
Effectiveness of Leaf Rust Resistance Genes in the Adult and Juvenile Stages in Southern Russia in 2011–2020




Abstract
:1. Introduction
2. Results
2.1. Efficiency of Lr Genes in Adult Phase
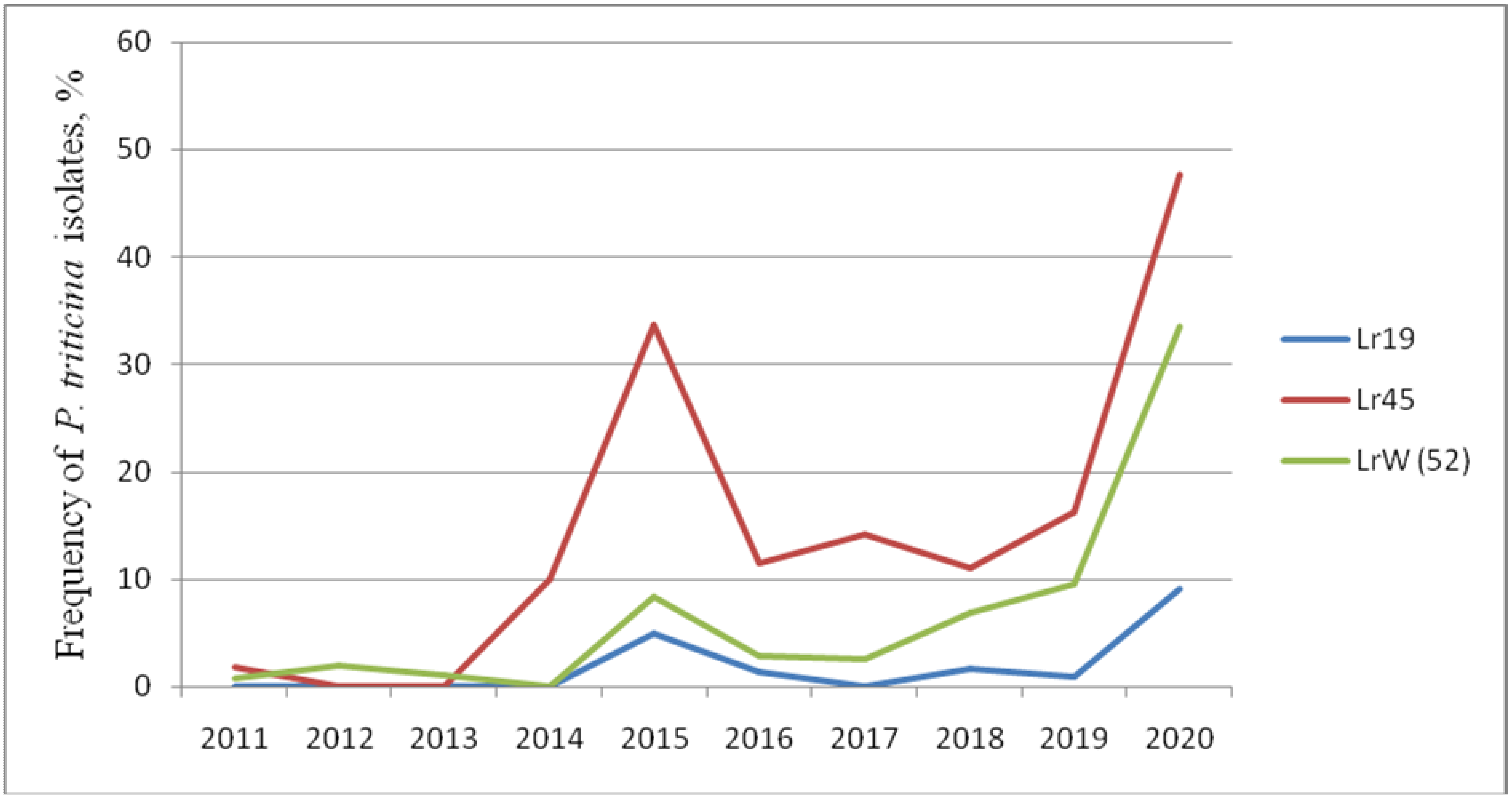
2.2. Efficiency of Lr Genes in Juvenile Phase
3. Discussion
4. Conclusions
5. Materials and Methods
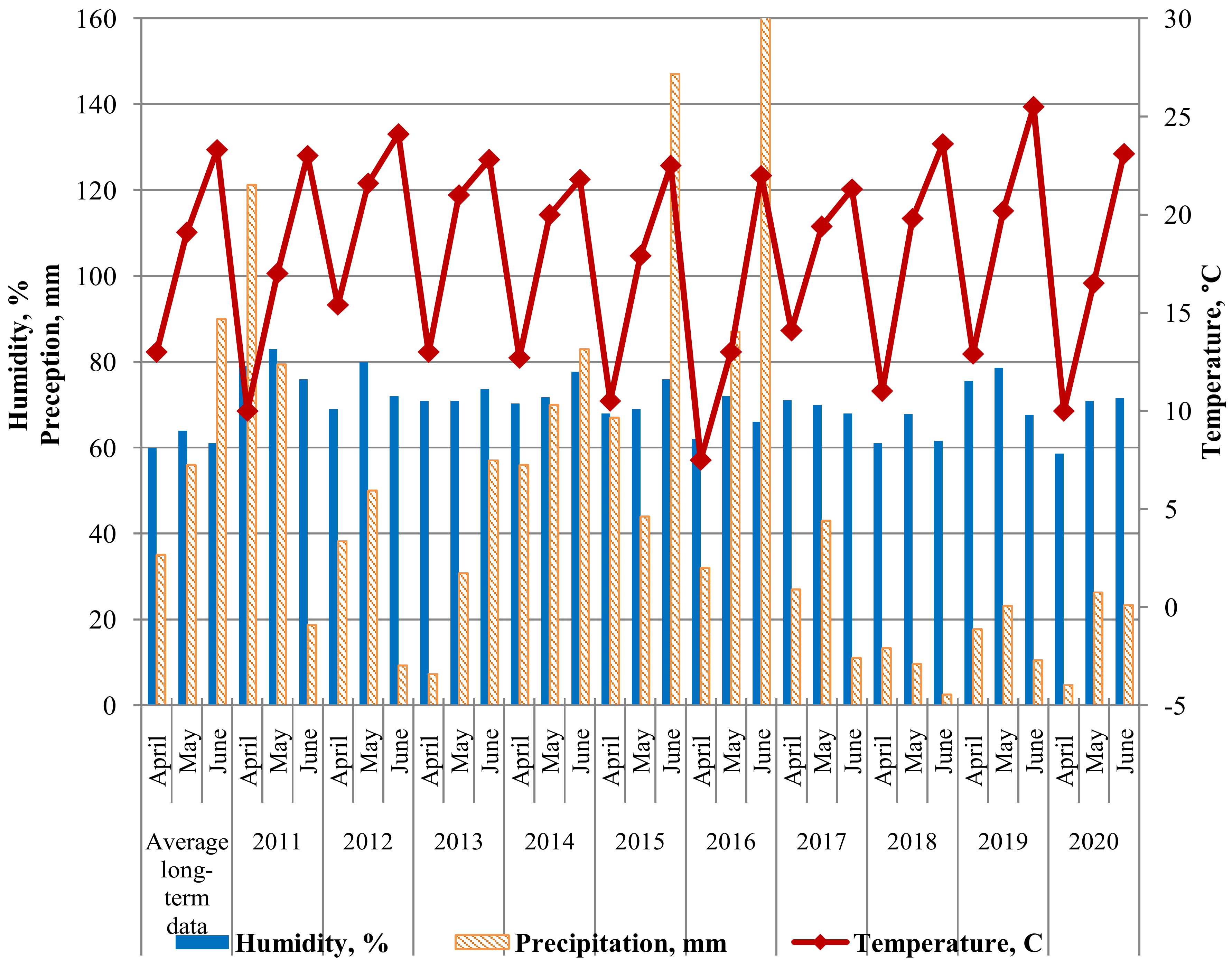
5.1. Growing Seasons 2011–2020
5.2. Obtaining Infectious Material of P. triticina
5.3. Efficiency Evaluation of the Lr Genes at the Adult Stage
5.4. Efficiency Evaluation of the Lr Genes at the Juvenile Stage
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Browse Data, By Domain—Production/Crops. FAOSTAT (Food and Agriculture Organisation of the United Nations Statistics Division). 2020. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 18 December 2021).
