
Identification of Genes in Xanthomonas euvesicatoria pv. rosa That Are Host Limiting in Tomato

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Whole-Genome Sequence Similarity
2.2. Type III Secreted Effectors Repertoires
2.3. xer3856 as Host-Limiting Candidate
2.4. XopA and Xer3856 Are Host-Limiting Factors in Tomato
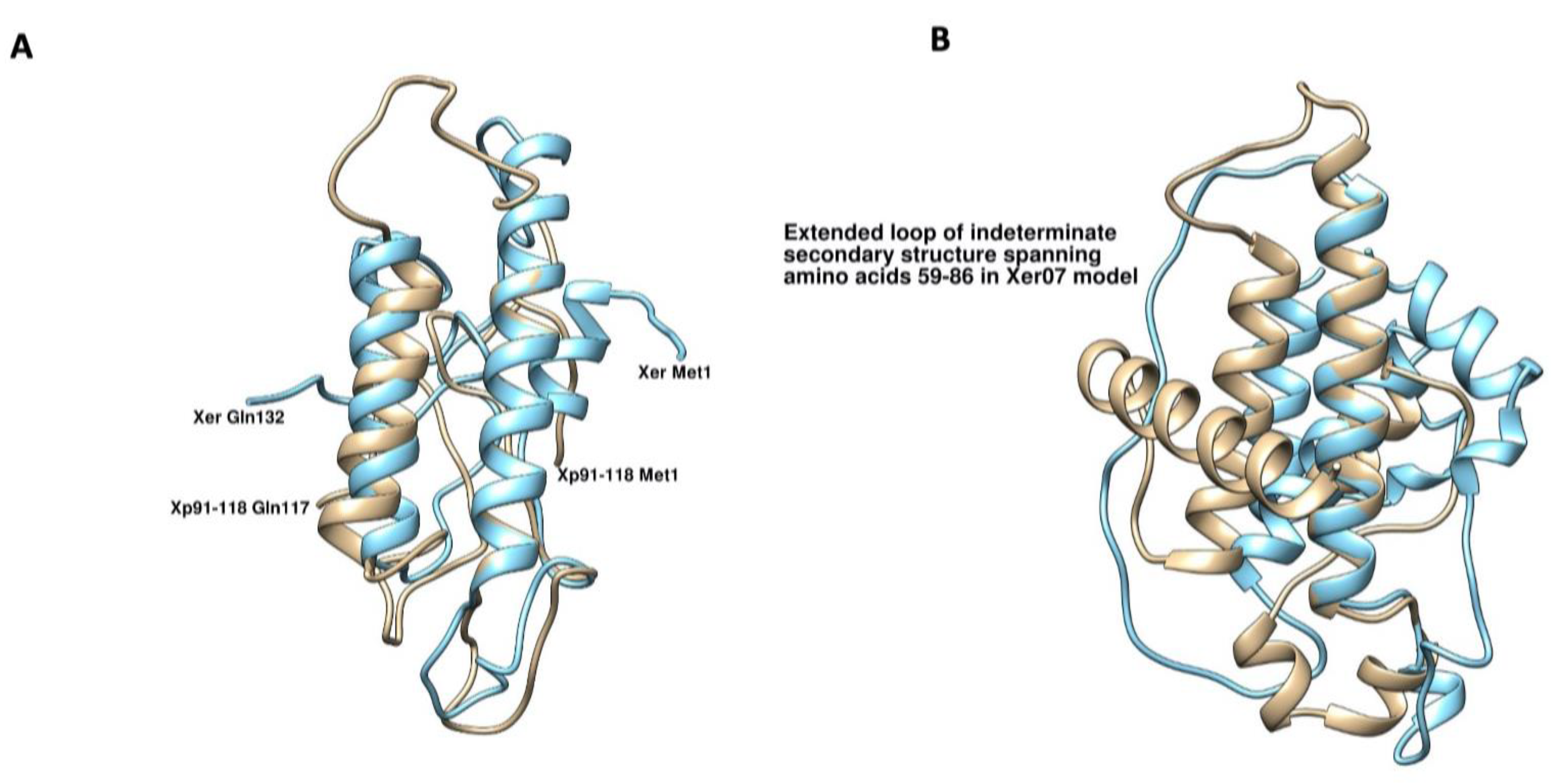
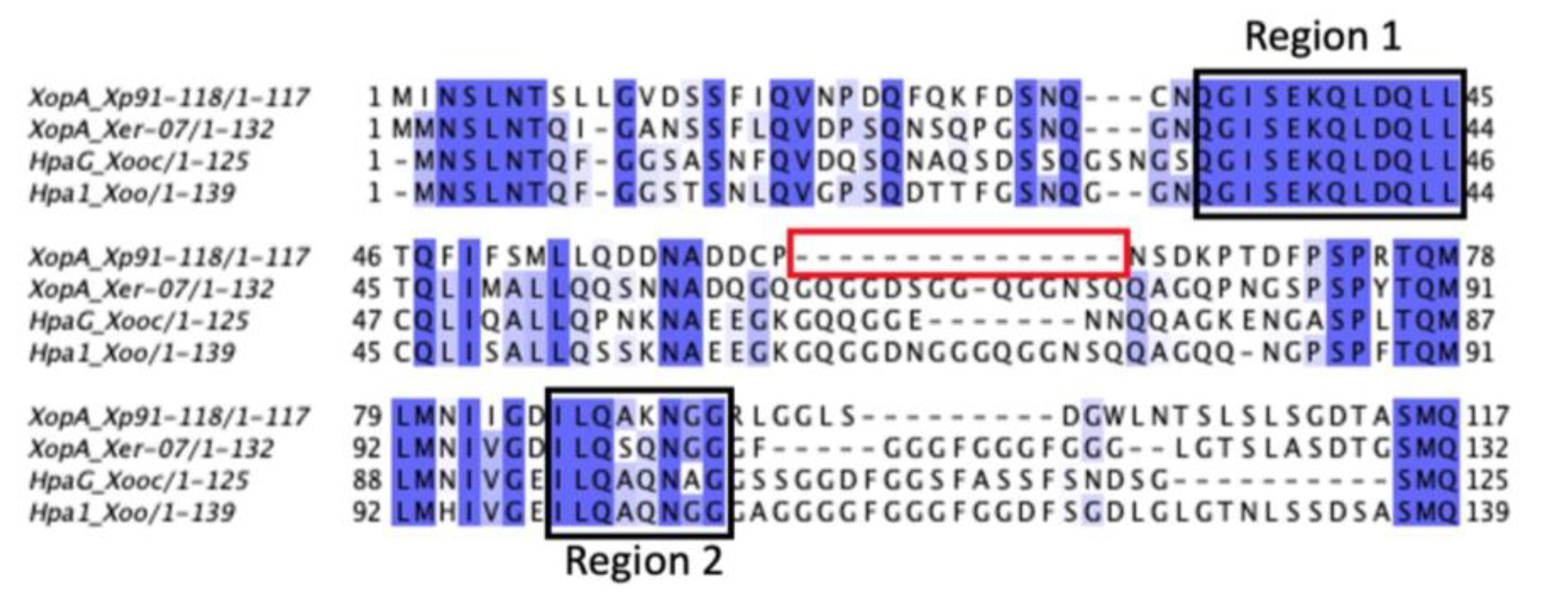
2.5. Structure of XopA
2.6. Comparative Genomics of Other Pathogenicity Associated Factors
3. Discussion
4. Material and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Genome Collection and Genome Statistics
4.3. Effector Repertoire and Pathogenicity Associated Genes
4.4. Mutants, Transconjugants and Population Dynamics
4.5. Comparative Genomics
4.6. Pathogenicity Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- USDA National Agricultural Statistics Service. Floriculture Crops 2020 Summary; USDA National Agricultural Statistics Service: Washington, DC, USA, 2021. [Google Scholar]
- Huang, C.-H.; Vallad, G.E.; Adkison, H.; Summers, C.; Margenthaler, E.; Schneider, C.; Hong, J.; Jones, J.B.; Ong, K.; Norman, D.J. A Novel Xanthomonas sp. Causes Bacterial Spot of Rose (Rosa spp.). Plant Dis. 2013, 97, 1301–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barak, J.D.; Vancheva, T.; Lefeuvre, P.; Jones, J.B.; Timilsina, S.; Minsavage, G.V.; Vallad, G.E.; Koebnik, R. Whole-Genome Sequences of Xanthomonas euvesicatoria Strains Clarify Taxonomy and Reveal a Stepwise Erosion of Type 3 Effectors. Front. Plant Sci. 2016, 7, 1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rademaker, J.L.; Louws, F.; Schultz, M.H.; Rossbach, U.; Vauterin, L.; Swings, J.; De Bruijn, F.J. A Comprehensive Species to Strain Taxonomic Framework for Xanthomonas. Phytopathology 2005, 95, 1098–1111. [Google Scholar] [CrossRef] [Green Version]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant–pathogen interactions. Nat. Rev. Genet. 2020, 18, 415–427. [Google Scholar] [CrossRef]
- He, S.Y.; Nomura, K.; Whittam, T.S. Type III protein secretion mechanism in mammalian and plant pathogens. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1694, 181–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buüttner, D.; He, S.Y. Type III Protein Secretion in Plant Pathogenic Bacteria. Plant Physiol. 2009, 150, 1656–1664. [Google Scholar] [CrossRef] [Green Version]
