
A New Formulation for Strigolactone Suicidal Germination Agents, towards Successful Striga Management

 ,
,  ,
,  , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
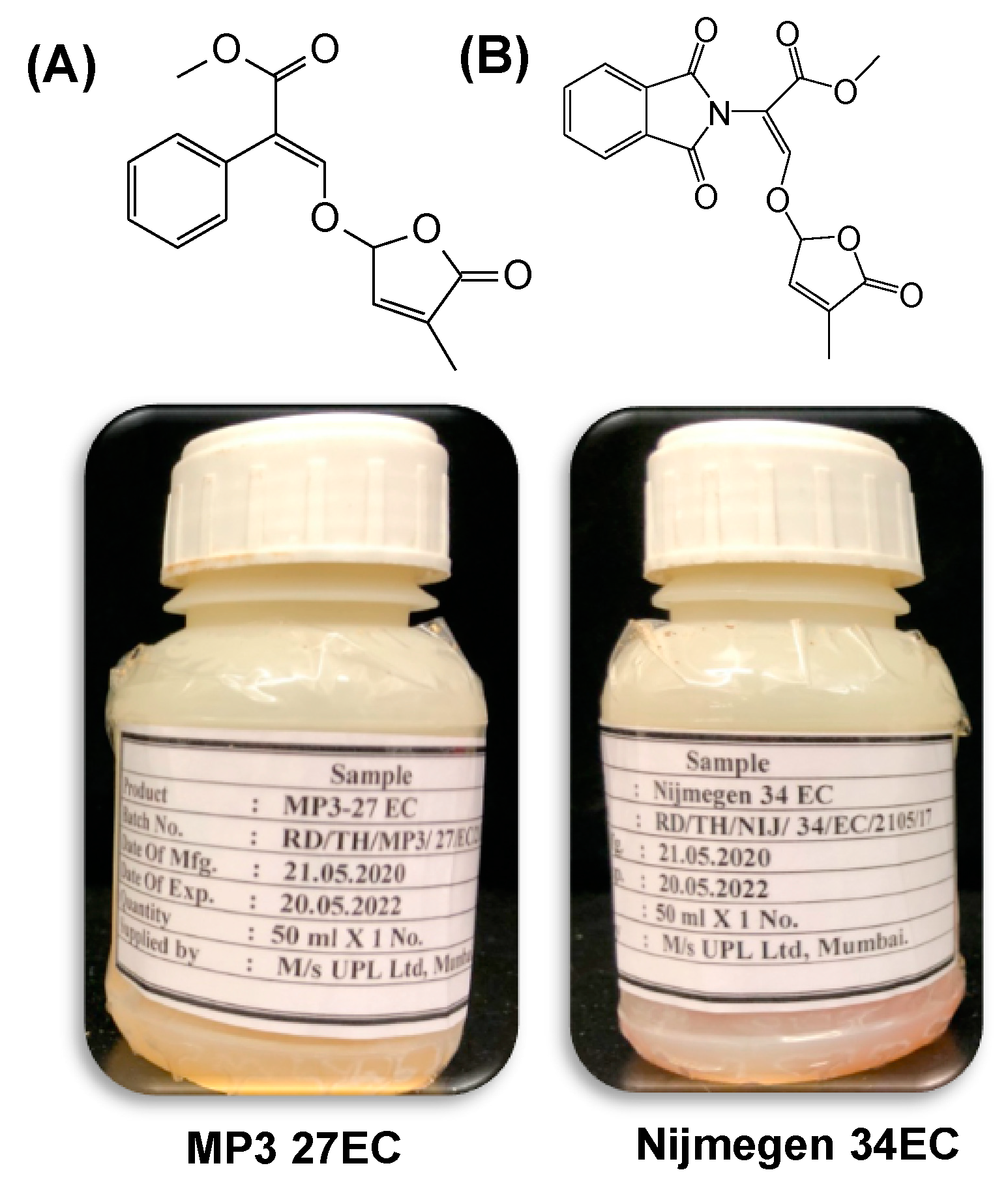
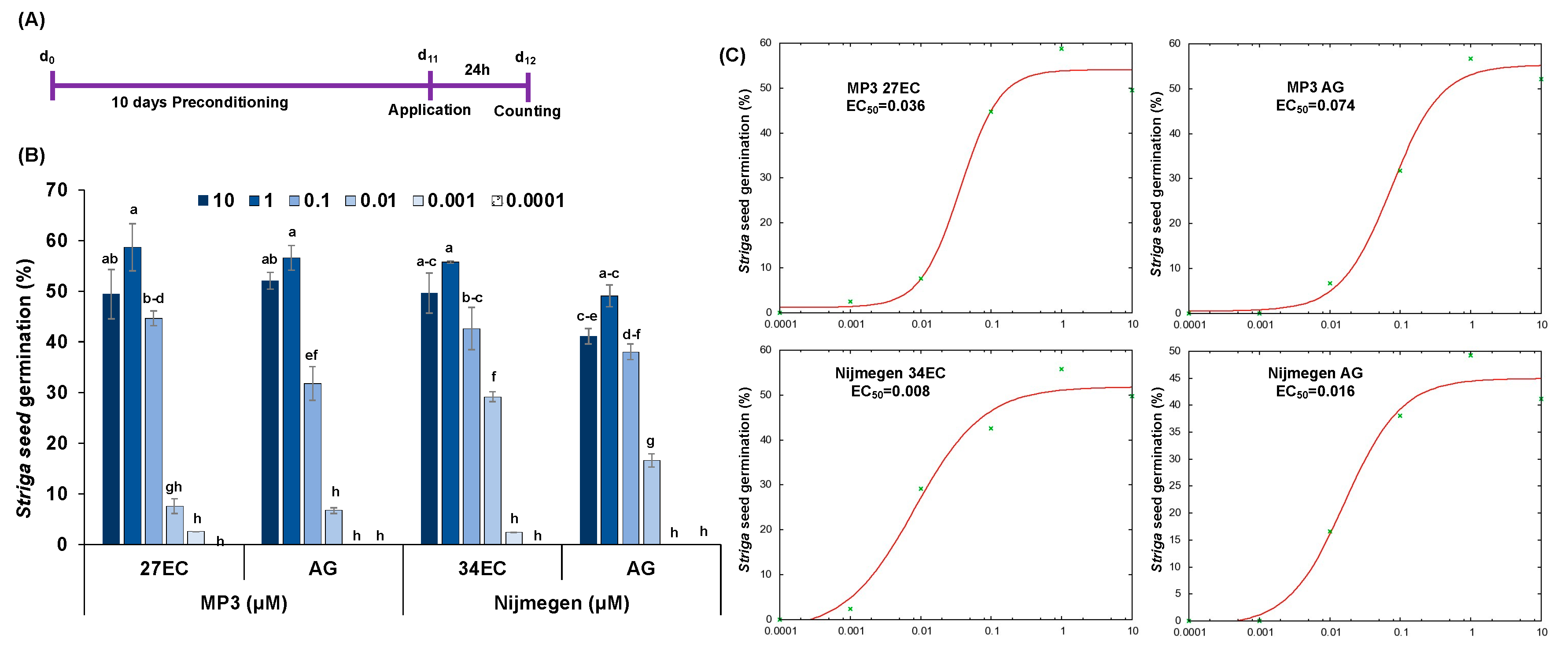
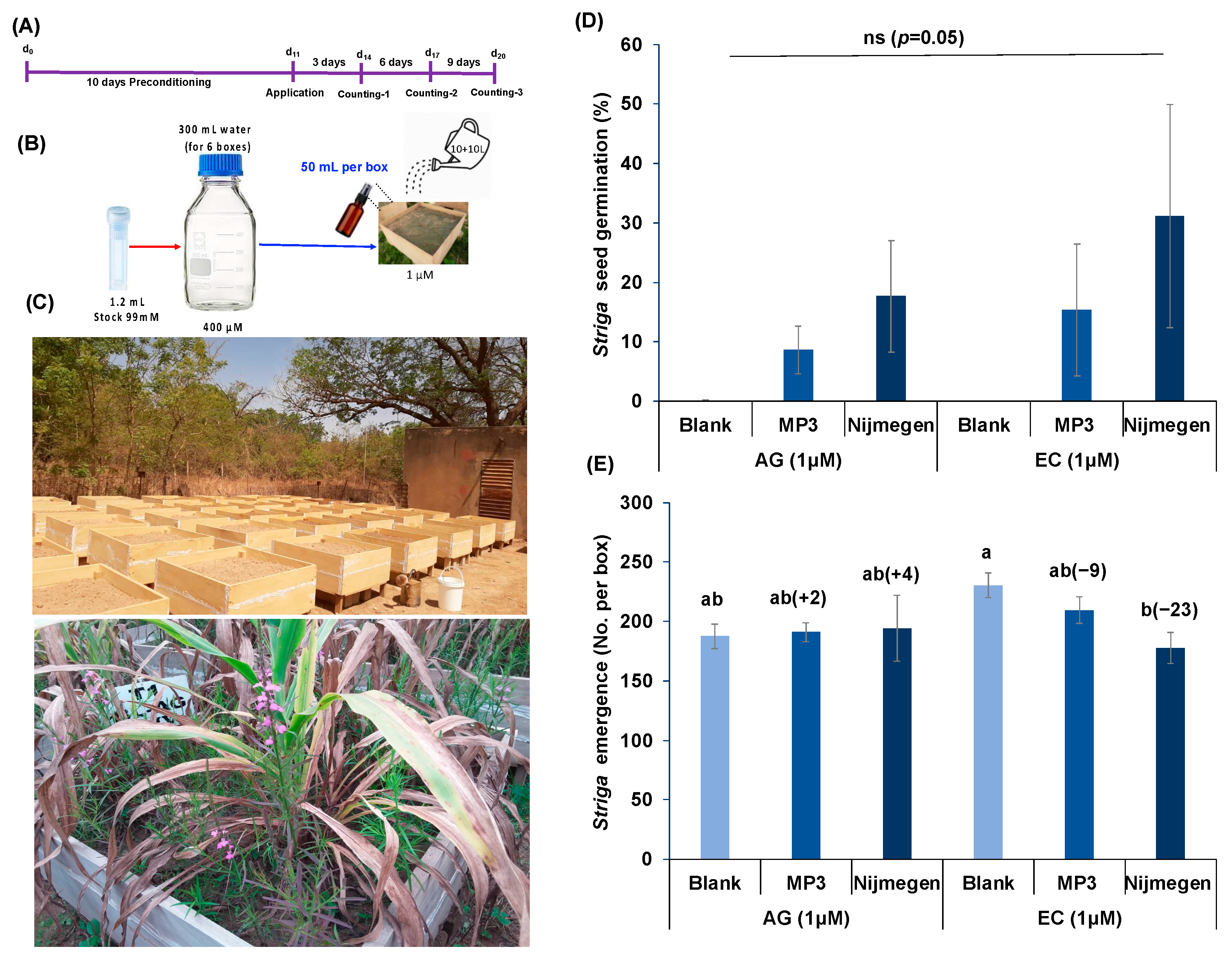
2.1. Striga Seed Germination in Response to EC and AG Formulations of SL Analogs under Lab Conditions
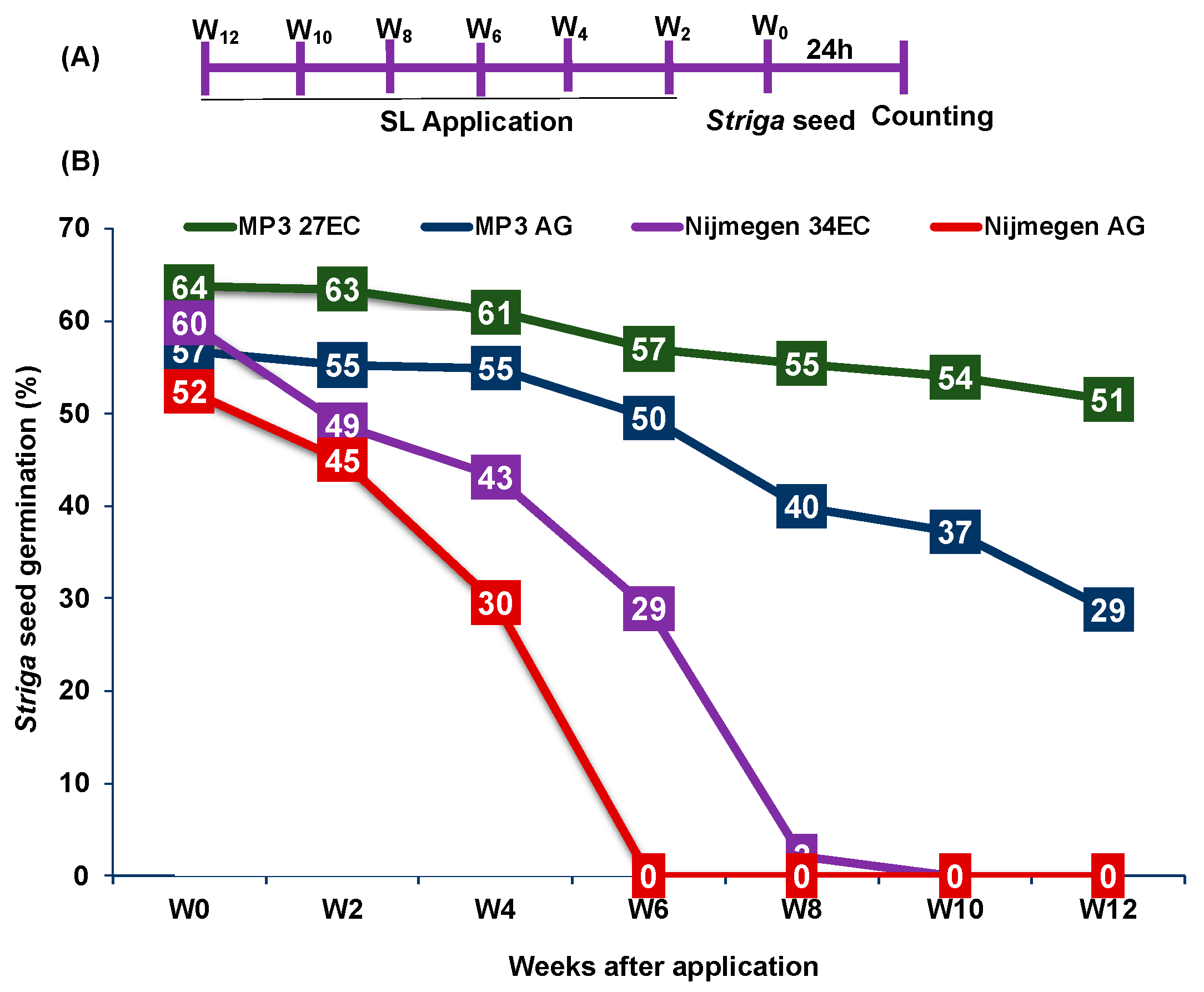
2.2. Stability of EC and AG Formulations of Strigolactone Analogs
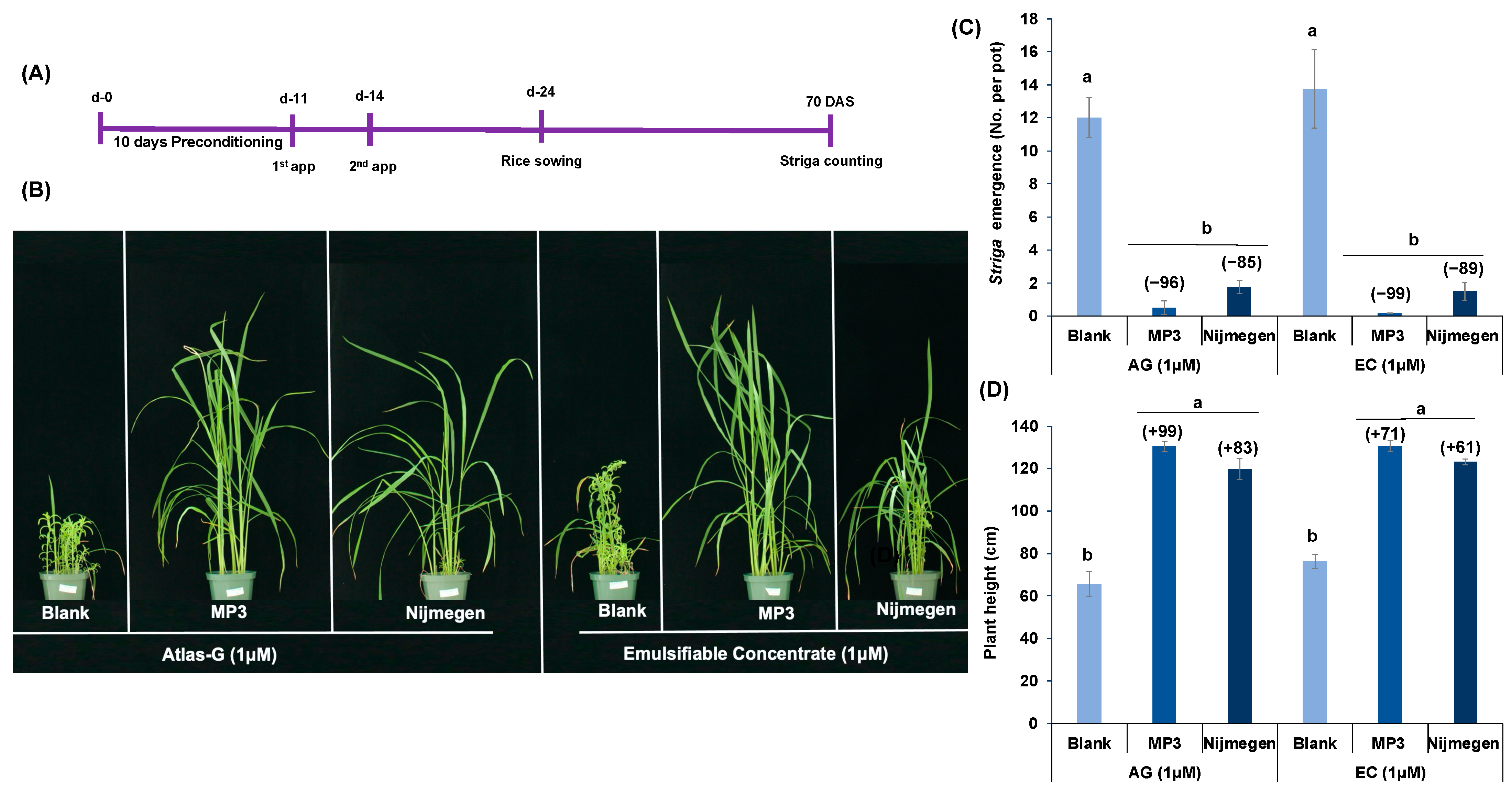
2.3. Effect of Various Formulations of Strigolactone Analogs on Striga Emergence in Pots under Greenhouse Conditions
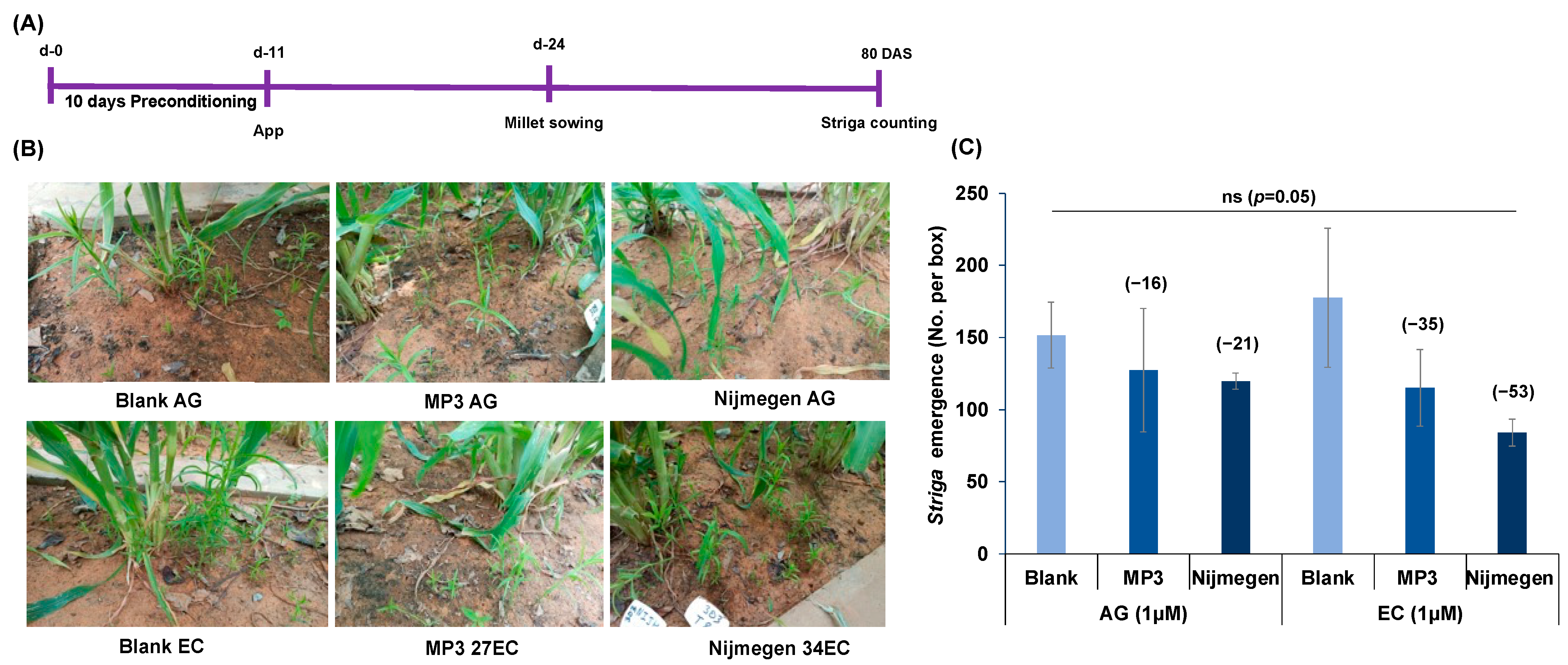
2.4. Effect of Various Formulations of Strigolactone Analogs on Striga Infection under Mini-Field Conditions
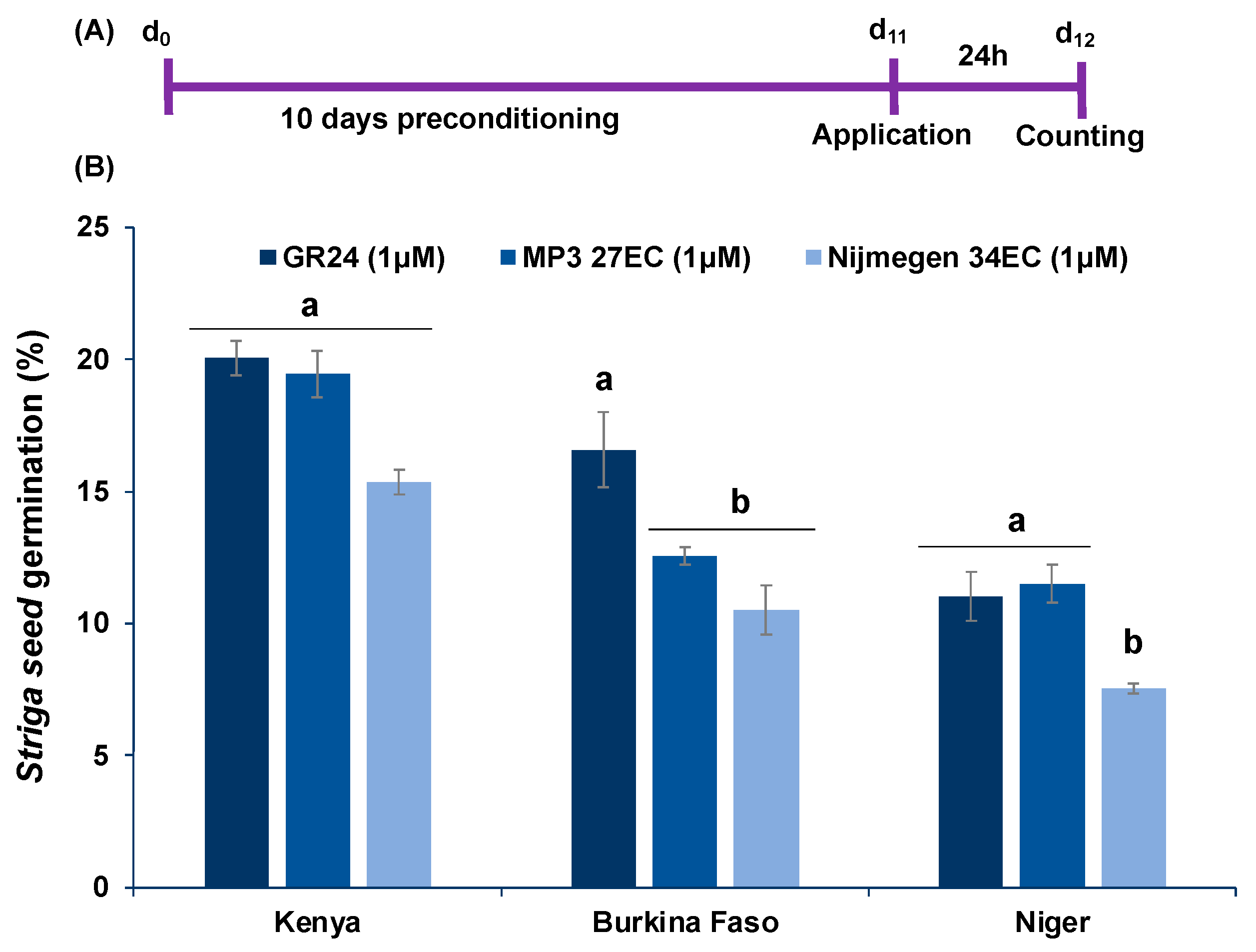
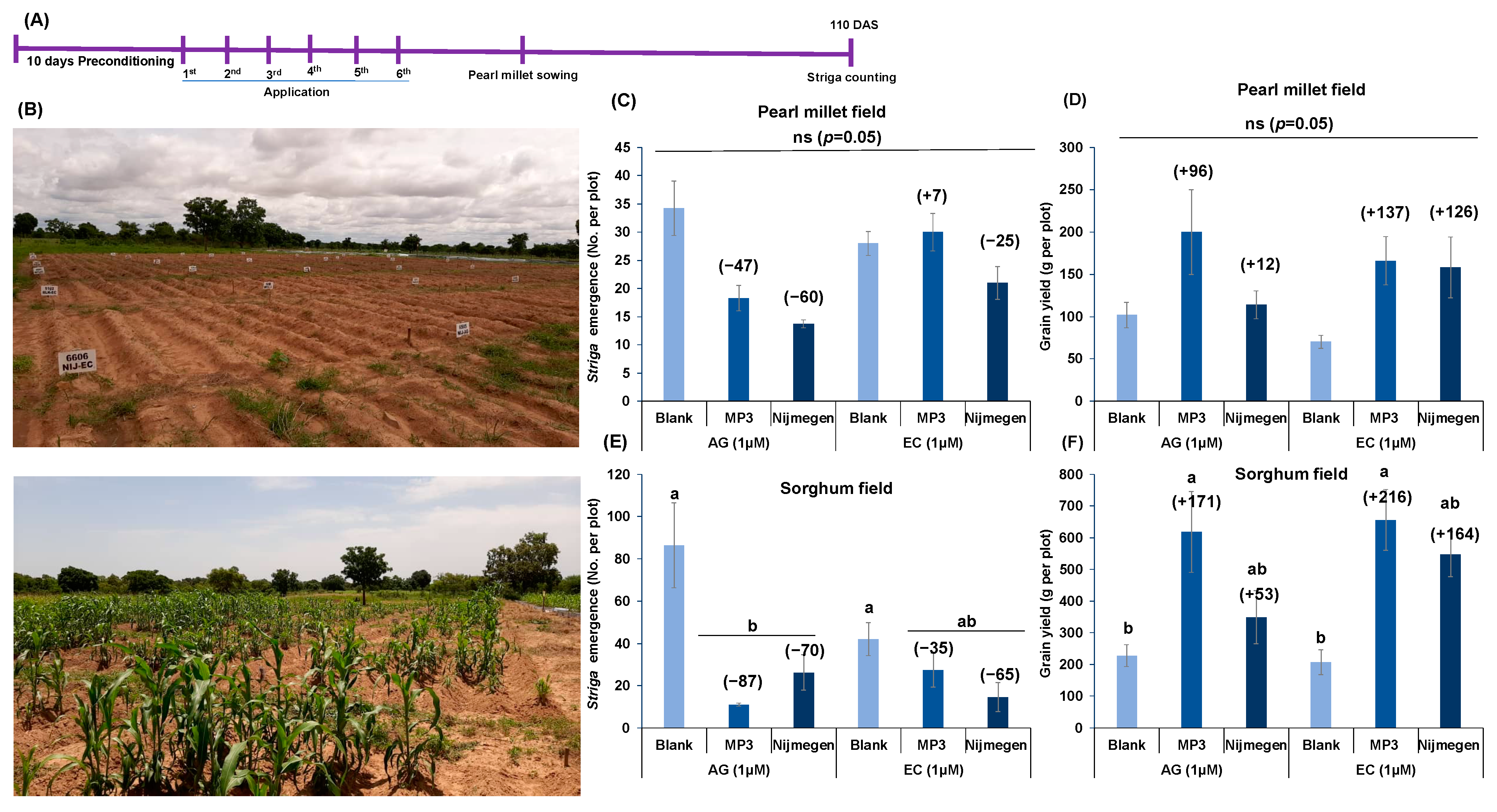
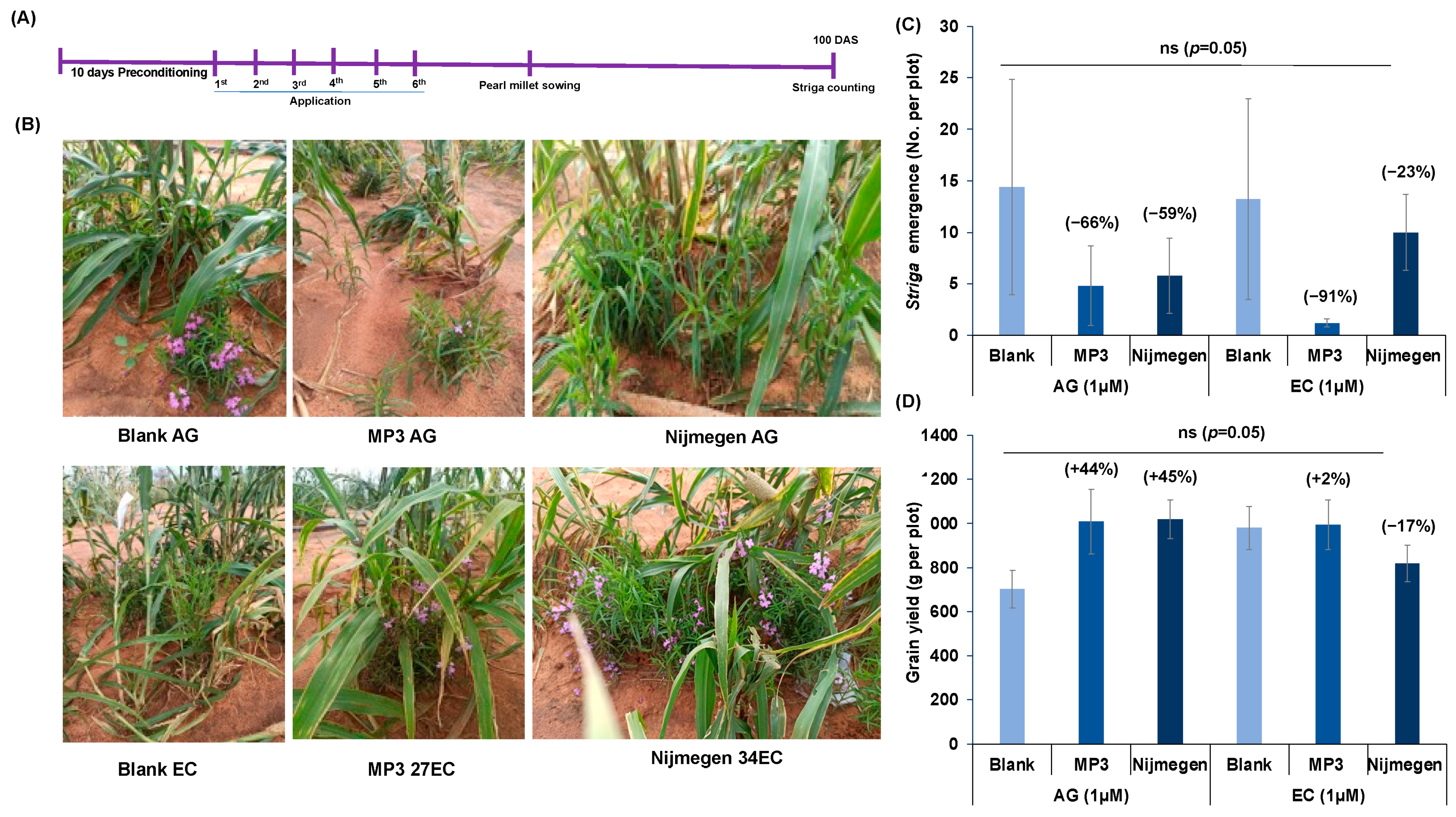
2.5. Effect of EC and AG Formulation of Strigolactone Analogs on Striga Infection under African Field Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemicals
4.2. Striga Seed Germination Bioassays
4.3. Stability Analysis
4.4. Striga Emergence in Pots under Greenhouse Conditions
4.5. Striga Emergence under Mini-Box Conditions
4.6. Striga Emergence under Field Conditions
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ejeta, G. Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host–Parasite Biology. Crop Sci. 2007, 47, 216–229. [Google Scholar] [CrossRef]