- Hudokormova, Z.N. Retrospective Analysis of the Development of Leaf Rust (Puccinia triticina Rob. Ex Desm. f. sp. tritici Erikss.) and Wheat Resistance and Triticale to the Pathogen. Dissertation of Candidate of Agricultural Sciences; All-Russian Research Institute of Rice: Krasnodar, Russia, 2008; 25p. [Google Scholar]
- Gultyaeva, E.I.; Aristova, M.K.; Shaidayuk, E.L.; Mironenko, N.V.; Kazartsev, I.A.; Akhmetova, A.; Kosman, E. Genetic differentiation of Puccinia triticina Erikss. in Russia. Russ. J. Genet. 2017, 53, 998–1005. [Google Scholar] [CrossRef]
- Kolomiets, T.M.; Zhemchuzhina, A.I.; Kiseleva, M.I.; Zhemchuzhina, N.S. Population and Genetic monitoring the Puccinia triticina to provide food safety of Russia. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 663, p. 012006. [Google Scholar] [CrossRef]
- Kon’kova, E.A. Population structure of Puccinia triticina Erikss. on crops of winter and spring soft wheat in the Saratov region. Plant Prot. News 2018, 4, 44–49. [Google Scholar]
- Volkova, G.V.; Vaganova, O.F.; Kudinova, O.A. Virulence of Puccinia triticina in the North Caucasus region of Russia. Span. J. Agric. Res. 2020, 18, e10SC01. [Google Scholar] [CrossRef]
- Skolotneva, E.S.; Kosman, E.; Patpour, M.; Kelbin, V.N.; Morgounov, A.I.; Shamanin, V.P.; Salina, E.A. Virulence phenotypes of Siberian wheat stem rust population in 2017–2018. Front. Agron. 2020, 2, 6. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Xia, X.C. Catalogue of Gene Symbols for Wheat: 2017 Supplement. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2017.pdf (accessed on 16 November 2021).
- And, M.D.; Linde, C. Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 2003, 40, 349–379. [Google Scholar]
- Kumar, S.; Phogat, B.S.; Vikas, V.K.; Sharma, A.K.; Saharan, M.S.; Singh, A.K.; Kumari, J.; Singh, R.; Jacob, S.R.; Singh, G.P.; et al. Mining of Indian wheat germplasm collection for adult plant resistance to leaf rust. PLoS ONE 2019, 14, e0213468. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Espino, J.; Singh, R.P.; Germán, S.; McCallum, B.D.; Park, R.F.; Chen, W.Q.; Bhardwaj, S.C.; Goyeau, H. Global status of wheat leaf rust caused by Puccinia triticina. Euphytica 2011, 179, 143–160. [Google Scholar] [CrossRef]
- Warren, R.M.; Cuddy, W.; Park, R.F.; Craigie, R.; Chng, S.F. Recent pathotype development of New Zealand cereal rust populations. N. Z. Plant Prot. 2018, 71, 314–324. [Google Scholar] [CrossRef]
- Lagudah, E.S. Molecular genetics of race non-specific rust resistance in wheat. Euphytica 2011, 179, 81–91. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; Bhavani, S.; Herrera-Foessel, S.A.; Singh, D.; Singh, P.K.; Velu, G.; Mason, R.E.; Jin, Y.; Njau, P.; et al. Race non-specific resistance to rust diseases in CIMMYT spring wheats. Euphytica 2011, 179, 175–186. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; Rajaram, S. Achieving near immunity to leaf and stripe rusts in wheat by combining slow rusting resistance genes. Acta Phytopathol. Entomol. Hung. 2000, 35, 133–139. [Google Scholar]
- Ellis, J.G.; Lagudah, E.S.; Spielmeyer, W.; Dodds, P.N. The past, present and future of breeding rust resistant wheat. Front. Plant Sci. 2014, 5, 641. [Google Scholar] [CrossRef] [Green Version]
