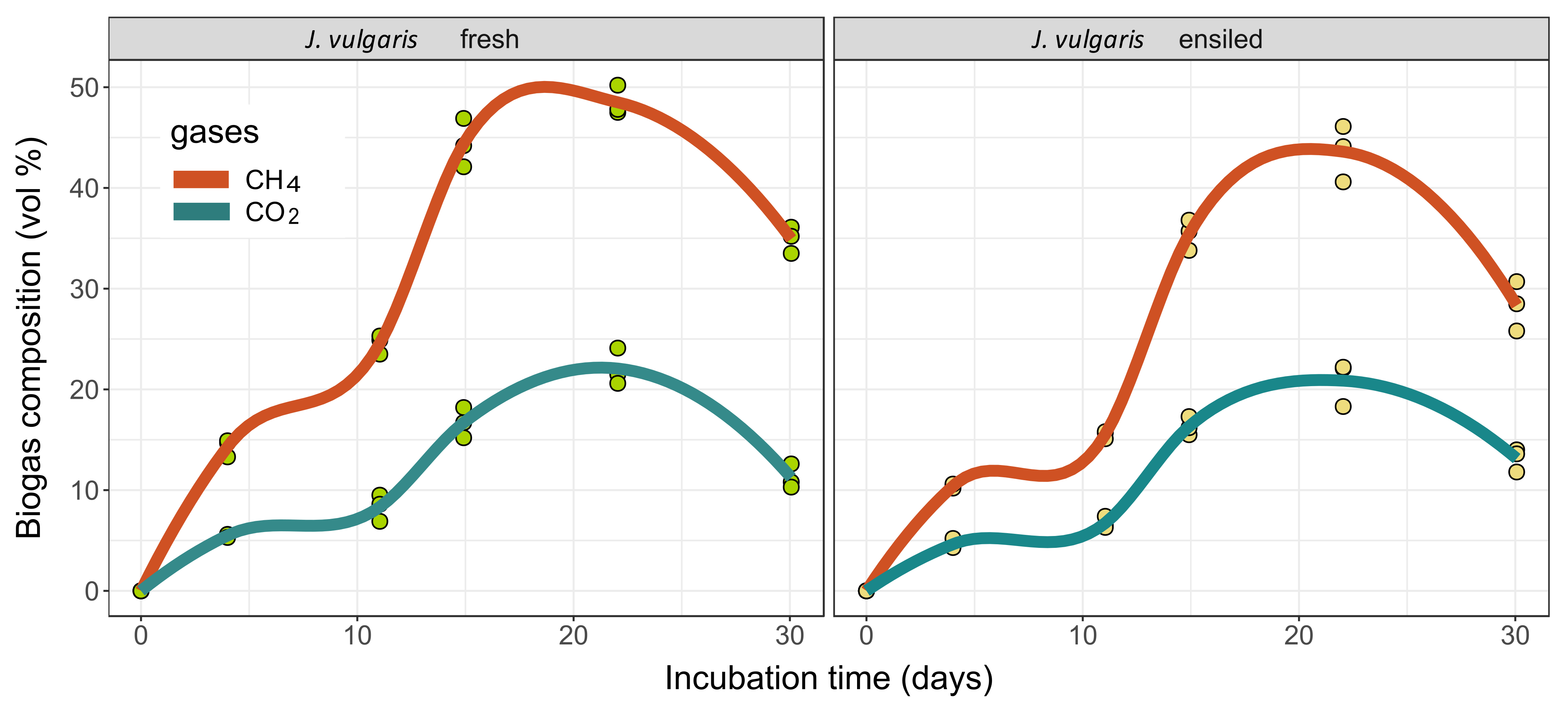
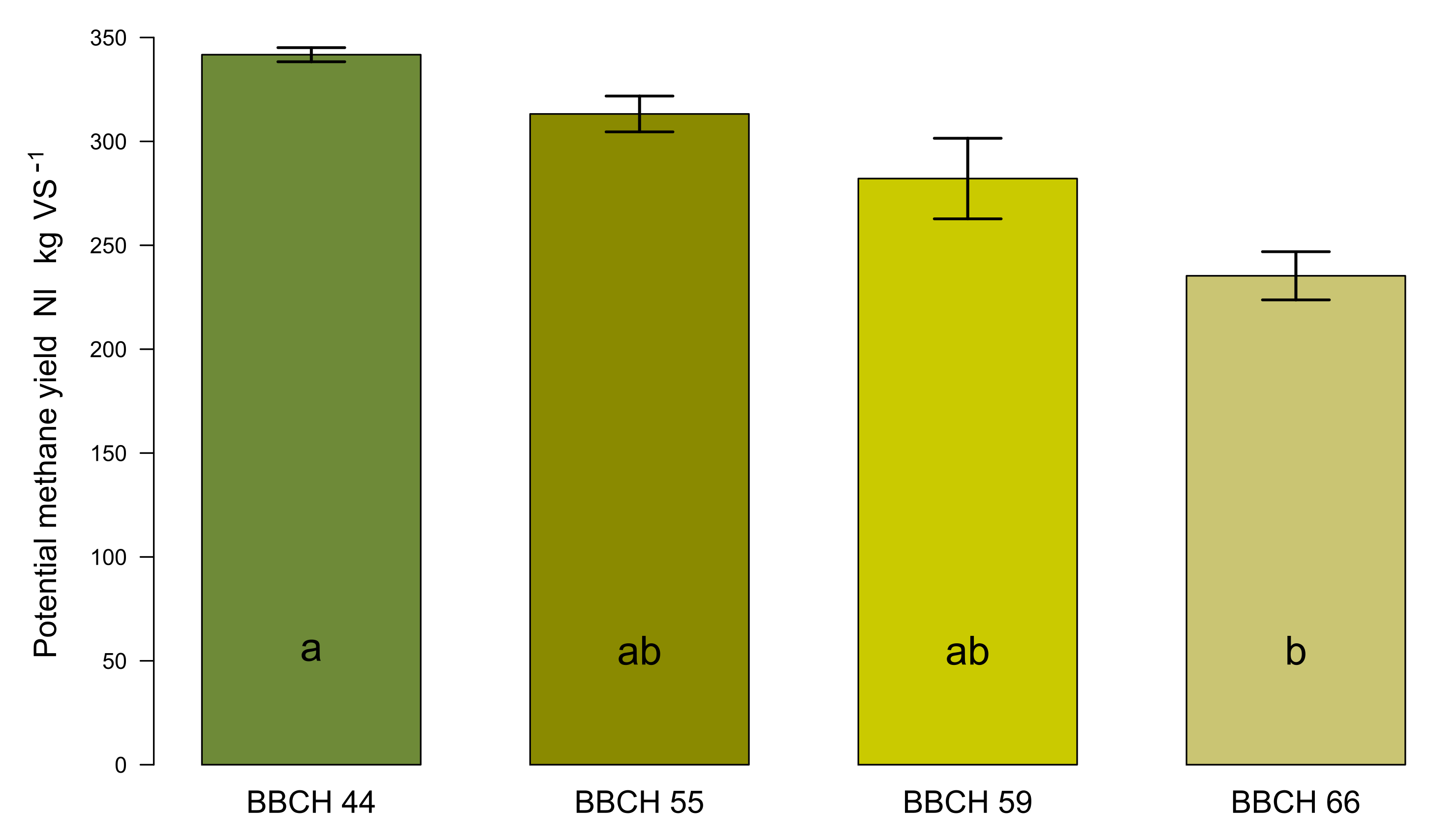
2.4. Methane Yields
2.4.1. Results of the Batch Wet Co-Fermentation Test
2.4.2. Potential Methane Yields at Different Growth Stages
3. Discussion
3.1. Characteristics of Common Ragwort Biomass as Feedstock for Ensiling and Biomethanization
3.2. Degradation of Pyrrolizidine Alkaloids and Their N-Oxides
3.3. Specific Methane Yields of Ragwort Biomasses
3.4. Synthesis
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
4.3. Silage Preparation
4.4. Biochemical Analysis
4.4.1. Substrate Characteristics
4.4.2. Silage Fermentation Characteristics
4.4.3. Quantification of the Total PA/PANO Content
4.4.4. Sample Preparation for GC-MS Analysis
4.4.5. GC-EI-MS PA Pattern Analysis of Raw Plant Materials
4.5. Biogas Yield Determination
4.5.1. Potential Methane Yield Estimation
4.5.2. Batch Wet Co-Fermentation Test
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors/Study | Plant under Study | Main Types of Pyrrolizidine Alkaloids | Percentage of Senecio in the Feedstock | Initial PA-Level before Ensiling | Feedstock Dry Matter Content | Extent of PA-Degradation |
|---|---|---|---|---|---|---|
| (% FM) | (mg/kg DM) | (% FM) | (% DM) | |||
| Müller et al. (2022, this study) | Jacobaea vulgaris Gaertn. | erucifoline | ||||
| lab silo A | 100 | 7714.6 | 22.9 | 80.3 | ||
| lab silo B | 100 | 7741.6 | 22 | 73.5 | ||
| lab silo C | 100 | 8429.1 | 21.4 | 76.2 | ||
| Berendonk & Hünting (2011) | Jacobaea vulgaris Gaertn. | senecionine | ||||
| 100 | 487.5 | 36.4 | 0 | |||
| 75 | 365.7 | 38.6 | 46.8 | |||
| 50 | 243.8 | 39.8 | 34.5 | |||
| 25 | 121.9 | 40.5 | 38.4 | |||
| 10 | 48.8 | 39.9 | 9.4 | |||
| 5 | 24.4 | 43.8 | 33.6 | |||
| Candrian et al. (1984) | Senecio alpinus L. | seneciphylline | ||||
| 100 | 4066.7 | 19.1 | 95.5 | |||
| 41 | 1637.4 | 26.2 | 90.9 | |||
| 23 | 918.6 | 29.1 | 91.4 | |||
| 7 | 280.6 | 30.9 | 80.4 | |||