- Astua-Monge, G.; Minsavage, G.V.; Stall, R.E.; Davis, M.J.; Bonas, U.; Jones, J.B. Resistance of Tomato and Pepper to T3 Strains of Xanthomonas campestris pv. Vesicatoria Is Specified by a Plant-Inducible Avirulence Gene. Mol. Plant-Microbe Interact. 2000, 13, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Minsavage, G.; Dahlbeck, D.; Whalen, M.; Kearney, B.; Bonas, U.; Staskawicz, B.; Stall, R. Gene-for-gene relationships specifying disease resistance in Xanthomonas campestris pv. vesicatoria-pepper interactions. Mol. Plant-Microbe Interact. 1990, 3, 41–47. [Google Scholar] [CrossRef]
- Schwartz, A.R.; Potnis, N.; Timilsina, S.; Wilson, M.; Patané, J.; Martins, J.J.; Minsavage, G.V.; Dahlbeck, D.; Akhunova, A.; Almeida, N.; et al. Phylogenomics of Xanthomonas field strains infecting pepper and tomato reveals diversity in effector repertoires and identifies determinants of host specificity. Front. Microbiol. 2015, 6, 535. [Google Scholar] [CrossRef] [Green Version]
- Potnis, N.; Krasileva, K.; Chow, V.; Almeida, N.F.; Patil, P.B.; Ryan, R.P.; Sharlach, M.; Behlau, F.; Dow, J.M.; Momol, M.; et al. Comparative genomics reveals diversity among xanthomonads infecting tomato and pepper. BMC Genom. 2011, 12, 146. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, N. Genome-Wide Mutagenesis of Xanthomonas axonopodis pv. citri Reveals Novel Genetic Determinants and Regulation Mechanisms of Biofilm Formation. PLoS ONE 2011, 6, e21804. [Google Scholar] [CrossRef] [Green Version]
- Rajeshwari, R.; Jha, G.; Sonti, R.V. Role of an In Planta-Expressed Xylanase of Xanthomonas oryzae pv. oryzae in Promoting Virulence on Rice. Mol. Plant-Microbe Interact. 2005, 18, 830–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczesny, R.; Jordan, M.; Schramm, C.; Schulz, S.; Cogez, V.; Bonas, U.; Büttner, D. Functional characterization of the Xcs and Xps type II secretion systems from the plant pathogenic bacterium Xanthomonas campestris pv vesicatoria. New Phytol. 2010, 187, 983–1002. [Google Scholar] [CrossRef] [PubMed]
- Steffens, T.; Duda, K.; Lindner, B.; Vorhölter, F.-J.; Bednarz, H.; Niehaus, K.; Holst, O. The lipopolysaccharide of the crop pathogen Xanthomonas translucenspv. translucens: Chemical characterization and determination of signaling events in plant cells. Glycobiology 2017, 27, 264–274. [Google Scholar] [CrossRef]
