
Influence of Ascophyllum nodosum Extract Foliar Spray on the Physiological and Biochemical Attributes of Okra under Drought Stress

 , ,
, ,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
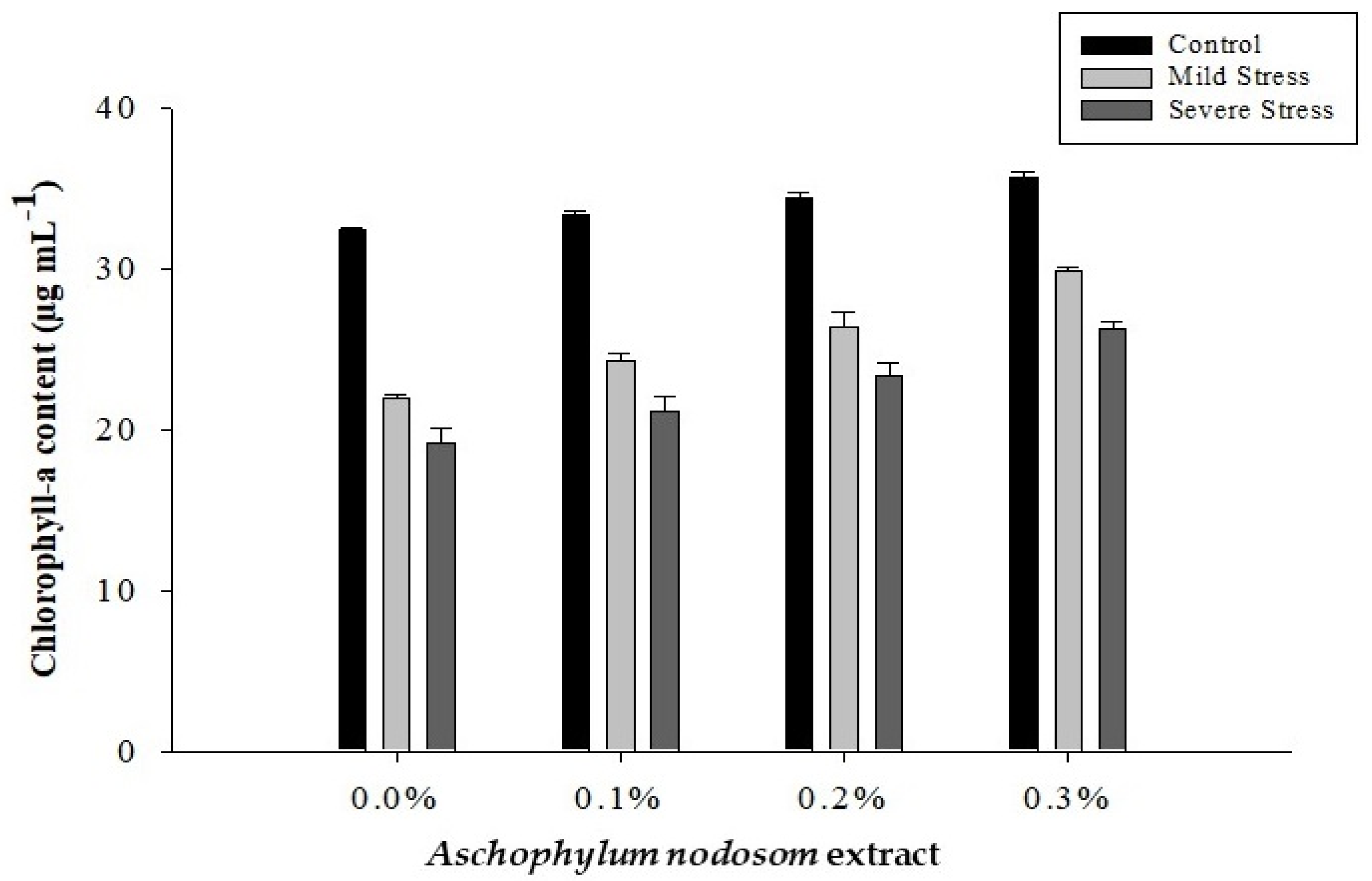
2.1. Chlorophyll-a Content (µg mL−1)
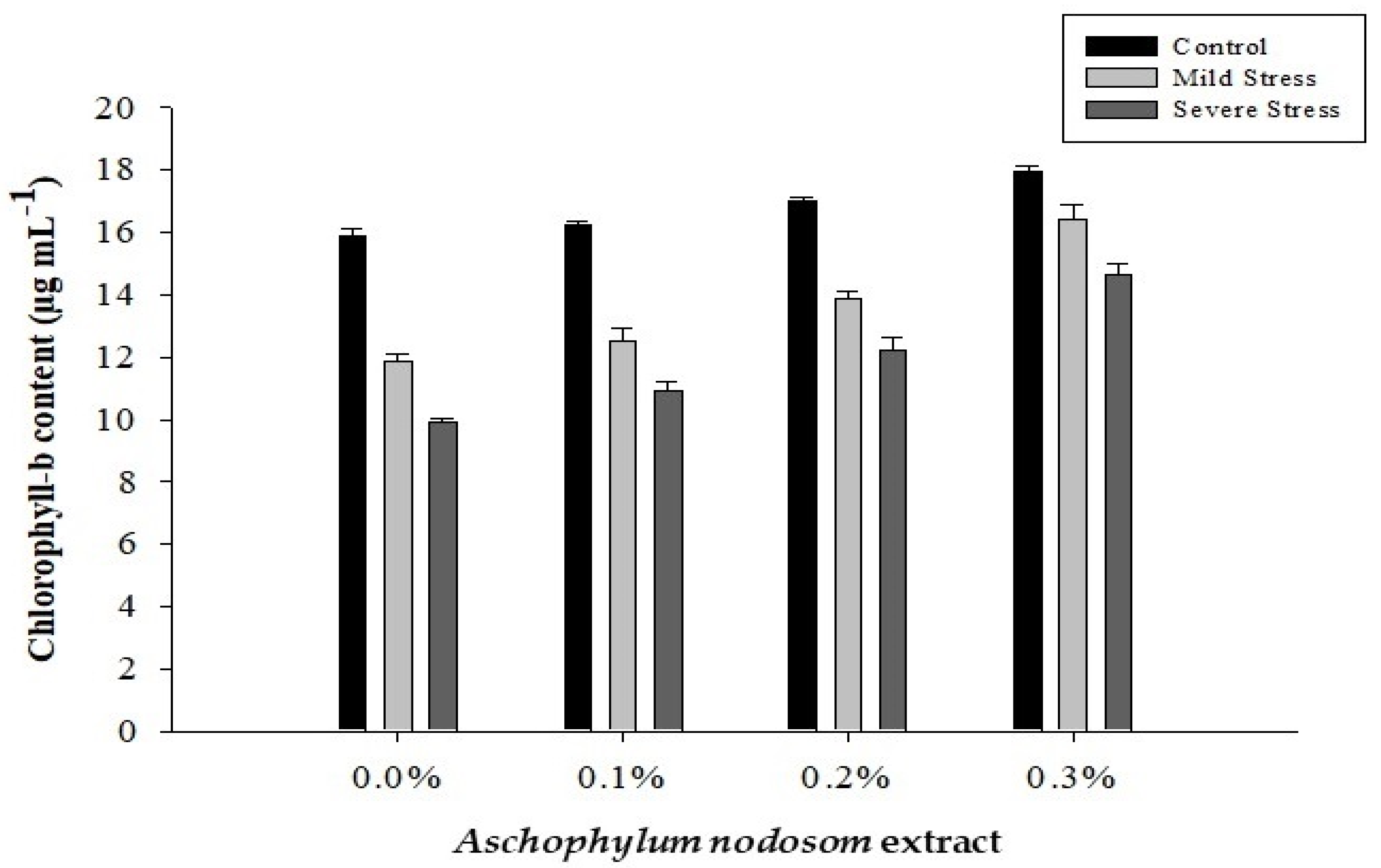
2.2. Chlorophyll-b Content (µg mL−1)
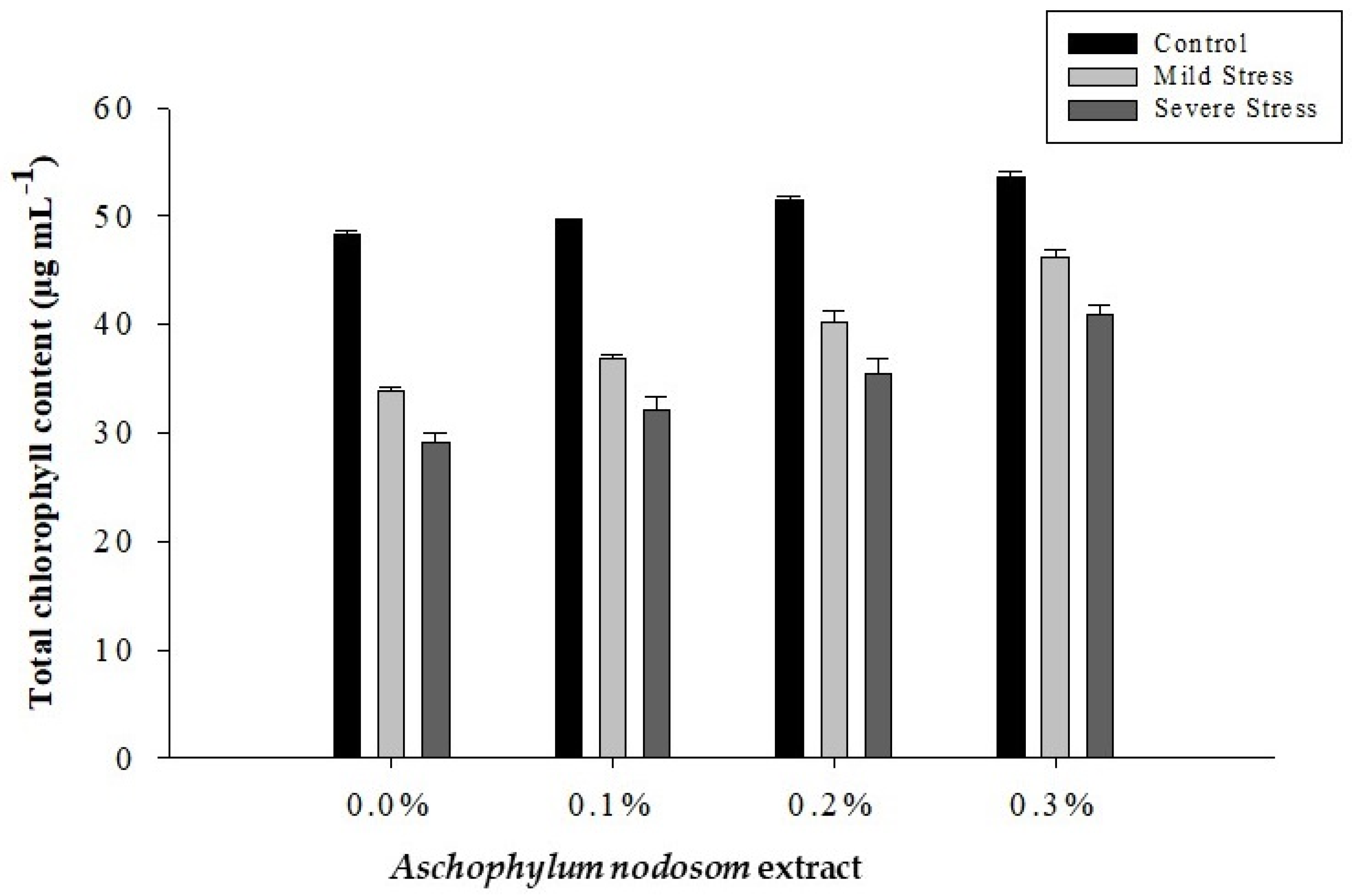
2.3. Total Chlorophyll Content (µg mL−1)
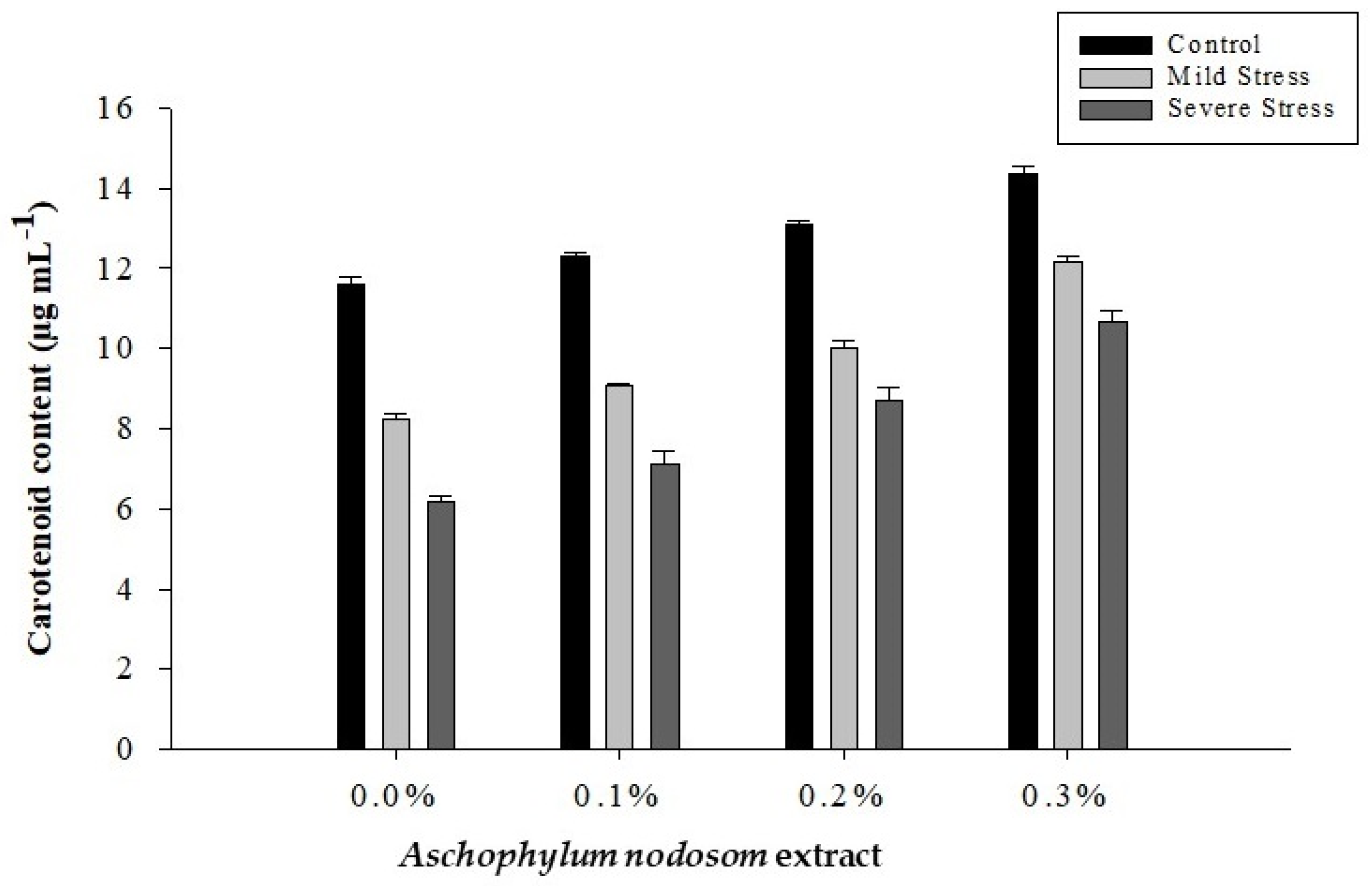
2.4. Carotenoid Content (µg mL−1)
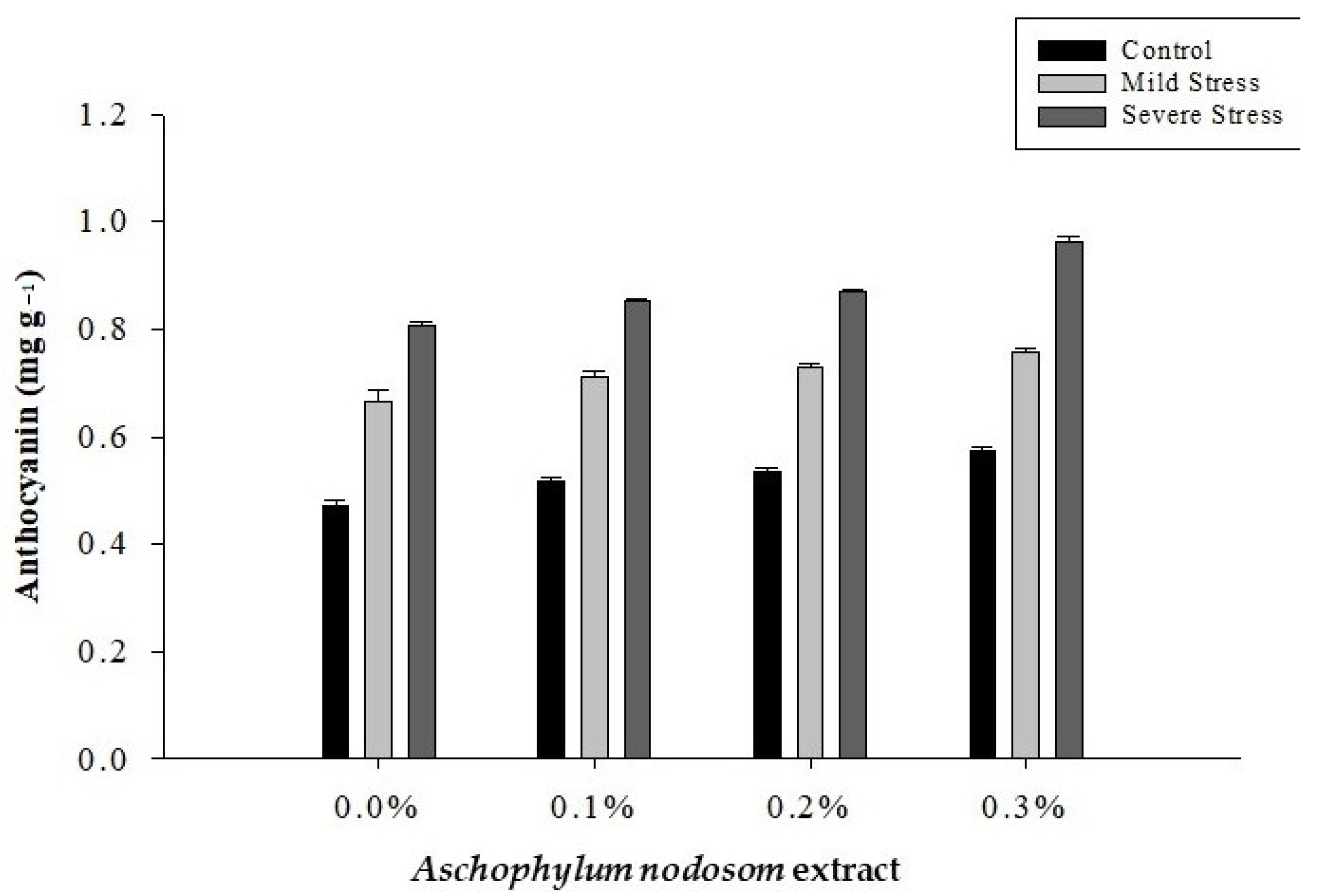
2.5. Anthocyanin Content (mg g−1)
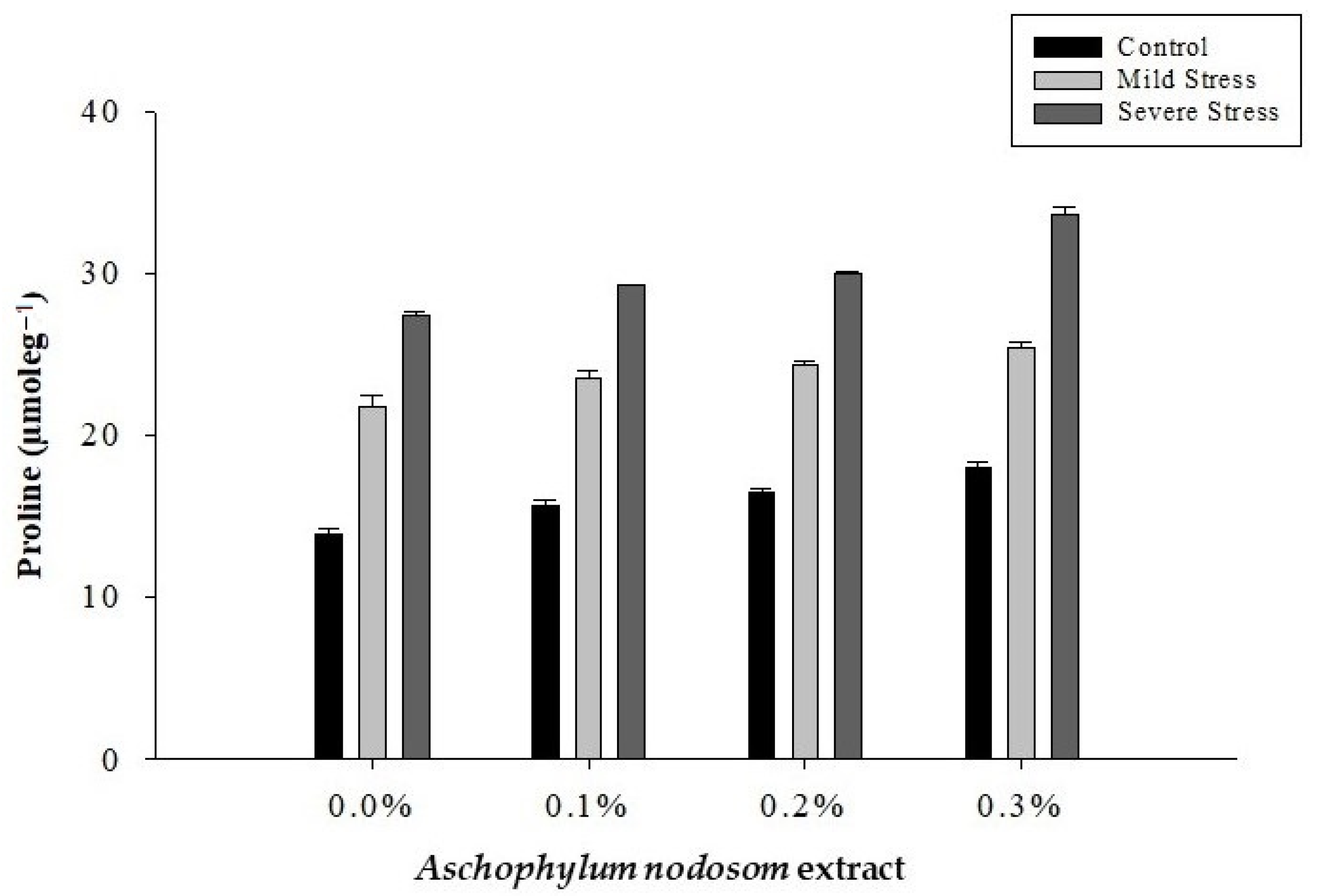
2.6. Proline Content (µmole g−1)
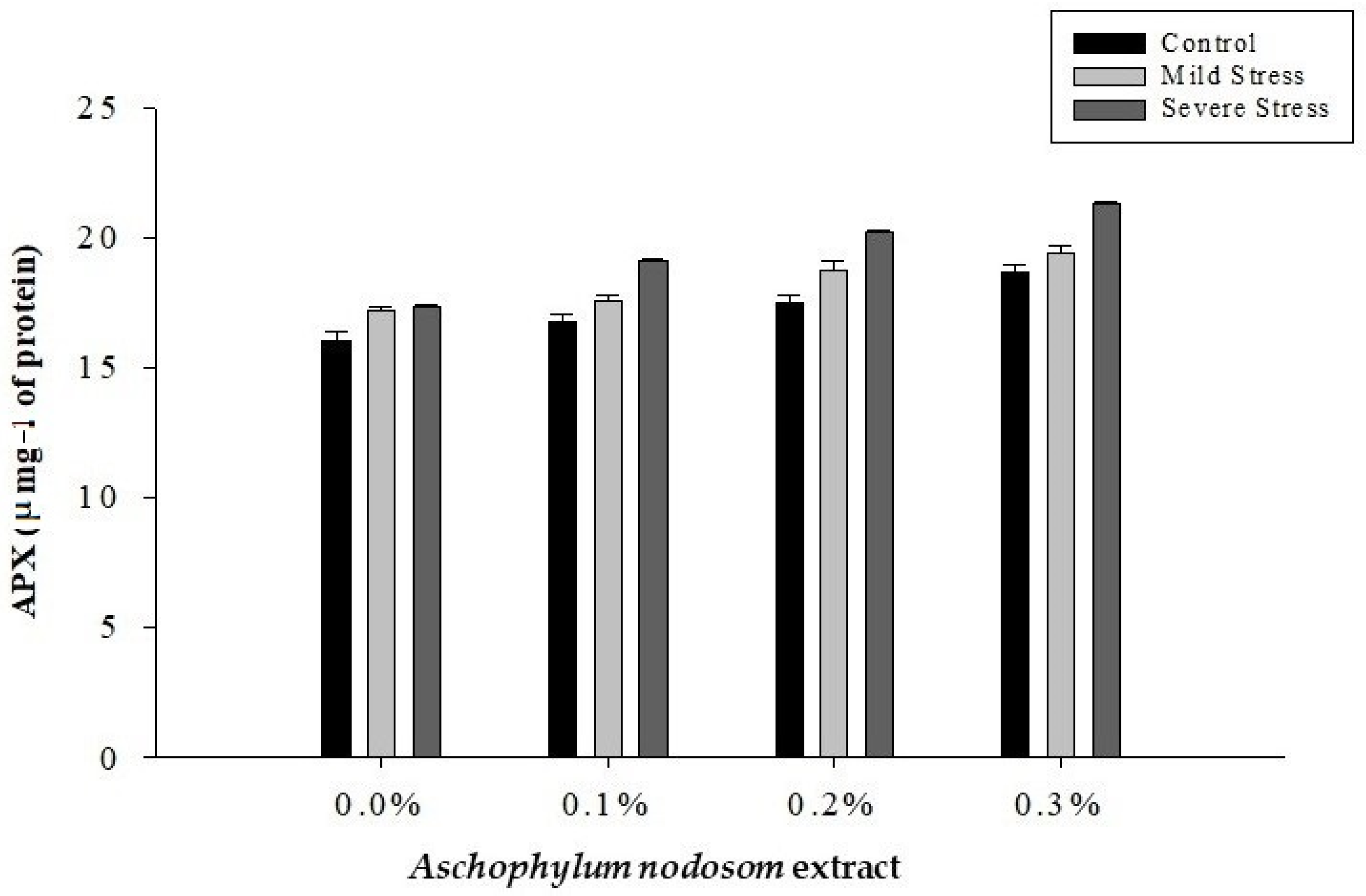
2.7. Ascorbate Peroxidase Activity (APX) (µ mg−1 Protein)
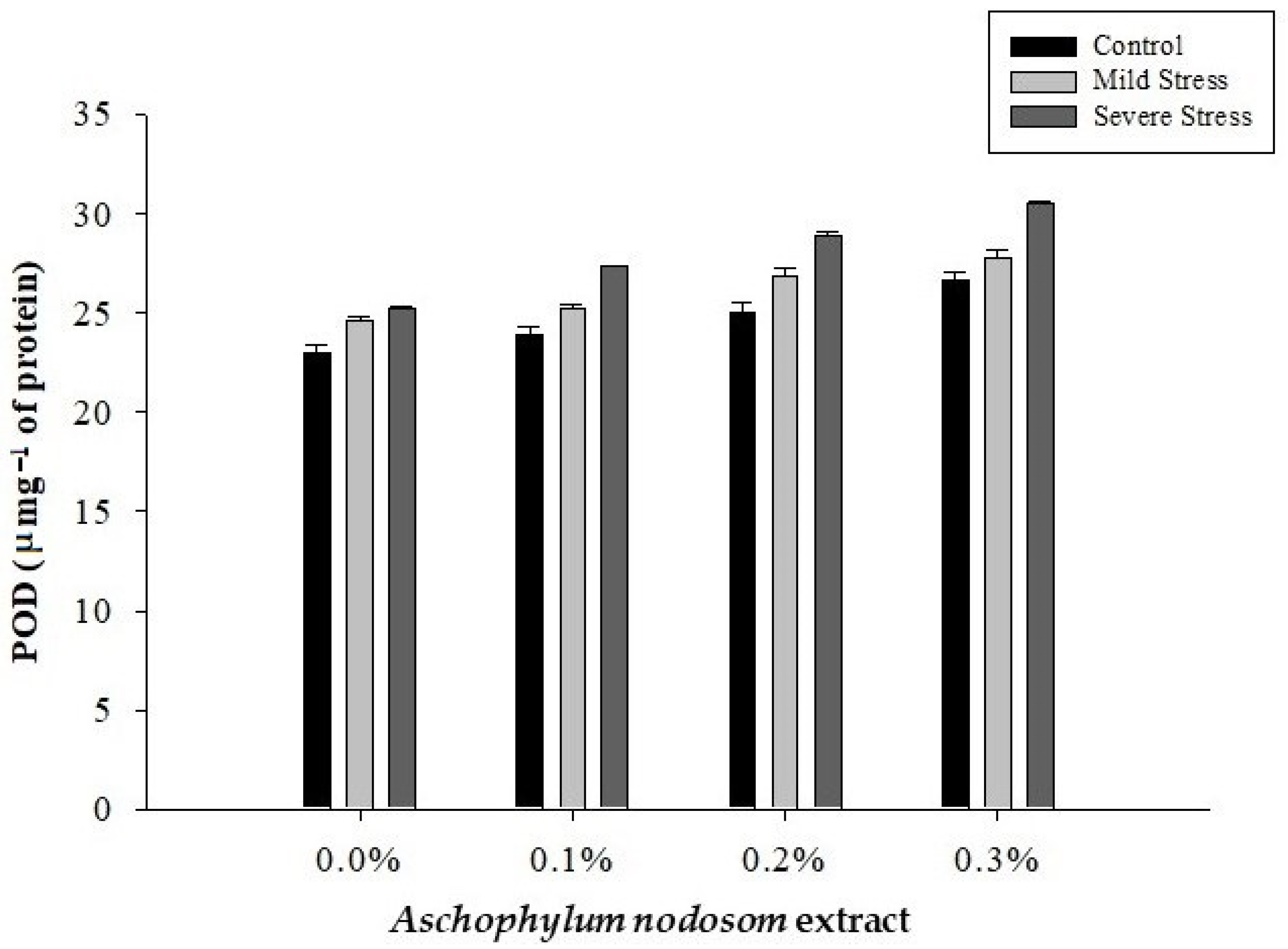
2.8. Peroxidase Activity (POD) (µ mg−1 Protein)
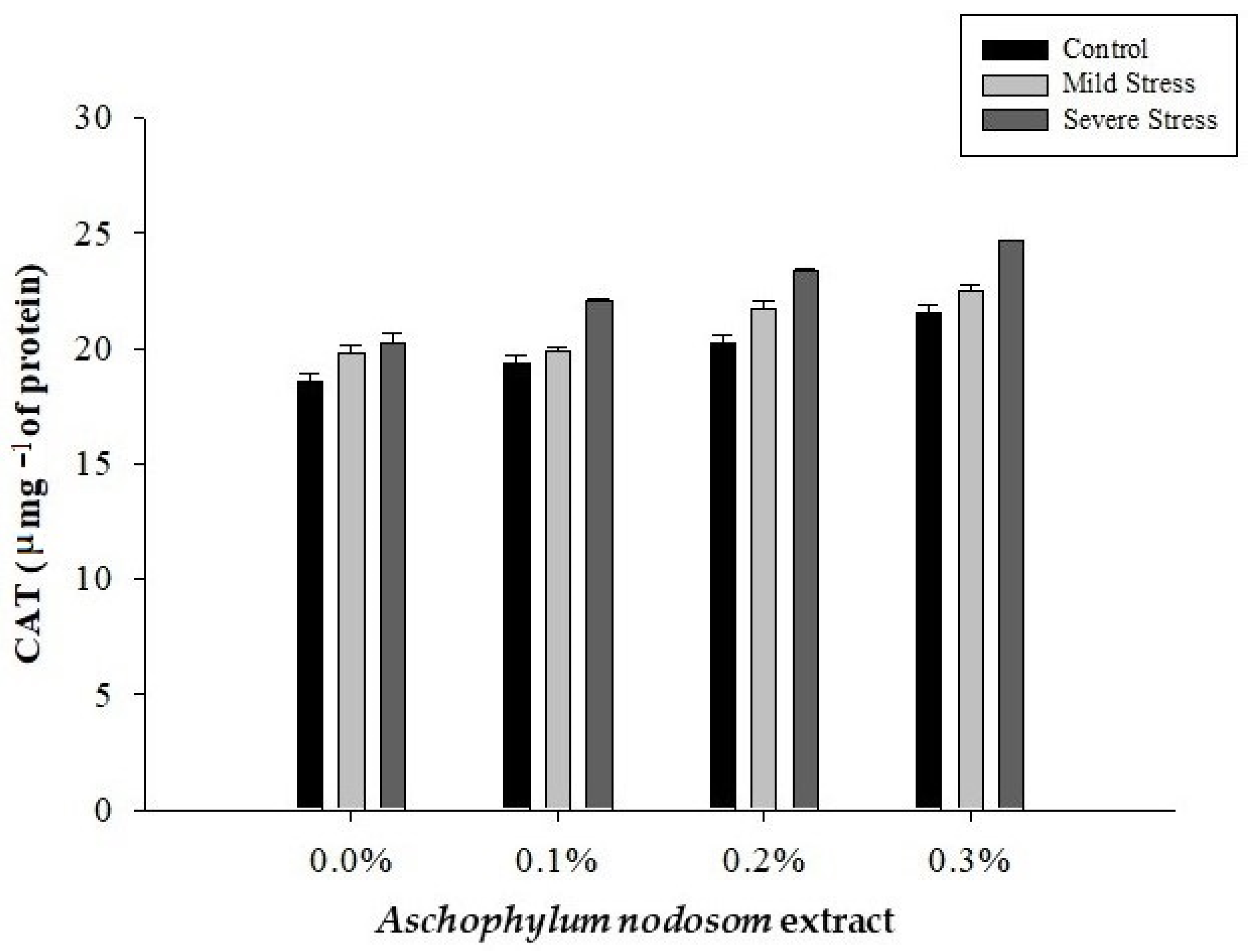
2.9. Catalase (CAT) (µ mg−1 Protein)
3. Discussion
4. Materials and Methods

4.1. Chl a, Chl b, Total Chlorophyll, and Carotenoid
4.2. Proline Determination
4.3. Antioxidant Enzymes
4.4. Anthocyanin Content
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wolf, J. Climate change adaptation as a social process. In Climate Change Adaptation in Developed Nations; Springer: Dordrecht, The Netherlands, 2001; pp. 21–32. [Google Scholar]
- Heltberg, R.; Siegel, P.B.; Jorgensen, S.L. Addressing human vulnerability to climate change: Toward a ‘no-regrets’ approach. Glob. Environ. Chang. 2009, 19, 89–99. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Kishorekumar, A.; Sankar, B.; Gopi, R.; Somasundaram, R.; Panneerselvam, R. Alterations in osmoregulation, antioxidant enzymes and indole alkaloid levels in Catharanthus roseus exposed to water deficit. Colloids Surf. B Biointerfaces 2007, 59, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; Volume 29, pp. 153–188. [Google Scholar] [CrossRef] [Green Version]