| 3.5 | 140 | 32.8 | 54.3 | |||
| Gottschalk et al. (2015) | Senecio vulgaris L. | mixed type | 4.5 | 59 | 25 | 91.8 |
| Klevenhusen et al. (2019) | Senecio vulgaris L. | senkirkine | ||||
| 10 | 565 | 22.1 | 35 | |||
| 10 | 635 | 22.9 | 40.9 |
References
- Harper, J.L.; Wood, W.A. Senecio jacobaea L. J. Ecol. 1957, 45, 617. [Google Scholar] [CrossRef]
- Jung, S.; Lauter, J.; Hartung, N.M.; These, A.; Hamscher, G.; Wissemann, V. Genetic and chemical diversity of the toxic herb Jacobaea vulgaris Gaertn. (syn. Senecio jacobaea L.) in Northern Germany. Phytochemistry 2020, 172, 112235. [Google Scholar] [CrossRef] [PubMed]
- Buckland, S.; Thompson, K.; Hodgson, J.; Grime, J. Grassland invasions: Effects of manipulations of climate and management. J. Appl. Ecol. 2001, 38, 301–309. [Google Scholar] [CrossRef]
- Hodkinson, D.J.; Thompson, K. Plant Dispersal: The Role of Man. J. Appl. Ecol. 1997, 34, 1484–1496. [Google Scholar] [CrossRef]
- Wardle, D.A. The Ecology of ragwort (Senecio jacobaea L.)—A review. N. Z. J. Ecol. 1987, 10, 67–76. [Google Scholar]
- Crawley, M.J.; Nachapong, M. The Establishment of Seedlings from Primary and Regrowth Seeds of Ragwort (Senecio jacobaea). J. Ecol. 1985, 73, 255. [Google Scholar] [CrossRef]
- Bosshard, A.; Joshi, J.; Lüscher, A.; Schaffner, U. Tansy ragwort and other ragwort species in Switzerland: An overview: (Jakobs-und andere Kreuzkrautarten: Eine Standortbestimmung). Agrarforschung 2003, 10, 231–235. [Google Scholar]
- Andersson, S. Floral display and pollination success in Senecio jacobaea (Asteraceae): Interactive effects of head and corymb size. Am. J. Bot. 1996, 83, 71–75. [Google Scholar] [CrossRef]
- Kunin, W.E. Patterns of Herbivore Incidence on Experimental Arrays and Field Populations of Ragwort, Senecio jacobaea. Oikos 1999, 84, 515. [Google Scholar] [CrossRef]
- Bradbury, R.B.; Culvenor, C.C. The Alkaloids of Senecio jacobaea L. Aust. J. Chem. 1954, 7, 378. [Google Scholar] [CrossRef]