- Duveiller, E.; Singh, R.P.; Singh, P.K.; Dababat, A.A.; Mezzalama, M. Wheat Diseases and Pests: A Guide for Field Identification; CIMMYT: Mexico City, Mexico, 2012; 138p. [Google Scholar]
- Anpilogova, L.K.; Volkova, G.V. Methods for Creating Artificial Infectious Backgrounds and Assessing Wheat Varieties for Resistance to Harmful Diseases (Head Fusarium, Rust, Powdery Mildew) Recommendations; LLC “Innovative Plant Protection Center”: Saint-Petersburg, Russia, 2000; 28p. [Google Scholar]
- Sochalova, L.P.; Khristov, Y.A. Influence of variety genotype on population structure of wheat leaf rust pathogen Puccinia recondita. Sib. Bull. Agric. Sci. 2009, 10, 61–67. [Google Scholar]
- Tyunin, V.A.; Schreider, E.R.; Gultyaeva, E.I.; Shaydayuk, E.L. Characteristics of the virulence of Puccinia triticina populations and prospects for the use of the Lr24, Lr25, and LrSp genes in spring soft wheat breeding in the Southern Urals. Vavilov J. Genet. Sel. 2017, 21, 523–529. [Google Scholar] [CrossRef]
- Zhemchuzhina, A.I.; Kiseleva, M.I.; Zhemchuzhina, N.S.; Belyakova, S.V. Virulence of populations of Puccinia triticina Erikss. in the Nonchernozem zone of Russia. Agrar. Sci. 2019, 1, 137–141. [Google Scholar] [CrossRef]
- Gultyaeva, E.I.; Sibikeev, S.N.; Druzhin, A.E.; Shaydayuk, E.L. Expansion of the genetic diversity of spring soft wheat varieties for leaf rust resistance (Puccinia triticina Eriks.) in the Lower Volga region. Agric. Biol. 2020, 55, 27–44. [Google Scholar] [CrossRef]
- Gill, H.S.; Li, C.; Sidhu, J.S.; Liu, W.; Wilson, D.; Bai, G.; Gill, B.S.; Sehgal, S.K. Fine mapping of the wheat leaf rust resistance gene Lr42. Int. J. Mol. Sci. 2019, 20, 2445. [Google Scholar] [CrossRef] [Green Version]
- Babayants, O.; Babayants, L.; Gorash, A.; Vasilev, A.; Traskovetskaya, V.; Galaev, A. Physiologic specialization of Puccinia triticina Erikss. and effectiveness of Lr-genes in the south of Ukraine during 2013–2014. Chil. J. Agric. Res. 2015, 75, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Anpilogova, L.K.; Volkova, G.V.; Vaganova, O.F.; Avdeeva, Y.V. Scientifically substantiated stages of immunological studies necessary to create rust-resistant wheat varieties. Agro XXI 2009, 10, 6. [Google Scholar]
- Ulukan, H. Wild wheats (Triticum spp.) and relatives in wheat rust diseases (Puccinia spp.) from a wheat breeder’s perspective: A general evaluation. Int. J. Agric. Biol. 2020, 23, 121–130. [Google Scholar] [CrossRef]
- Skolotneva, E.S.; Leonova, I.N.; Bukatich, E.Y.; Boiko, N.I.; Piskarev, V.V.; Salina, E.A. Effectiveness of leaf rust resistance genes against Puccinia triticina populations in Western Siberia during 2008–2017. J. Plant Dis. Prot. 2018, 125, 549–555. [Google Scholar] [CrossRef]
- Figlan, S.; Ntushelo, K.; Mwadzingeni, L.; Terefe, T.; Tsilo, T.J.; Shimelis, H. Breeding Wheat for Durable Leaf Rust Resistance in Southern Africa: Variability, Distribution, Current Control Strategies, Challenges and Future Prospects. Front. Plant Sci. 2020, 11, 549. [Google Scholar] [CrossRef]