- Senthilkumar, K.; Manivasagan, P.; Venkatesan, J.; Kim, S.-K. Brown seaweed fucoidan: Biological activity and apoptosis, growth signaling mechanism in cancer. Int. J. Biol. Macromol. 2013, 60, 366–374. [Google Scholar] [CrossRef]
- Tardieu, F.; Parent, B.; Caldeira, C.F.; Welcker, C.; Oh, D.-H.; Hong, H.; Lee, S.Y.; Yun, D.-J.; Bohnert, H.J.; Dassanayake, M. Genetic and Physiological Controls of Growth under Water Deficit. Plant Physiol. 2014, 164, 1628–1635. [Google Scholar] [CrossRef] [Green Version]
- Mitra, J. Genetics and genetic improvement of drought resistance in crop plants. Curr. Sci. 2001, 80, 758–763. [Google Scholar]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop Sci. 2011, 5, 764–777. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K. Photosynthetic Response of Plants Under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Hajiboland, R.; Cheraghvareh, L.; Poschenrieder, C. Improvement of drought tolerance in Tobacco (Nicotiana rustica L.) plants by Silicon. J. Plant Nutr. 2017, 40, 1661–1676. [Google Scholar] [CrossRef]
- Lawson, T.; Vialet-Chabrand, S. Speedy stomata, photosynthesis and plant water use efficiency. New Phytol. 2019, 221, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Gagné-Bourque, F.; Bertrand, A.; Claessens, A.; Aliferis, K.A.; Jabaji, S. Alleviation of Drought Stress and Metabolic Changes in Timothy (Phleum pratense L.) Colonized with Bacillus subtilis B26. Front. Plant Sci. 2016, 7, 584. [Google Scholar] [CrossRef] [Green Version]