- Parker, C. Observations on the Current Status of Orobanche and Striga Problems Worldwide. Pest Manag. Sci. 2009, 65, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Spallek, T.; Mutuku, M.; Shirasu, K. The Genus Striga: A Witch Profile. Mol. Plant Path. 2013, 14, 861–869. [Google Scholar] [CrossRef]
- Jamil, M.; Kountche, B.A.; Al-Babili, S. Current Progress in Striga Management. Plant Physiol. 2021, 185, 1339–1352. [Google Scholar] [CrossRef] [PubMed]
- Scholes, J.D.; Press, M.C. Striga Infestation of Cereal Crops–an Unsolved Problem in Resource Limited Agriculture. Curr. Opin. Plant Biol. 2008, 11, 180–186. [Google Scholar] [CrossRef]
- Rodenburg, J.; Riches, C.R.; Kayeke, J.M. Addressing Current and Future Problems of Parasitic Weeds in Rice. Crop Prot. 2010, 29, 210–221. [Google Scholar] [CrossRef]
- Ejeta, G.; Gressel, J. Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt; World Scientific: Singapore, 2007; pp. 3–6. [Google Scholar]
- Atera, E.A.; Itoh, K.; Azuma, T.; Ishii, T. Farmers’ Perspectives on the Biotic Constraint of Striga hermonthica and its Control in Western Kenya. Weed Biol. Manag. 2012, 12, 53–62. [Google Scholar] [CrossRef]
- Pennisi, E. Armed and Dangerous. Science 2010, 327, 804–805. [Google Scholar] [PubMed] [Green Version]