- Witte, L.; Ernst, L.; Adam, H.; Hartmannt, T. Chemotypes of two pyrrolizidine alkaloid-containing Senecio species. Phytochemistry 1992, 31, 559–565. [Google Scholar] [CrossRef]
- Macel, M.; Vrieling, K.; Klinkhamer, P.G.L. Variation in pyrrolizidine alkaloid patterns of Senecio jacobaea. Phytochemistry 2004, 65, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Goeger, D.E.; Cheeke, P.R.; Schmitz, J.A.; Buhler, D.R. Toxicity of tansy ragwort (Senecio jacobaea) to goats. Am. J. Vet. Res. 1982, 43, 252–254. [Google Scholar] [PubMed]
- Smith, L.W.; Culvenor, C.C. Plant sources of hepatotoxic pyrrolizidine alkaloids. J. Nat. Prod. 1981, 44, 129–152. [Google Scholar] [CrossRef]
- Wiedenfeld, H.; Edgar, J. Toxicity of pyrrolizidine alkaloids to humans and ruminants. Phytochem. Rev. 2011, 10, 137–151. [Google Scholar] [CrossRef]
- White, H.D.; Krumperman, P.H.; Cheeke, P.R.; Deinzer, M.L.; Buhler, D.R. Mutagenic responses of tansy ragwort (Senecio jacobaea) plant, pyrrolizidine alkaloids and metabolites in goat milk with the salmonella/mammalian-microsome mutagenic test. J. Anim. Sci. 1984, 58, 1245–1254. [Google Scholar] [CrossRef]
- Xia, Q.; He, X.; Shi, Q.; Lin, G.; Fu, P.P. Quantitation of DNA reactive pyrrolic metabolites of senecionine—A carcinogenic pyrrolizidine alkaloid by LC/MS/MS analysis. J. Food Drug Anal. 2020, 28, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Edgar, J.A.; Molyneux, R.J.; Colegate, S.M. Pyrrolizidine Alkaloids: Potential Role in the Etiology of Cancers, Pulmonary Hypertension, Congenital Anomalies, and Liver Disease. Chem. Res. Toxicol. 2015, 28, 4–20. [Google Scholar] [CrossRef]
- Cameron, E. A Study of the Natural Control of Ragwort (Senecio jacobaea L.). J. Ecol. 1935, 23, 265. [Google Scholar] [CrossRef]