- Conde, A.; Chaves, M.M.; Gerós, H. Membrane Transport, Sensing and Signaling in Plant Adaptation to Environmental Stress. Plant Cell Physiol. 2011, 52, 1583–1602. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Valliyodan, B.; Nguyen, H.T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol. 2006, 9, 189–195. [Google Scholar] [CrossRef]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon-Mediated Tolerance to Salt Stress; Silicon in Agriculture; Springer: Dordrecht, The Netherlands, 2015; pp. 123–142. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Ugarte, R.A.; Sharp, G. A new approach to seaweed management in eastern Canada: The case of Ascophyllum nodosum. Cah. Biol. Mar. 2001, 42, 63–70. [Google Scholar]
- Rayirath, P.; Benkel, B.; Hodges, D.M.; Allan-Wojtas, P.; MacKinnon, S.; Critchley, A.T.; Prithiviraj, B. Lipophilic components of the brown seaweed, Ascophyllum nodosum, enhance freezing tolerance in Arabidopsis thaliana. Planta 2009, 230, 135–147. [Google Scholar] [CrossRef]
- Jithesh, M.N.; Wally, O.S.D.; Manfield, I.; Critchley, A.T.; Hiltz, D.; Prithiviraj, B. Analysis of Seaweed Extract-induced Transcriptome Leads to Identification of a Negative Regulator of Salt Tolerance in Arabidopsis. HortScience 2012, 47, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Oyelade, O.J.; Ade-Omowaye, B.I.O.; Adeomi, V.F. Influence of variety on protein, fat contents and some physical characteristics of okra seeds. J. Food Eng. 2003, 57, 111–114. [Google Scholar] [CrossRef]
- Kumar, S.; Dagnoko, S.; Haougui, A.; Ratnadass, D.; Pasternak, D.; Kouame, C. Okra (Abelmoschus spp.) in West and Central Africa: Potential and progress on its improvement. Afr. J. Agric. Res. 2010, 5, 3590–3598. [Google Scholar]
- Schalau, J. Backyard Gardener. 2002. Available online: http://ag.arizona.edu./yavapai/anr/hort/byg/ (accessed on 30 August 2021).