- Pennisi, E. How Crop-Killing Witchweed Senses its Victims. Science 2015, 350, 146–147. [Google Scholar] [CrossRef]
- Rubiales, D.; Verkleij, J.; Vurro, M.; Murdoch, A.J.; Joel, D. Parasitic Plant Management in Sustainable Agriculture. Weed Res. 2009, 49, 1–5. [Google Scholar] [CrossRef]
- Berner, D.; Kling, J.; Singh, B. Striga Research and Control. A Perspective from Africa. Plant Dis. 1995, 79, 652–660. [Google Scholar] [CrossRef]
- Hearne, S.J. Control—the Striga Conundrum. Pest Manag. Sci. 2009, 65, 603–614. [Google Scholar] [CrossRef]
- Bebawi, F.F.; Eplee, R.E.; Harris, C.E.; Norris, R.S. Longevity of Witchweed (Striga asiatica) Seed. Weed Sci. 1984, 32, 494–497. [Google Scholar] [CrossRef]
- Abayo, G.O.; English, T.; Eplee, R.E.; Kanampiu, F.K.; Ransom, J.K.; Gressel, J. Control of Parasitic Witchweeds (Striga spp.) on Corn (Zea mays) Resistant to Acetolactate Synthase Inhibitors. Weed Sci. 1998, 46, 459–466. [Google Scholar] [CrossRef]
- Matusova, R.; Rani, K.; Verstappen, F.W.; Franssen, M.C.; Beale, M.H.; Bouwmeester, H.J. The Strigolactone Germination Stimulants of the Plant-Parasitic Striga and Orobanche spp. are Derived from the Carotenoid Pathway. Plant Physiol. 2005, 139, 920–934. [Google Scholar] [CrossRef] [Green Version]
- Matusova, R.; van Mourik, T.; Bouwmeester, H.J. Changes in the Sensitivity of Parasitic Weed Seeds to Germination Stimulants. Seed Sci. Res. 2004, 14, 335–344. [Google Scholar] [CrossRef]