- Dueholm, M.S. Biophysical Characterization of Naturally Occurring Bacterial Amyloids. Ph.D. Thesis, Aalborg University, Aalborg, Denmark, 2010. [Google Scholar]
- Shanmugam, N.; Baker, M.O.D.G.; Ball, S.R.; Steain, M.; Pham, C.L.L.; Sunde, M. Microbial functional amyloids serve diverse purposes for structure, adhesion and defence. Biophys. Rev. 2019, 11, 287–302. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, L.; Ji, H.; Mo, X.; Li, P.; Wang, J.; Dong, H. Hpa1 is a type III translocator in Xanthomonas oryzae pv. oryzae. BMC Microbiol. 2018, 18, 105. [Google Scholar] [CrossRef]
- Kim, J.-G.; Taylor, K.W.; Hotson, A.; Keegan, M.; Schmelz, E.A.; Mudgett, M.B. XopD SUMO Protease Affects Host Transcription, Promotes Pathogen Growth, and Delays Symptom Development in Xanthomonas-Infected Tomato Leaves. Plant Cell 2008, 20, 1915–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bibi, S.; Pereira, J.; Timilsina, S.; Minsavage, G.V.; Hulbert, J.C.; Jones, J.B. 2018. Examining The Role of an EF-Hand Protein in Regulating Virulence in Xanthomonas. In Proceedings of the International Congress of Plant Pathology (ICPP 2018), Boston, MA, USA, 29 July–3 August 2018. [Google Scholar]
- Charkowski, A.O.; Alfano, J.R.; Preston, G.; Yuan, J.; He, S.Y.; Collmer, A. The Pseudomonas syringae pv. tomato HrpW Protein Has Domains Similar to Harpins and Pectate Lyases and Can Elicit the Plant Hypersensitive Response and Bind to Pectate. J. Bacteriol. 1998, 180, 5211–5217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, J. Genetic and molecular analysis of pathogenicity genes in Xanthomonas citri subsp. citri. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2009. [Google Scholar]
- White, F.F.; Potnis, N.; Jones, J.B.; Koebnik, R. The type III effectors of Xanthomonas. Mol. Plant Pathol. 2009, 10, 749–766. [Google Scholar] [CrossRef]
- Noël, L.; Thieme, F.; Nennstiel, D.; Bonas, U. Two Novel Type III-Secreted Proteins of Xanthomonas campestris pv. vesicatoria Are Encoded within the hrp Pathogenicity Island. J. Bacteriol. 2002, 184, 1340–1348. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-G.; Park, B.K.; Yoo, C.-H.; Jeon, E.; Oh, J.; Hwang, I. Characterization of the Xanthomonas axonopodis pv. glycines Hrp Pathogenicity Island. J. Bacteriol. 