- Benchasri, S. Okra (Abelmoschus esculentus L.). Moench as a valuable vegetable of the world. Ratar. Povrt. 2012, 49, 105–112. [Google Scholar]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Saeed, A.M.; Abid, A.J.; Malik, A.; Kumbhar, M.B. Response of okra to water stress. Sarhad J. Agric. 2003, 19, 73–79. [Google Scholar]
- Grimes, D.W.; Miller, R.J.; Dickens, L. Water stress during flowering of cotton. Calif. Agric. 1970, 24, 4. [Google Scholar]
- Mbagwu, J.S.C.; Adesipe, F.A. Response of three okra (Abelmoschus esculentus L. Moench) cultivars to irrigation at specific growth stages. Sci. Hortic. 1987, 31, 35–43. [Google Scholar] [CrossRef]
- Naveed, A.; Khan, A.A.; Rauf, S. The potential of breeding okra (Abelmoschus esculentus L.) for water stress tolerance. In Crop Production for Agricultural Improvement; Ashraf, M., Ozturk, M., Ahmad, M.S.A., Aksoy, A., Eds.; Springer Science and Business Media B.V.: Berlin/Heidelberg, Germany, 2012; pp. 217–235. [Google Scholar]
- Siemonsma, J.S. West African okra – Morphological and cytogenetical indications for the existence of a natural amphidiploid of Abelmoschus esculentus (L.) Moench and A. Manihot (L.) Medikus. Euphytica 1982, 31, 241–252. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Etienne, P.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Brassica napus Growth is Promoted by Ascophyllum nodosum (L.) Le Jol. Seaweed Extract: Microarray Analysis and Physiological Characterization of N, C, and S Metabolisms. J. Plant Growth Regul. 2013, 32, 31–52. [Google Scholar] [CrossRef]
- Kałużewicz, A.; Krzesiński, W.; Spiżewski, T.; Zaworska, A. Effect of Biostimulants on Several Physiological Characteristics and Chlorophyll Content in Broccoli under Drought Stress and Re-watering. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Wally, O.S.; Critchley, A.T.; Hiltz, D.; Craigie, J.S.; Han, X.; Zaharia, L.I.; Prithiviraj, B. Regulation of phytohormone biosynthesis and accumulation in Arabidopsis following treatment with commercial extract from the marine macroalga Ascophyllum nodosum. J. Plant Growth Regul. 2013, 32, 324–339. [Google Scholar] [CrossRef]