- Al-Babili, S.; Bouwmeester, H.J. Strigolactones, a Novel Carotenoid-Derived Plant Hormone. Ann. Rev. Plant Biol. 2015, 66, 161–186. [Google Scholar] [CrossRef]
- Xie, X.N.; Yoneyama, K.; Yoneyama, K. The Strigolactone Story. Ann. Rev. Phytopath. 2010, 48, 93–117. [Google Scholar] [CrossRef] [Green Version]
- Fiorilli, V.; Wang, J.Y.; Bonfante, P.; Lanfranco, L.; Al-Babili, S. Apocarotenoids: Old and New Mediators of the Arbuscular Mycorrhizal Symbiosis. Front. Plant Sci. 2019, 10, 1186–1195. [Google Scholar] [CrossRef]
- Sibhatu, B. Review on Striga Weed Management. Int. J. Life Sci. Sci. Res. 2016, 2, 110–120. [Google Scholar]
- Zwanenburg, B.; Mwakaboko, A.S.; Kannan, C. Suicidal Germination for Parasitic Weed Control. Pest Manag. Sci. 2016, 72, 2016–2025. [Google Scholar] [CrossRef] [PubMed]
- Kountche, B.A.; Jamil, M.; Yonli, D.; Nikiema, M.P.; Blanco-Ania, D.; Asami, T.; Zwanenburg, B.; Al-Babili, S. Suicidal Germination as a Control Strategy for Striga hermonthica (Benth.) in Smallholder Farms of Sub-Saharan Africa. Plants People Planet 2019, 1, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, D.; Kuwata, K.; Hijikata, Y.; Yamaguchi, R.; Imaizumi, H.; Sathiyanarayanan, A.; Rakers, C.; Mori, N.; Akiyama, K.; Irle, S. A Femtomolar-Range Suicide Germination Stimulant for the Parasitic Plant Striga hermonthica. Science 2018, 362, 1301–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamil, M.; Kountche, B.A.; Wang, J.Y.; Haider, I.; Jia, K.