2003, 185, 3155–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, Y.; Wang, S.; Fang, A.; Wang, J.; Liu, L.; Zhang, K.; Mao, Y.; Sun, W. The Type III Effector AvrBs2 in Xanthomonas oryzae pv. oryzicola Suppresses Rice Immunity and Promotes Disease Development. Mol. Plant-Microbe Interact. 2015, 28, 869–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Zhang, L.; Mo, X.; Ji, H.; Bian, H.; Hu, Y.; Majid, T.; Long, J.; Pang, H.; Tao, Y.; et al. Rice aqua-porin PIP1;3 and harpin Hpa1 of bacterial blight pathogen cooperate in a type III effector translocation. J. Exp. Bot. 2019, 70, 3057–3073. [Google Scholar] [CrossRef] [Green Version]
- Rybak, M.; Minsavage, G.V.; Stall, R.E.; Jones, J.B. Identification of Xanthomonas citri ssp. citri host specificity genes in a heterologous expression host. Mol. Plant Pathol. 2009, 10, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Chosed, R.; Tomchick, D.; Brautigam, C.; Mukherjee, S.; Negi, V.S.; Machius, M.; Orth, K. Structural Analysis of Xanthomonas XopD Provides Insights into Substrate Specificity of Ubiquitin-like Protein Proteases. J. Biol. Chem. 2007, 282, 6773–6782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auch, A.F.; Von Jan, M.; Klenk, H.-P.; Göker, M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand. Genom. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [Green Version]
- Delcher, A.; Phillippy, A.; Carlton, J.; Salzberg, S. Fast algorithms for large-scale genome alignment and comparison. Nucleic Acids Res. 2002, 30, 2478–2483. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Aranda, J.; Poza, M.; Pardo, B.G.; Rumbo, S.; Rumbo, C.; Parreira, J.R.; Rodríguez-Velo, P.; Bou, G. A rapid and simple method for constructing stable mutants of Acinetobacter baumannii. BMC Microbiol. 2010, 10, 279. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Host Name | GenBank Accession No. | Total Sequence Length (bp) | GC Content (%) | Gene Count |
|---|---|---|---|---|---|
| X. euvesicatoria pv. rosa GEV-Rose-07 | Rosa sp. | GCA_001855615.1 | 4,970,862 | 64.87 | 4510 |
| X. euvesicatoria pv. allii CFBP 6369 | Allium cepa (onion) | GCA_000730305.1 | 5,427,242 | 64.35 | 4696 |
| X. alfalfae subsp. citrumelonis F1 | Rutaceae citrus (citrus) | GCA_000225915.1 | 4,967,469 | 64.92 | 4188 |
| X. euvesicatoria LMG 12749 | Philodendron | GCA_001401675.2 | 4,886,158 | 64.91 | 4388 |