- Zavaleta-Mancera, H.A.; López-Delgado, H.; Loza-Tavera, H.; Mora-Herrera, M.; Trevilla-García, C.; Vargas-Suárez, M.; Ougham, H. Cytokinin promotes catalase and ascorbate peroxidase activities and preserves the chloroplast integrity during dark-senescence. J. Plant Physiol. 2007, 164, 1572–1582. [Google Scholar] [CrossRef]
- Thirumaran, G.; Arumugam, M.; Arumugam, R.; Anantharaman, P. Effect of seaweed liquid fertilizer on growth and pigment concentration of Cyamopsis tetrogonolaba (L) Taub. Am.-Eurasian J. Agron. 2009, 2, 50–56. [Google Scholar]
- Shehata, S.M.; Schmidhalter, U.; Valšíková, M.; Junge, H. Effect of Bio-stimulants on Yield and Quality of Head Lettuce Grown Under Two Sources of Nitrogen. Gesunde Pflanz. 2016, 68, 33–39. [Google Scholar] [CrossRef]
- Longstreth, D.J.; Bolaños, J.A.; Smith, J.E. Salinity Effects on Photosynthesis and Growth in Alternanthera philoxeroides (Mart.) Griseb. Plant Physiol. 1984, 75, 1044–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A. Activity and Mechanism of Ascophyllum nodosum Extract Induced Salinity Tolerance in Tomato. Master’s Thesis, Dalhousie University Halifax, Halifax, NS, Canada, 2014. [Google Scholar]
- Bhusal, N.; Bhusal, S.J.; Yoon, T.-M. Comparisons of physiological and anatomical characteristics between two cultivars in bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- Vemmos, S.N.; Petri, E.; Stournaras, V. Seasonal changes in photosynthetic activity and carbohydrate content in leaves and fruit of three fig cultivars (Ficus carica L.). Sci. Hortic. 2013, 160, 198–207. [Google Scholar] [CrossRef]