- Khadegah Najeeb, M.A.; Thabet, M.; Negm, S.S.; El-Deeb, S.H. Monitoring of Puccinia triticina Erikss. physiologic races and effectiveness of Lr-genes in Egyptian wheat during 2014-2016 growing seasons. Int. J. Agric. Technol. 2019, 15, 35–54. [Google Scholar]
- Imbaby, I.A.; Mahmoud, M.A.; Hassan, M.E.M.; Abd-El-Aziz, A.R.M. Identification of Leaf Rust Resistance Genes in Selected Egyptian Wheat Cultivars by Molecular Markers. Sci. World J. 2014, 2014, 574285. [Google Scholar] [CrossRef]
- Nemati, Z.; Mostowfizadeh-Ghalamfarsa, R.; Dadkhodaie, A.; Mehrabi, R.; Steffenson, B.J. Virulence of Leaf Rust Physiological Races in Iran From 2010 to 2017. Plant Dis. 2020, 104, 363–372. [Google Scholar] [CrossRef]
- Park, R.F.; Bariana, H.S.; Wellings, C.R.; Wallwork, H. Detection and occurrence of a new pathotype of Puccinia triticina with virulence for Lr24 in Australia. Aust. J. Agric. Res. 2002, 53, 1069–1076. [Google Scholar] [CrossRef]
- Gessese, M.K. Description of Wheat Rusts and Their Virulence Variations Determined through Annual Pathotype Surveys and Controlled Multi-Pathotype Tests. Adv. Agric. 2019, 2019, 2673706. [Google Scholar] [CrossRef]
- Kosman, E.; Ben-Yehuda, P.; Manisterski, J.; Sela, H. Diversity of virulence phenotypes among annual populations of Puccinia triticina originating from common wheat in Israel during the period 2000–2015. Plant Pathol. 2019, 68, 1741–1748. [Google Scholar] [CrossRef]
- D’yakov, Y.T. Population Biology of Phytopathogenic Fungi; Muravey: Moscow, Russia, 1998; 382p. [Google Scholar]
- Kolmer, J.A.; Hughes, M.E. Physiologic specialization of Puccinia triticina on wheat in the United States in 2015. Plant Dis. 2017, 101, 1968–1973. [Google Scholar] [CrossRef] [Green Version]
- Roelfs, A.P.; Singh, R.P.; Saari, E.E. Rust Diseases of Wheat: Concept and Methods of Disease Management; CIMMYT: Veracruz, Mexico, 1992; 81p. [Google Scholar]
- Kolmer, J.A. Virulence of Puccinia triticina, the wheat leaf rust fungus, in the United States in 2017. Plant Dis. 2019, 103, 2113–2120. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genes Lr | Line Names, RL | Plant Infection Types by Years | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | ||
| 1 | Tc6/Centenario RL 6003 | 70MS | 40MS | 60MS | 80MS | 70MS | 80MS | 80MS | 60MS | 50MS | 60S |
| 2a | Tc6/Webster RL 6016 | 40MS | 20MS | 50MS | 50MS | 50MS | 50MS | 50MS | 50MS | 30MR | 20S |
| 2b | Tc6/Carina RL 6019 | 70S | 80S | 90S | 90S | 70S | 60S | 70S | 70S | 60S | 30S |
| 2c | Tc6/Loros RL 6025 | 60S | 80S | 90S | 100S | 90S | 80S | 80S | 80S | 70S | 55S |
| 3 | Tc6/Democrat RL 6002 | 80S | 90S | 90S | 90S | 80S | 90S | 80S | 90S | 80S | 45S |
| 3bg | Bage/Tc8 RL 6042 | 80S | 70S | 80S | 90S | 70S | 50S | 50S | 30S | 30MS | 25S |
| 3ka | Tc6/Aniversario RL 6007 | 80S | 60S | 90S | 90S | 70MS | 70MS | 80MS | 80MS | 40MS | 35S |
| 9 | Transfer/Tc6 RL 6010 | R | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 10 | Tc6/Exchange RL 6004 | 90S | 90S | 100S | 90S | 90S | 90S | 80S | 90S | 70S | 70S |
| 11 | Tc6/Hussar RL 6053 | 70S | 50S | 80S | 90S | 90S | 80S | 80S | 80S | 50S | 60S |
| 12 | Exchange/Tc6 RL 6011 | 5R | 10MR | 20MR | 30MR | 20MR | 15MR | 15MR | 10MR | 10R | 25S |