- Leiss, K.A. Management practices for control of ragwort species. Phytochem. Rev. 2011, 10, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk, C.; Kaltner, F.; Zimmermann, M.; Korten, R.; Morris, O.; Schwaiger, K.; Gareis, M. Spread of Jacobaea vulgaris and Occurrence of Pyrrolizidine Alkaloids in Regionally Produced Honeys from Northern Germany: Inter- and Intra-Site Variations and Risk Assessment for Special Consumer Groups. Toxins 2020, 12, 441. [Google Scholar] [CrossRef] [PubMed]
- Neumann, H.; Huckauf, A. Jakobs-Kreuzkraut (Senecio jacobaea): Eine Ursache für Pyrrolizidin-Alkaloide im Sommerhonig?: Tansy ragwort (Senecio jacobaea): A source of pyrrolizidine alkaloids in summer honey? J. Verbr. Lebensm. 2016, 11, 105–115. [Google Scholar] [CrossRef]
- Schrenk, D.; Gao, L.; Lin, G.; Mahony, C.; Mulder, P.P.J.; Peijnenburg, A.; Pfuhler, S.; Rietjens, I.M.C.M.; Rutz, L.; Steinhoff, B.; et al. Pyrrolizidine alkaloids in food and phytomedicine: Occurrence, exposure, toxicity, mechanisms, and risk assessment—A review. Food Chem. Toxicol. 2020, 136, 111107. [Google Scholar] [CrossRef] [PubMed]
- Kempf, M.; Wittig, M.; Reinhard, A.; von der Ohe, K.; Blacquière, T.; Raezke, K.-P.; Michel, R.; Schreier, P.; Beuerle, T. Pyrrolizidine alkaloids in honey: Comparison of analytical methods. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2011, 28, 332–347. [Google Scholar] [CrossRef] [Green Version]
- Suter, M.; Siegrist-Maag, S.; Connolly, J.; Lüscher, A. Can the occurrence of Senecio jacobaea be influenced by management practice? Weed Res. 2007, 47, 262–269. [Google Scholar] [CrossRef]
- Möhler, H.; Diekötter, T.; Herrmann, J.D.; Donath, T.W. Allelopathic vs. autotoxic potential of a grassland weed—evidence from a seed germination experiment. Plant Ecol. Divers. 2018, 11, 539–549. [Google Scholar] [CrossRef]
- UK Department for Environment Food and Rural Affairs. The Weeds Act, Chapter 54. Available online: https://www.legislation.gov.uk/ukpga/2003/40/introduction/enacted (accessed on 6 January 2022).