- Allen, V.G.; Pond, K.R.; Saker, K.E.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Schmidt, R.E.; Fike, J.H.; Zhang, X.; et al. Tasco: Influence of a brown seaweed on antioxidants in forages and livestock—A review. J. Anim. Sci. 2001, 79, E21–E31. [Google Scholar] [CrossRef]
- Basavaraja, P.K.; Yogendra, N.D.; Zodape, S.T.; Prakash, R.; Ghosh, A. Effect of seaweed sap as foliar spray on growth and yield of hybrid maize. J. Plant Nutr. 2018, 41, 1851–1861. [Google Scholar] [CrossRef]
- El-Kaoaua, M.; Chernane, H.; Benaliat, A.; Neamallah, L. Seaweed liquid extracts effect on Salvia officinalis growth, biochemical compounds and water deficit tolerance. Afr. J. Biotechnol. 2013, 12, 4481–4489. [Google Scholar]
- El-Tayeb, M.A.; El-Enany, A.E.; Ahmed, N.L. Salicylic acid-induced adaptive response to copper stress in sunflower (Helianthus annuus L.). Plant Growth Regul. 2006, 50, 191–199. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Y.; Yang, H.; Li, H.; Xu, W.; Chen, G.; Zhu, H. Physicochemical Characterization, Antioxidant and Immunostimulatory Activities of Sulfated Polysaccharides Extracted from Ascophyllum nodosum. Molecules 2018, 23, 1912. [Google Scholar] [CrossRef] [Green Version]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Effect on yield, total phenolic, total flavonoid and total isothiocyanate content of two broccoli cultivars (Brassica oleraceae var italica) following the application of a commercial brown seaweed extract (Ascophyllum nodosum). Agric. Food Sci. 2014, 23, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Rahbarian, R.; Khavari-Nejad, R.; Ganjeali, A.; Bagheri, A.; Najafi, F. Drought Stress Effects on Photosynthesis, Chlorophyll Fluorescence and Water Relations in Tolerant and Susceptible Chickpea (Cicer Arietinum L.) Genotypes. Acta Biol. Crac. Bot. 2011, 53, 47–56. [Google Scholar] [CrossRef]