-P.; Takahashi, I.; Ota, T.; Asami, T.; Al-Babili, S. A New Series of Carlactonoic Acid Based Strigolactone Analogs for Fundamental and Applied Research. Front. Plant Sci. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samejima, H.; Babiker, A.G.; Takikawa, H.; Sasaki, M.; Sugimoto, Y. Practicality of the Suicidal Germination Approach for Controlling Striga hermonthica. Pest Manag. Sci. 2016, 72, 2035–2042. [Google Scholar] [CrossRef] [PubMed]
- Mwakha, F.A.; Budambula, N.L.; Neondo, J.O.; Gichimu, B.M.; Odari, E.O.; Kamau, P.K.; Odero, C.; Kibet, W.; Runo, S. Witchweed’s Suicidal Germination: Can Slenderleaf Help? Agronomy 2020, 10, 873. [Google Scholar] [CrossRef]
- Parker, C.; Riches, C. Parasitic Weeds of the World: Biology and Control; CAB International: Wallingford, UK, 1993; pp. 1–74. [Google Scholar]
- Fukui, K.; Ito, S.; Asami, T. Selective Mimics of Strigolactone Actions and Their Potential Use for Controlling Damage Caused by Root Parasitic Weeds. Mol. Plant 2013, 6, 88–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco-Ania, D.; Mateman, J.J.; Hylova, A.; Spichal, L.; Debie, L.M.; Zwanenburg, B. Hybrid-type Strigolactone Analogues Derived from Auxins. Pest Manag. Sci. 2019, 75, 3113–3121. [Google Scholar] [CrossRef] [Green Version]