- Roberts, P.D.; Pullin, A.S. The effectiveness of management interventions used to control ragwort species. Environ. Manag. 2007, 39, 691–706. [Google Scholar] [CrossRef]
- Augustin, B.; Oehme, S.; Mattern, G.; Roth, W. Influence of different measures against Senecio jacobaea concerning efficacy and biological diversity. Julius-Kühn-Archiv 2018, 458, 47–53. (In German) [Google Scholar] [CrossRef]
- Möhler, H. Basic and Applied Studies to Evaluate Management Options for the Regulation of Tansy Ragwort (Jacobaea vulgaris). Inaugural Dissertation, Christian-Albrechts-University, Kiel, Germany, 2020. [Google Scholar]
- Siegrist-Maag, S.; Luescher, A.; Suter, M. Reaktion von Jakobs-Kreuzkraut auf Schnitt: Sensitive reaction of ragwort (Senecio jacobaea) to cutting dates. Agrarforschung 2008, 15, 338–343. [Google Scholar]
- Gottschalk, C.; Ronczka, S.; Preiß-Weigert, A.; Ostertag, J.; Klaffke, H.; Schafft, H.; Lahrssen-Wiederholt, M. Pyrrolizidine alkaloids in natural and experimental grass silages and implications for feed safety. Anim. Feed Sci. Technol. 2015, 207, 253–261. [Google Scholar] [CrossRef]
- Driehuis, F.; Wilkinson, J.M.; Jiang, Y.; Ogunade, I.; Adesogan, A.T. Silage review: Animal and human health risks from silage. J. Dairy Sci. 2018, 101, 4093–4110. [Google Scholar] [CrossRef] [PubMed]
- Kalač, P.; Kaltner, F. Pyrrolizidine alkaloids of European Senecio/Jacobaea species in forage and their carry-over to milk: A review. Anim. Feed Sci. Technol. 2021, 280, 115062. [Google Scholar] [CrossRef]
- Hama, J.R.; Strobel, B.W. Occurrence of pyrrolizidine alkaloids in ragwort plants, soils and surface waters at the field scale in grassland. Sci. Total Environ. 2021, 755, 142822. [Google Scholar] [CrossRef] [PubMed]
- Crews, C.; Driffield, M.; Berthiller, F.; Krska, R. Loss of pyrrolizidine alkaloids on decomposition of ragwort (Senecio jacobaea) as measured by LC-TOF-MS. J. Agric. Food Chem. 2009, 57, 3669–3673. [Google Scholar] [CrossRef]
- Chmit, M.S.; Müller, J.; Wiedow, D.; Horn, G.; Beuerle, T. Biodegradation and utilization of crop residues contaminated with poisonous pyrrolizidine alkaloids. J. Environ. Manag. 2021, 290, 112629. [Google Scholar] [CrossRef]
- Zhao, S.; Schmidt, S.; Gao, H.; Li, T.; Chen, X.; Hou, Y.; Chadwick, D.; Tian, J.; Dou, Z.; Zhang, W.; et al. A Prominent Role for Precision Composting in Sustainable Agriculture; Research Square Platform LLC: Durham, NC, USA, 2021. [Google Scholar]
- Capodaglio, A.; Callegari, A.; Lopez, M. European Framework for the Diffusion of Biogas Uses: Emerging Technologies, Acceptance, Incentive Strategies, and Institutional-Regulatory Support. Sustainability 2016, 8, 298. [Google Scholar] [CrossRef] [Green Version]
- Van Meerbeek, K.; Appels, L.; Dewil, R.; van Beek, J.; Bellings, L.; Liebert, K.; Muys, B.; Hermy, M. Energy potential for combustion and anaerobic digestion of biomass from low-input high-diversity systems in conservation areas. GCB Bioenergy 2015, 7, 888–898. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Hahn, J. Ensilability of Biomass From Effloresced Flower Strips as Co-substrate in Bioenergy Production. Front. Bioeng. Biotechnol. 2020, 8, 14. [Google Scholar] [CrossRef]
- Berendonk, C.; Cerff, D.; Hünting, K.; Wiedenfeld, H.; Becerra, J.; Kuschak, M. Pyrrolizidine alkaloid level in Senecio jacobaea and Senecio erraticus-the effect of plant organ and forage conservation. Grassl. Sci. Eur. 2010, 15, 669–671. [Google Scholar]
- Becerra-Jimenez, J.; Kuschak, M.; Röder, E.T.; Wiedenfeld, H. Toxic pyrrolizidinalkaloids as undesired contaminants in food and feed: Degradation of the PAs from Senecio jacobaea in silage. Pharmazie 2013, 68, 636–639. [Google Scholar] [CrossRef]
- Ferrero, F.; Dinuccio, E.; Rollé, L.; Tabacco, E.; Borreani, G. Suitability of cardoon (Cynara cardunculus L.) harvested at two stages of maturity to ensiling and methane production. Biomass Bioenergy 2020, 142, 105776. [Google Scholar] [CrossRef]