- Guo, J.Y.; White, E. Autophagy, metabolism, and cancer. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Cold Springer Harbor, NY, USA, 2016; Volume 81, pp. 73–78. [Google Scholar]
- Merzlyak, M.N.; Chivkunova, O.B.; Solovchenko, A.E.; Naqvi, K.R. Light absorption by anthocyanin in juvenile, stressed, and senescing leaves. J. Exp. Bot. 2008, 59, 3903–3911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, N.M.; Smith, W.K. Attenuation of incident light in Galax urceolata (Diapensiaceae): Concerted influence of adaxial and abaxial anthocyanic layers on photoprotection. Am. J. Bot. 2007, 94, 784–790. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A novel type of seaweed extract as a natural alternative to the use of iron chelates in strawberry production. Sci. Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Alam, M.Z.; Braun, G.; Norrie, J.; Hodges, D.M. Effect of Ascophylum Nodosum extract application on plant growth, fruit yield and soil microbial communities of strawberry. Can. J. Plant Sci. 2013, 93, 23–36. [Google Scholar] [CrossRef]
- Roussos, P.; Denaxa, N.-K.; Damvakaris, T. Strawberry fruit quality attributes after application of plant growth stimulating compounds. Sci. Hortic. 2009, 119, 138–146. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52. [Google Scholar] [CrossRef] [Green Version]
- Ahammed, G.J.; Li, X.; Wan, H.; Zhou, G.; Cheng, Y. SlWRKY81 reduces drought tolerance by attenuating proline biosynthesis in tomato. Sci. Hortic. 2020, 270, 109444. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Yazici, I.; Türkan, I.; Sekmen, A.H.; Demiral, T. Salinity tolerance of purslane (Portulaca oleracea L.) is achieved by enhanced antioxidative system, lower level of lipid peroxidation and proline accumulation. Environ. Exp. Bot. 2007, 61, 49–57. [Google Scholar] [CrossRef]