| 13 | Tc7/Frontana RL 4031 | 80S | 60S | 70S | 90S | 80S | 80S | 70MS | 80MS | 60S | 60S |
| 14a | Selkirk/Tc6 | 70S | 60S | 80S | 80S | 30MS | 40S | 50S | 50S | 30MR | 65S |
| 14b | Tc6/M.Escobar RL 6006 | 70S | 40MS | 50MS | 80MS | 70MS | 60MS | 60MS | 70MS | 50MS | 45S |
| 15 | Tc6/Kenya W1483 RL 6052 | 50MS | 30MR | 40MS | 40MS | 20MS | 20MS | 30MS | 40MS | 30MR | 25S |
| 16 | Tc6/Exchange RL 6005 | 60S | 40S | 50S | 70MS | 50MS | 60MS | 60MS | 60MS | 40MS | 65S |
| 17 | K. Lucero/Tc6 RL 6008 | 7MR | 7MR | 10MR | 15MR | 10MR | 10MR | 10MR | 10MR | 10R | 10MR |
| 18 | Tc7/Africa 43 RL 6090 | 5R | 5R | 15MR | 15MR | 10MR | 10MR | 10MR | 5MR | 5R | 15MS |
| 19 | Tc7/Tr.4A.elong. RL 6040 | 5R | R | 5R | R | R | R | R | R | 1R | 5R |
| 20 | Tc6/Timmo RL6092 | 50MR | 20MR | 30MS | 40MS | 60MS | 30MS | 30MS | 40MS | 30MR | 25S |
| 21 | Tc6/RL5406 Tetra RL6043 | 20MR | 20MR | 40S | 40MS | 20MR | 15MR | 15MR | 10MR | 10R | 10MR |
| 22a | Tc6/RL5404 Tetra RL6044 | 10R | 5R | 10R | 15MR | 15MR | 15MR | 10MR | 10MR | 10R | 10MR |
| 22b | Thatcher | 70S | 40S | 60MS | 80MS | 80MS | 70MS | 70MS | 80MS | 50MS | 45S |
| 23 | LeeFL310/Tc6 RL 6012 | 10R | 10MR | 20MR | 30MS | 50MS | 30MS | 40MS | 50MS | 50MS | 40S |
| 24 | Tc6/Agent RL 6064 | R | 0 | 5R | R | R | R | R | R | 1R | 5R |
| 25 | Tc6/Transec RL 6084 | 5R | 5R | 20MR | 20MR | 20MR | 20MR | 20MR | 30MR | 20MR | 40MS |
| 26 | Tc6/St 1.25 RL 6078 | 70S | 40MS | 60MS | 80MS | 70MS | 70MS | 70MS | 80MS | 70MS | 60S |
| 28 | Tc6/C.77.1 RL 6079 | 10R | 10R | 15MR | 10R | 10R | 10R | 10R | 10R | 10R | 40MS |
| 29 | Tc6/CS7D/Ag.11 RL 6080 | 3R | R | R | R | R | R | R | 5R | 5R | 15S |
| 30 | Tc6/Terenzio RL 6049 | 20MR | 20S | 50S | 80S | 50S | 50S | 10S | 20S | 60S | 60S |
| 32 | Tc7/Ae.squarrosa RL 6086 | 10R | 10MR | 20MR | 20MR | 20MR | 20MR | 10MR | 10MR | 10MR | 15S |
| 33 | Tc6/PI58548(1+gene) RL 6057 | 60S | 40S | 50S | 50S | 50S | 40S | 30S | 50S | 30MR | 25S |
| 34 | Tc6/PI58548 (2+gene) RL6058 | 70S | 30S | 60S | 60S | 50S | 50S | 40S | 40S | 30MR | 20S |
| 35 | Tc6/RL5711 | 5R | 5R | 7R | 10MR | 10MR | 10MR | 10MR | 5MR | 20MR | 10S |
| 36 | Neepawa5/T.speltoi 2-9 | 5R | 5R | 10MR | 10R | R | 5R | 5R | 5R | 5R | 10S |
| 37 | Tc8/VPM1 RL 6081 | 5R | 5MR | 10MR | 10MR | 10MR | 5MR | 5MR | 5MR | 20MR | 35S |
| 38 | Tc6/TMR-S74-12-24 | 1R | 5R | 20MR | 10R | 10R | 10R | 10R | 10R | 10R | 45S |
| 40 | KS89WGRC07RL50117 | 50MS | 40MS | 30MS | 40MS | 50MS | 30MS | 30MS | 40MS | 30MR | 45S |
| 39=41 | TAM107*3/T.tauschi TA2460 | R | R | 5R | 15MR | 10R | 10R | 10R | 10R | 5R | 5R |
| 42 | Century (Lr24)*3/T.tauschi TA2450 | R | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1R |
| 43 | KS92WGRC16 | - * | - | - | - | - | 0 | 0 | 0 | 0 | 0 |
| 43+24 | TAM-200 | R | 0 | 0 | R | 0 | 0 | 0 | 0 | 0 | 1R |
| 44 | Tc6/T.spelta | 40MR | 20MR | 10R | 15MR | 20MR | 20MR | 30MR | 20MR | 30MR | 20S |
| 45 | Secale cereale RL 6144 | 5R | 5R | 5R | 10R | 5R | 5R | 5R | 5R | 5R | 5S |
| 47 | Untitled | 1R | 0 | R | R | R | R | R | 5R | 5R | 0 |
| B | Tc6/Carina RL6051 | 80S | 80S | 90S | 80S | 80S | 60S | 60S | 60S | 60S | 45S |