- Jamil, M.; Kountche, B.A.; Haider, I.; Wang, J.Y.; Aldossary, F.; Zarban, R.A.; Jia, K.-P.; Yonli, D.; Hameed, U.F.S.; Takahashi, I.; et al. Methylation at the C-3’ in D-Ring of Strigolactone Analogs Reduces Biological Activity in Root Parasitic Plants and Rice. Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jamil, M.; Kountche, B.A.; Haider, I.; Guo, X.J.; Ntui, V.O.; Jia, K.-P.; Ali, S.; Hameed, U.F.S.; Nakamura, H.; Lyu, Y.; et al. Methyl Phenlactonoates are Efficient Strigolactone Analogs with Simple Structure. J. Exp. Bot. 2018, 69, 2319–2331. [Google Scholar] [CrossRef]
- Jia, K.-P.; Kountche, B.A.; Jamil, M.; Guo, X.; Ntui, V.O.; Rüfenacht, A.; Rochange, S.; Al-Babili, S. Nitro-phenlactone, a Carlactone Analog with Pleiotropic Strigolactone Activities. Mol. Plant 2016, 9, 1341–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwanenburg, B.; Blanco-Ania, D. Strigolactones: New Plant Hormones in the Spotlight. J. Exp. Bot. 2018, 69, 2205–2218. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Charnikhova, T.; Verstappen, F.; Bouwmeester, H. Carotenoid Inhibitors Reduce Strigolactone Production and Striga hermonthica Infection in Rice. Arch. Biochem. Biophys. 2010, 504, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Adetimirin, V.; The, C.; Dossou, R. Yield Losses in Maize Due to Striga hermonthica in West and Central Africa. Int. J. Pest Manag. 2002, 48, 211–217. [Google Scholar] [CrossRef]
- Oswald, A. Striga Control—Technologies and Their Dissemination. Crop Prot. 2005, 24, 333–342. [Google Scholar] [CrossRef]