| X. perforans 91-118 | S. lycopersicum (tomato) | GCA_000192045.3 | 4,898,349 | 65.04 | 4186 |
| X. euvesicatoria 85-10 | C. annuum (pepper) | GCA_000009165.1 | 5,420,152 | 64.56 | 4707 |
| X. gardneri ATCC 19865 | Tomato | GCA_000192065.2 | 5,528,124 | 63.68 | 5424 |
| X. vesicatoria ATCC 35937 | Tomato | GCA_000192025.2 | 5,531,089 | 64.07 | 5117 |
| Effectors | Synonyms | Xer07 | Xe 85-10 | Xp 91-118 | Xac F1 | Xea CFBP6369 |
|---|---|---|---|---|---|---|
| AvrBs2 | + 1 | 704/714 (99%) 2 | 708/714 (99%) | 708/714 (99%) | 710/714 (99%) | |
| XopE1 | avrXacE1 | + | 397/400 (99%) | 394/400 (99%) | 393/400 (98%) | 396/400 (99%) |
| HpaA | + | 273/275 (99%) | 272/275 (99%) | 272/275 (99%) | 271/275 (99%) | |
| XopA | Hpa1 | + | 69/133 (52%) | 68/133 (51%) | 132/132 (100%) | 132/132 (100%) |
| XopAD | + | 510/530 (96%) | 530/530 (100%) | 463/531 (87%) | 461/530 (87%) | |
| XopAE | HpaF/G | + | − 3 | 644/650 (99%) | 632/650 (97%) | 643/650 (99%) |
| XopAP | + | 417/423 (99%) | 417/423 (99%) | 424/427 (99%) | 419/423 (99%) | |
| XopAU | + | 515/517 (99%) | 513/517 (99%) | 511/517 (99%) | 513/517 (99%) | |
| XopAW | + | 216/221 (98%) | 221/221 (100%) | 220/221 (99%) | 221/221 (100%) | |
| XopC2 | + | 206/206 (100%) &196/200 (98%) 4 | 432/437 (99%) | 432/437 (99%) | 434/437 (99%) | |
| XopF1 | Hpa4 | + | 664/670 (99%) | 661/672 (98%) | 668/670 (99%) | 666/670 (99%) |
| XopF2 | + | 653/667 (98%) | 662/667 (99%) | 660/667 (99%) | 639/647 (99%) | |
| XopK | + | 630/634 (99%) | 634/634 (100%) | 614/634 (97%) | 614/635 (97%) | |
| XopL | + | 634/661 (96%) | 641/661 (97%) | 633/661 (96%) | 639/661 (97%) | |
| XopN | + | 718/733 (98%) | 731/733 (99%) | 724/733 (99%) | 728/733 (99%) | |
| XopQ | + | 460/464 (99%) | 460/464 (99%) | 459/464 (99%) | 456/464 (98%) | |
| XopR | + | 399/404 (99%) | 400/404 (99%) | 401/404 (99%) | 399/404 (99%) | |
| XopS | + | 298/308 (97%) | 301/308 (98%) | 295/307 (96%) | 302/307 (98%) | |
| XopV | + | 338/346 (98%) | 339/346 (98%) | 341/346 (98%) | 342/346 (99%) | |
| XopX | + | 688/721 (95%) | 683/714 (96%) | 680/714 (95%) | 683/721 (95%) | |
| XopZ | + | 1376/1388 (99%) | 1369/1388 (99%) | 1372/1388 (99%) | 1377/1388 (99%) | |
| XopP | + | 567/577 (98%) +50/51 (98%) | 622/627 (99%) | 617/627 (98%) | 629/641 (98%) | |
| XopAF | avrXv3 | + | − | 217/218 (99%) | − | 217/218 (99%) |