| W(52) | Tc6/V336 | 5MR | 5MR | 10MR | 20MR | 20MR | 10MR | 10MR | 10MR | 10R | 5R |
| Exch | Tc6/Exchange | 80S | 60S | 60MS | 80S | 50S | 50S | 70S | 60S | 40MS | 45S |
| KR1KR2 | Kanred | 90S | 70S | 80S | 80MS | 70MS | 60MS | 70MS | 70MS | 70S | 65S |
| 50 | KS96WGRC 36 | - | - | - | - | - | 0 | 0 | 0 | 0 | 0 |
| 57, 58 | WL711 | - | - | - | - | - | - | - | 10MS | 10MS | 15S |
| 73 | Morocco | - | - | - | - | - | - | -- | 0 | 10MS | |
| Genes Lr | Line Names, RL | Frequency of Virulent Isolates, % (Predominant Infection Type) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | ||
| 1 | Tc6/Centenario RL 6003 | 25.7(1) | 17.6(1) | 86.0(3) | 40.0(2) | 58.0(3) | 77.6(3) | 54.6(3) | 86.8(3) | 85.5(3) | 87.8(3) |
| 2a | Tc6/Webster RL 6016 | 49.0(2) | 21.1(1) | 50.0(2) | 10.0(2) | 30.2(1) | 23.6(2) | 37.3(1) | 10.0(1) | 23.5(2) | 39.2(2) |
| 2c | Tc6/Loros RL 6025 | 70.2(3) | 47.5(2) | 86.0(3) | 65.0(3) | 56.1(3) | 74.6(3) | 62.5(3) | 40.0(2) | 79.0(3) | 52.7(3) |
| 3 | Tc6/Democrat RL 6002 | 72.3(3) | 25.7(1) | 99.0(3) | 60.0(3) | 67.7(3) | 74.9(3) | 66.8(3) | 71.7(3) | 87.4(3) | 78(3) |
| 3bg | Bage/Tc8 RL 6042 | 29.0(2) | 24.9(1) | 67.0(3) | 20.0(2) | 27.7(1) | 58.5(3) | 18.2(1) | 55.0(2) | 89.3(3) | 61(3) |
| 3ka | Tc6/Aniversario RL 6007 | 87.3(3) | 40.1(1) | 95.0(3) | 65.0(3) | 59.4(3) | 75.9(3) | 56.5(3) | 73.3(3) | 85.5(3) | 86.9(3) |
| 9 | Transfer/Tc6 RL 6010 | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 12.8(0) |
| 10 | Tc6/Exchange RL 6004 | 60.0(3) | 25.8(1) | 29.0(2) | 40.0(0) | 44.3(2) | 71.5(3) | 38.1(2) | 68.3(3) | 67.4(3) | 88.9(3) |
| 11 | Tc6/Hussar RL 6053 | 89.8(3) | 33.5(1) | 66.0(3) | 30.0(2) | 59.1(3) | 70.1(3) | 38.3(2) | 70.0(3) | 95.8(3) | 83.8(3) |
| 14a | Selkirk/Tc6 | 46.0(2) | 27.5(1) | 60.0(3) | 20.0(2) | 32.1(1) | 56.5(3) | 25.2(2) | 63.3(3) | 75.4(3) | 91.3(3) |
| 14b | Tc6/M.Escobar RL 6006 | 77.7(3) | 40.0(1) | 83.0(3) | 55.0(3) | 45.2(2) | 64.0(3) | 37.5(2) | 55.0(3) | 89.6(3) | 64.5(3) |
| 15 | Tc6/Kenya W1483 RL 6052 | 7.7(1) | 0.8(0) | 21.0(1) | 0(2) | 4.1(1) | 3.3(1) | 0(0) | 25.0(1) | 4.8(1) | 3.2(2) |
| 16 | Tc6/Exchange RL 6005 | 71.8(3) | 46.7(2) | 52.0(3) | 35.0(2) | 38.1(2) | 57.5(3) | 10.9(2) | 68.3(3) | 81.0(3) | 93.8(3) |
| 17 | K. Lucero/Tc6 RL 6008 | 50.5(3) | 42.9(2) | 64.0(3) | 20.0(2) | 11.4(1) | 27.0(2) | 14.7(2) | 75.0(3) | 96.5(3) | 89.1(3) |
| 18 | Tc7/Africa 43 RL 6090 | 6.7(0) | 28.8(1) | 66.0(3) | 20.0(2) | 1.5(0) | 8.1(1) | 11.3(0) | 58.3(3) | 69.5(3) | 80.1(3) |
| 19 | Tc7/Tr.4A.elong. RL 6040 | 0(0) | 0(0) | 0.0(0) | 0(0) | 4.9(0) | 1.3(0) | 0(0) | 1.6(0) | 0.9(0) | 9.1(0) |
| 20 | Tc6/Timmo RL6092 | 6.3(1) | 6.7(0) | 52.0(2) | 0(0) | 0(0) | 24.7(1) | 4.0(0) | 9.5(1) | 4.9(1) | 47.8(2) |
| 21 | Tc6/RL5406 Tetra RL6043 | 2.0(1) | 17.4(1) | 22.0(2) | 10.0(2) | 27.0(1) | 25.7(1) | 14.5(0) | 3.6(2) | 21.8(2) | 68.4(2) |
| 23 | LeeFL310/Tc6 RL 6012 | 62.8(2) | 38.4(2) | 43.0(2) | 30.0(2) | 26.0(1) | 68.1(3) | 35.1(2) | 80.4(3) | 66.8(3) | 69.2(3) |
| 24 | Tc6/Agent RL 6064 | 0.5(0) | 0(0) | 1.0(0) | 0(0) | 0(0) | 0(1) | 0(0) | 20.0(1) | 0(1) | 14.8(1) |
| 25 | Tc6/Transec RL 6084 | 5.8(1) | 7.7(1) | 9.0(1) | 15.0(1) | 0(1) | 22.9(1) | 10.3(1) | 60.8(3) | 35.8(2) | 81.0(3) |