- Berendonk, C.; Hünting, K. Einfluss der Silierung auf den Gehalt an Pyrrolizidinalkaloiden von Senecio jacobaea in Abhängigkeit vom Senecio-Gehalt im Siliergut. Mitteilungen AGGF 2011, 12, 159–162. [Google Scholar]
- Candrian, U.; Luethy, J.; Schmid, P.; Schlatter, C.; Gallasz, E. Stability of pyrrolizidine alkaloids in hay and silage. J. Agric. Food Chem. 1984, 32, 935–937. [Google Scholar] [CrossRef]
- Klevenhusen, F.; Pieper, R.; Winter, J.; Ronczka, S.; Speer, K. Stability of pyrrolizidine alkaloids from Senecio vernalis in grass silage under different ensilage conditions. J. Sci. Food Agric. 2019, 99, 6649–6654. [Google Scholar] [CrossRef] [Green Version]
- Meserszmit, M.; Chrabąszcz, M.; Chylińska, M.; Szymańska-Chargot, M.; Trojanowska-Olichwer, A.; Kącki, Z. The effect of harvest date and the chemical characteristics of biomass from Molinia meadows on methane yield. Biomass Bioenergy 2019, 130, 105391. [Google Scholar] [CrossRef]
- Scherzinger, M.; Kulbeik, T.; Kaltschmitt, M. Autoclave pre-treatment of green wastes–Effects of temperature, residence time and rotation speed on fuel properties. Fuel 2020, 273, 117796. [Google Scholar] [CrossRef]
- Herrmann, C.; Idler, C.; Heiermann, M. Biogas crops grown in energy crop rotations: Linking chemical composition and methane production characteristics. Bioresour. Technol. 2016, 206, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivamani, S.; Saikat, B.; Naveen Prasad, B.S.; Baalawy, A.A.S.; Al-Mashali, S.M.A. A Comprehensive Review on Microbial Technology for Biogas Production. In Bioenergy Research: Revisiting Latest Development, 1st ed.; Srivastava, M., Srivastava, N., Singh, R., Eds.; Springer: Singapore, 2021; pp. 53–78. ISBN 978-981-33-4614-7. [Google Scholar]
- Triolo, J.M.; Pedersen, L.; Qu, H.; Sommer, S.G. Biochemical methane potential and anaerobic biodegradability of non-herbaceous and herbaceous phytomass in biogas production. Bioresour. Technol. 2012, 125, 226–232. [Google Scholar] [CrossRef]
- Dandikas, V.; Heuwinkel, H.; Lichti, F.; Drewes, J.E.; Koch, K. Correlation between biogas yield and chemical composition of energy crops. Bioresour. Technol. 2014, 174, 316–320. [Google Scholar] [CrossRef]
- Stachowiak–Wencek, A.; Bocianowski, J.; Waliszewska, H.; Borysiak, S.; Waliszewska, B.; Zborowska, M. Statistical prediction of biogas and methane yields during anaerobic digestion based on the composition of lignocellulosic biomass. BioRes 2021, 16, 7086–7100. [Google Scholar] [CrossRef]
- Raffrenato, E.; Fievisohn, R.; Cotanch, K.W.; Grant, R.J.; Chase, L.E.; van Amburgh, M.E. Effect of lignin linkages with other plant cell wall components on in vitro and in vivo neutral detergent fiber digestibility and rate of digestion of grass forages. J. Dairy Sci. 2017, 100, 8119–8131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weißbach, F. On Assessing the Gas Production Potential of Renewable Primary Products (Zur Bewertung des Gasbildungspotenzials von nachwachsenden Rohstoffen). Landtechnik 2008, 63, 356–358. [Google Scholar] [CrossRef]
- Meak, P. Biochemische Charakterisierung von Pflanzen Unterschiedlicher Nutzungsintensität zur Ableitung von Parametern für die Ermittlung des Energetischen Futterwertes. Inaugural Dissertation, Humboldt University, Berlin, Germany, 2002. [Google Scholar]
- Frost, R.A.; Wilson, L.M.; Launchbaugh, K.L.; Hovde, E.M. Seasonal Change in Forage Value of Rangeland Weeds in Northern Idaho. Invasive Plant Sci. Manag. 2008, 1, 343–351. [Google Scholar] [CrossRef]
- Purcell, P.J.; Boland, T.M.; O’Brien, M.; O’Kiely, P. In vitro rumen methane output of forb species sampled in spring and summer. AFSci 2012, 21, 83–90. [Google Scholar] [CrossRef]
- Millard, P. The accumulation and storage of nitrogen by herbaceous plants. Plant Cell Environ. 1988, 11, 1–8. [Google Scholar] [CrossRef]