- Wigchert, S.; Kuiper, E.; Boelhouwer, G.; Nefkens, G.; Verkleij, J.; Zwanenburg, B. Dose−Response of Seeds of the Parasitic Weeds Striga and Orobanche toward the Synthetic Germination Stimulants GR24 and Nijmegen 1. J. Agric. Food Chem. 1999, 47, 1705–1710. [Google Scholar] [CrossRef]
- Yoneyama, K.; Awad, A.A.; Xie, X.; Yoneyama, K.; Takeuchi, Y. Strigolactones as Germination Stimulants for Root Parasitic Plants. Plant Cell Physiol. 2010, 51, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Kgosi, R.L.; Zwanenburg, B.; Mwakaboko, A.S.; Murdoch, A.J. Strigolactone Analogues Induce Suicidal Seed Germination of Striga spp. in Soil. Weed Res. 2012, 52, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Zwanenburg, B.; Mwakaboko, A.S.; Reizelman, A.; Anilkumar, G.; Sethumadhavan, D. Structure and Function of Natural and Synthetic Signalling Molecules in Parasitic Weed Germination. Pest Manag. Sci. 2009, 65, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Kanampiu, F.K.; Karaya, H.; Charnikhova, T.; Bouwmeester, H.J. Striga hermonthica Parasitism in Maize in Response to N and P Fertilisers. Field Crops Res. 2012, 134, 1–10. [Google Scholar] [CrossRef]