| XopI | + | 443/450 (98%) | 443/450 (98%) | 444/450 (99%) | 443/450 (98%) | |
| XopE2 | avrXacE3 /avrXccE1 | + | 352/358 (98%) | − | − | − |
| XopAJ | avrRxo1 | + | 412/421 (98%) | − | 320/324 (99%) | − |
| Strain | Characteristics | Source |
|---|---|---|
| Xer07 | Xanthomonas strain isolated from Rose | This study |
| Xp 91-118 | X. perforans isolated from tomato | This study |
| Xe 85-10 | X. euvesicatoria isolated from pepper | This study |
| E. coli DH5α | Competent cell for hosting the plasmid | Bethesda Research Laboratories |
| Xer07Ωxer3856 (TOPO-xer3856) | Xer07, xer3856 mutated using TOPO, KanR | This study |
| Xp 91-118::xer3856 (pUFR034::xer3856) | Xp 91-118 complemented with xer3856, KanR | This study |
| Xp 91-118::xopA (pUFR034::xopA) | Xp 91-118, complemented with xopA from Xer 07, KanR | This study |
| Plasmids | ||
| pCR2.1-TOPO | TOPO plasmid, KanR | Thermofisher |
| pCR2.1-TOPO-xer3856 | PCR product of xer3856-int-F and xer3856-int-R into pCR2.1-TOPO, KanR | This study |
| pUFR034 | Cloning vector, KanR | |
| pUFR034-xer3856 | PCR product of xer3856-out-F and xer3856-out-R into pUFR034, KanR | This study |
| pUFR034-xopA | PCR product of xopA-F and xopA-R from Xer07 into pUFR034, KanR | This study |
| pGEM-T Easy | Cloning vector, AmpR | Promega |
| pGEM-T Easy-xer3856 | xer3856 in pGEM-T for cloning, AmpR | This study |
| pGEM-T Easy-xopA | xopA from Xer07 in pGEM-T for cloning to pUFR034-xopA, AmpR | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Q.; Bibi, S.; Vallad, G.E.; Goss, E.M.; Hurlbert, J.C.; Paret, M.L.; Jones, J.B.; Timilsina, S. Identification of Genes in Xanthomonas euvesicatoria pv. rosa That Are Host Limiting in Tomato. Plants 2022, 11, 796. https://doi.org/10.3390/plants11060796
Fan Q, Bibi S, Vallad GE, Goss EM, Hurlbert JC, Paret ML, Jones JB, Timilsina S. Identification of Genes in Xanthomonas euvesicatoria pv. rosa That Are Host Limiting in Tomato. Plants. 2022; 11(6):796. https://doi.org/10.3390/plants11060796
Chicago/Turabian StyleFan, Qiurong, Shaheen Bibi, Gary E. Vallad, Erica M. Goss, Jason C. Hurlbert, Matthews L. Paret, Jeffrey B. Jones, and Sujan Timilsina. 2022. "Identification of Genes in Xanthomonas euvesicatoria pv. rosa That Are Host Limiting in Tomato" Plants 11, no. 6: 796. https://doi.org/10.3390/plants11060796