- Debessai, W.T. Evaluation of Endophyte-Infected Tall Fescue Products, Their Interaction with Senecio jacobaea in Ruminants, and Detoxification of Alkaloids by Ammoniation or Ensiling after Grinding. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1998. [Google Scholar]
- McDonald, P.; Henderson, A.R.; Heron, S. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Elferink, S.J.W.H.O.; Spoelstra, S.F. Microbiology of Ensiling. In Silage Science and Technology; Buxton, D.R., Muck, R.E., Harrison, J., Eds.; Soil Science Society of America: Madison, WI, USA, 2003; pp. 31–93. ISBN 9780891182344. [Google Scholar]
- Cramer, L.; Schiebel, H.-M.; Ernst, L.; Beuerle, T. Pyrrolizidine alkaloids in the food chain: Development, validation, and application of a new HPLC-ESI-MS/MS sum parameter method. J. Agric. Food Chem. 2013, 61, 11382–11391. [Google Scholar] [CrossRef]
- Murphy, J.D.; Thamsiriroj, T. Fundamental science and engineering of the anaerobic digestion process for biogas production. In The Biogas Handbook: Science, Production and Applications; Wellinger, A., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 104–130. ISBN 9780857094988. [Google Scholar]
- Teixeira Franco, R.; Buffière, P.; Bayard, R. Ensiling for biogas production: Critical parameters. A review. Biomass Bioenergy 2016, 94, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, C.; Heiermann, M.; Idler, C. Effects of ensiling, silage additives and storage period on methane formation of biogas crops. Bioresour. Technol. 2011, 102, 5153–5161. [Google Scholar] [CrossRef]
- Mangold, A.; Lewandowski, I.; Hartung, J.; Kiesel, A. Miscanthus for biogas production: Influence of harvest date and ensiling on digestibility and methane hectare yield. GCB Bioenergy 2019, 11, 50–62. [Google Scholar] [CrossRef]
- Wannasek, L.; Ortner, M.; Amon, B.; Amon, T. Sorghum, a sustainable feedstock for biogas production? Impact of climate, variety and harvesting time on maturity and biomass yield. Biomass Bioenergy 2017, 106, 137–145. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono-and Dicotyledonous Plants: BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Berlin/Braunschweig, Germany, 2001. [Google Scholar]
- De Boever, J.L.; Cottyn, B.G.; Buysse, F.X.; Wainman, F.W.; Vanacker, J.M. The use of an enzymatic technique to predict digestibility, metabolizable and net energy of compound feedstuffs for ruminants. Anim. Feed Sci. Technol. 1986, 14, 203–214. [Google Scholar] [CrossRef]
- Naumann, C.; Bassler, R. Die Chemische Untersuchung von Futtermitteln, 3. Aufl.; VDLUFA-Verl.: Darmstadt, Germany, 2012; ISBN 9783941273146. [Google Scholar]
- Letsyo, E.; Jerz, G.; Winterhalter, P.; Lindigkeit, R.; Beuerle, T. Incidence of Pyrrolizidine Alkaloids in Herbal Medicines from German Retail Markets: Risk Assessments and Implications to Consumers. Phytother. Res. 2017, 31, 1903–1909. [Google Scholar] [CrossRef] [PubMed]
- Frölich, C.; Ober, D.; Hartmann, T. Tissue distribution, core biosynthesis and diversification of pyrrolizidine alkaloids of the lycopsamine type in three Boraginaceae species. Phytochemistry 2007, 68, 1026–1037. [Google Scholar] [CrossRef] [PubMed]
- Weißbach, F.; Strubelt, C. Correcting the dry matter content of grass silages as a substrate for biogas production. Landtechnik 2008, 63, 210–211. [Google Scholar] [CrossRef]
- Verein Deutscher Ingenieure. Fermentation of Organic Materials-Characterisation of the Substrate, Sampling, Collection of Material Data, Fermentation Tests: Vergärung Organischer Stoffe-Substratcharakterisierung, Probenahme, Stoffdatenerhebung, Gärversuche; Beuth-Verlag: Berlin, Germany, 2016; ICS 13.030.30, 27.190 (VDI 4630); Available online: https://www.vdi.de/fileadmin/pages/vdi_de/redakteure/richtlinien/inhaltsverzeichnisse/2385990.pdf (accessed on 24 January 2022).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.eea.europa.eu/data-and-maps/indicators/oxygen-consuming-substances-in-rivers/r-development-core-team-2006 (accessed on 24 January 2022).