- Braguy, J.; Ramazanova, M.; Giancola, S.; Jamil, M.; Kountche, B.A.; Zarban, R.A.Y.; Felemban, A.; Wang, J.Y.; Lin, P.-Y.; Haider, I.; et al. SeedQuant: A Deep Learning-Based Tool for Assessing Stimulant and Inhibitor Activity on Root Parasitic Seeds. Plant Physiol. 2021, 186, 1632–1644. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Jamil, M.; Lin, P.-Y.; Ota, T.; Fiorilli, V.; Novero, M.; Zarban, R.A.; Kountche, B.A.; Takahashi, I.; Martínez, C. Efficient Mimics for Elucidating Zaxinone Biology and Promoting Agricultural Applications. Mol. Plant 2020, 13, 1654–1661. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamil, M.; Wang, J.Y.; Yonli, D.; Patil, R.H.; Riyazaddin, M.; Gangashetty, P.; Berqdar, L.; Chen, G.-T.E.; Traore, H.; Margueritte, O.; et al. A New Formulation for Strigolactone Suicidal Germination Agents, towards Successful Striga Management. Plants 2022, 11, 808. https://doi.org/10.3390/plants11060808
Jamil M, Wang JY, Yonli D, Patil RH, Riyazaddin M, Gangashetty P, Berqdar L, Chen G-TE, Traore H, Margueritte O, et al. A New Formulation for Strigolactone Suicidal Germination Agents, towards Successful Striga Management. Plants. 2022; 11(6):808. https://doi.org/10.3390/plants11060808
Chicago/Turabian StyleJamil, Muhammad, Jian You Wang, Djibril Yonli, Rohit H. Patil, Mohammed Riyazaddin, Prakash Gangashetty, Lamis Berqdar, Guan-Ting Erica Chen, Hamidou Traore, Ouedraogo Margueritte, and et al. 2022. "A New Formulation for Strigolactone Suicidal Germination Agents, towards Successful Striga Management" Plants 11, no. 6: 808. https://doi.org/10.3390/plants11060808