| 26 | Tc6/St 1.25 RL 6078 | 58.3(3) | 43.8(2) | 60.0(3) | 30.0(2) | 22.8(2) | 55.1(3) | 27.8(1) | 73.3(3) | 72.6(3) | 84.2(3) |
| 28 | Tc6/C.77.1 RL 6079 | 2.0(1) | 36.7(1) | 55.0(3) | 45.0(2) | 36.7(1) | 39.7(2) | 23.6(1) | 55.0(3) | 24.4(1) | 83.2(3) |
| 29 | Tc6/CS7D/Ag.11 RL 6080 | 0(1) | 0.8(0) | 2.0(0) | 0(0) | 21.4(0) | 0(0) | 2.6(0) | 0(1) | 2.3(1) | 10.0(1) |
| 30 | Tc6/Terenzio RL 6049 | 78.7(3) | 63.2(3) | 69.0(3) | 60.0(3) | 1.5(2) | 75.4(3) | 45.8(2) | 73.3(3) | 40.1(2) | 91.4(3) |
| 32 | Tc7/Ae.squarrosa RL 6086 | 1.8(1) | 1.6(1) | 5.0(1) | 0(1) | 44.9(1) | 0(1) | 22.9(1) | 22.1(2) | 12.3(2) | 26.2(2) |
| 33 | Tc6/PI58548(1+gene) RL 6057 | 66.7(3) | 46.0(2) | 65.0(3) | 50.0(3) | 13.5(2) | 61.2(3) | 38.7(2) | 62.6(3) | 58.8(3) | 92(3) |
| 36 | Neepawa5/T.speltoi 2-9 | 35.5(2) | 29.4(2) | 17.0(2) | 5.0(1) | 64.2(3) | 33.9(2) | 7.6(2) | 78.7(3) | 54.8(3) | 74.8(3) |
| 38 | Tc6/TMR-S74-12-24 | 1.0(0) | 12.5(1) | 71.0(3) | 0(0) | 34.9(0) | 1.2(0) | 3.9(0) | 77.3(3) | 31.3(1) | 9.2(1) |
| 40 | KS89WGRC07RL50117 | 70.5(3) | 47.2(2) | 79.0(3) | 45.0(2) | 2.0(1) | 60.4(3) | 54.7(3) | 68.5(3) | 46.3(2) | 96.2(3) |
| 41 | TAM107*3/T.tauschi TA2460 | 0(0) | 0(0) | 0(0) | 0(0) | 48.1(1) | 8.8(0) | 2.6(0) | 2.7(0) | 0(1) | 2.2(1) |
| 42 | Century (Lr24)*3/T.tauschi TA2450 | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 7.5(1) | 1.7(1) | 2.4(1) |
| 43+24 | TAM-200 | 0(0) | 0.7(0) | 0(0) | 0(0) | 0(0) | 1.0(0) | 6.4(0) | 4.7(0) | 9.1(0) | 0(0) |
| 44 | Tc6/T.spelta | 3.2(1) | 5.6(1) | 12.0(2) | 5.0(2) | 4.7(1) | 42.0(2) | 44.8(1) | 36.7(2) | 31.4(2) | 43.7(2) |
| 45 | Secale cereale RL 6144 | 1.8(1) | 0(1) | 0(1) | 10.0(1) | 33.7(0) | 11.5(1) | 14.1(1) | 11.0(1) | 16.3(2) | 47.7(2) |
| 47 | Untitled | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 21.5(1) | 5.1(1) | 3.0(1) |
| B | Tc6/Carina RL6051 | 72.5(3) | 69.4(3) | 84.0(3) | 55.0(3) | 53.1(3) | 72.6(3) | 45.3(2) | 81.8(3) | 64.6(3) | 91.3(3) |
| W(52) | Tc6/V336 | 0.7(1) | 2.0(1) | 1.0(1) | 0(1) | 8.4(1) | 2.9(1) | 2.5(1) | 6.8(1) | 9.6(1) | 33.5(2) |
| Exch | Tc6/Exchange | 88.0(3) | 49.1(2) | 68.0(3) | 55.0(3) | 53.2(3) | 56.6(3) | 53.5(3) | 59.5(3) | 60.8(3) | 59.3(3) |
| KR1KR2 | Kanred | 96.0(3) | 89.0(3) | 93.0(3) | 60.0(3) | 62.4(3) | 69.1(3) | 56.2(3) | 17.3(2) | 62.8(3) | 85.2(3) |
| 50 | KS96WGRC 36 | - | - | - | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 13.4(1) | 41.5(2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volkova, G.; Kudinova, O.; Vaganova, O.; Agapova, V. Effectiveness of Leaf Rust Resistance Genes in the Adult and Juvenile Stages in Southern Russia in 2011–2020. Plants 2022, 11, 793. https://doi.org/10.3390/plants11060793
Volkova G, Kudinova O, Vaganova O, Agapova V. Effectiveness of Leaf Rust Resistance Genes in the Adult and Juvenile Stages in Southern Russia in 2011–2020. Plants. 2022; 11(6):793. https://doi.org/10.3390/plants11060793
Chicago/Turabian StyleVolkova, Galina, Olga Kudinova, Olga Vaganova, and Valeria Agapova. 2022. "Effectiveness of Leaf Rust Resistance Genes in the Adult and Juvenile Stages in Southern Russia in 2011–2020" Plants 11, no. 6: 793. https://doi.org/10.3390/plants11060793