| Status of Ragwort Biomass | Feedstock Characteristics | Means (sd) |
|---|---|---|
| Chopped starting material before ensiling | Parameters of Ensilability | n = 3 |
| Dry matter content at start (DMC, g kg−1) | 224.3 (3.79) | |
| Crude protein (CP, g kg−1 DM) | 108.7 (0.41) | |
| Water-soluble carbohydrates (WSCH, g kg−1 DM) | 102.3 (11.72) | |
| WSCH:CP-ratio | 0.94 (0.12) | |
| Substrate after 92 days of lab-scale ensiling | Fermentation patterns | |
| Dry matter content after ensiling (DMC, g kg−1) | 222.5 (0.12) | |
| pH-value | 4.50 (0.00) | |
| Lactic acid (g kg−1 DM) | 34.40 (4.48) | |
| Acetic acid (g kg−1 DM) | 8.65 (0.83) | |
| Ethanol (g kg−1 DM) | 1.60 (0.01) | |
| Butyric acid (g kg−1 DM) | <0.50 | |
| Ammonia (g kg−1 FM) | 0.34 (0.01) |
| Stage of Development (BBCH) 1 | Vegetative Parts Reached ca. 40% of Final Size (44) | First Flowers Visible, Largely Still Closed (55) | Inflorescence Fully Emerged (59) | Flowering Nearly Finished (66) | Effect of Stage 3 |
|---|---|---|---|---|---|
| Sampling Date | 1 June 2021 | 8 June 2021 | 16 June 2021 | 5 July 2021 | |
| Corresponding grassland use | cut for silage | cut for hay | late cut for hay | conservation cut | |
| Averaged plant height (cm) 2 | 33.70 (3.91) | 56.80 (0.43) | 84.33 (4.41) | 99.33 (3.26) | p < 0.001 *** |
| Dry matter content (g kg−1) 2 | 116.79 (3.70) | 156.98 (0.88) | 192.57 (7.34) | 267.26 (6.88) | p < 0.001 *** |
| Stage of Development (BBCH) 1 | BBCH 44 | BBCH 55 | BBCH 59 | BBCH 66 | Effect of Stage 2 |
|---|---|---|---|---|---|
| Crude ash (CA, g kg−1 DM) | 100.73 (2.37) | 76.13 (0.90) | 55.60 (3.22) | 45.30 (1.34) | p < 0.001 *** |
| Crude protein (CP, g kg−1 DM) | 155.92 (4.32) | 118.92 (6.20) | 93.04 (3.17) | 79.95 (4.48) | p < 0.001 *** |
| Crude fiber (CF, g kg−1 DM) | 161.28 (7.02) | 220.28 (10.11) | 293.31 (14.93) | 340.30 (7.78) | p < 0.001 *** |
| Crude lipid (CL, g kg−1 DM) | 23.93 (1.35) | 22.58 (2.15) | 18.35 (1.02) | 18.02 (0.53) | p = 0.002 ** |
| Neutral detergent fiber (aNDFOM, g kg−1 DM) | 284.39 (8.79) | 388.12 (16.04) | 479.97 (22.89) | 537.45 (12.12) | p < 0.001 *** |
| Acid detergent fiber (ADFOM, g kg−1 DM) | 224.26 (6.11) | 299.65 (12.96) | 378.60 (14.20) | 428.86 (7.08) | p < 0.001 *** |
| Hemicellulose (HC, g kg−1 DM) | 60.13 (6.26) | 88.46 (4.15) | 101.37 (8.69) | 108.59 (5.06) | p < 0.001 *** |
| Water-soluble carbohydrates (WSCH, g kg−1 DM) | 106.15 (3.34) | 93.00 (8.15) | 77.58 (1.71) | 42.15 (4.71) | p < 0.001 *** |
| Enzyme-insoluble organic matter (EISOM, g kg−1 DM) | 131.94 (8.63) | 262.70 (14.22) | 386.38 (32.71) | 534.36 (14.54) | p < 0.001 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, J.; Wiedow, D.; Chmit, M.S.; Beuerle, T. Utilization of Biomasses from Landscape Conservation Growths Dominated by Common Ragwort (Jacobaea vulgaris Gaertn.) for Biomethanization. Plants 2022, 11, 813. https://doi.org/10.3390/plants11060813
Müller J, Wiedow D, Chmit MS, Beuerle T. Utilization of Biomasses from Landscape Conservation Growths Dominated by Common Ragwort (Jacobaea vulgaris Gaertn.) for Biomethanization. Plants. 2022; 11(6):813. https://doi.org/10.3390/plants11060813
Chicago/Turabian StyleMüller, Jürgen, Denny Wiedow, Mohammad Said Chmit, and Till Beuerle. 2022. "Utilization of Biomasses from Landscape Conservation Growths Dominated by Common Ragwort (Jacobaea vulgaris Gaertn.) for Biomethanization" Plants 11, no. 6: 813. https://doi.org/10.3390/plants11060813