- Sangha, J.S.; Kelloway, S.; Critchley, A.T.; Prithiviraj, B. Seaweeds (macroalgae) and their extracts as contributors of plant productivity and quality: The current status of our understanding. Adv. Bot. Res. 2014, 71, 189–219. [Google Scholar]
- Renuka, R.; Rathinavel, S. Alleviating potential of seaweed liquid extract of Enteromorpha prolifera on salt induced changes in cluster bean plants. Plant Arch. 2006, 6, 845–848. [Google Scholar]
- Carvalho, M.E.A.; e Castro, P.R.D.C. Seaweeds as Plant Biostimulants. In Seaweeds as Plant Fertilizer, Agricultural Biostimulants and Animal Fodder; CRC Press: Boca Raton, FL, USA, 2019; pp. 80–99. [Google Scholar]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed Extracts Enhance Salam Turfgrass Performance during Prolonged Irrigation Intervals and Saline Shock. Front. Plant Sci. 2017, 8, 830. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Luan, S. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 2012, 35, 53–60. [Google Scholar] [CrossRef]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Santaniello, A.; Scartazza, A.; Gresta, F.; Loreti, E.; Biasone, A.; Di Tommaso, D.; Piaggesi, A.; Perata, P. Ascophyllum nodosum Seaweed Extract Alleviates Drought Stress in Arabidopsis by Affecting Photosynthetic Performance and Related Gene Expression. Front. Plant Sci. 2017, 8, 1362. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Dubé, M.A.; McLean, D.D.; Kates, M. Biodiesel production from waste cooking oil: 2. Economic assessment and sensitivity analysis. Bioresour. Technol. 2003, 90, 229–240. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-Based Biostimulants: Sustainable Applications in Agriculture for the Stimulation of Plant Growth, Stress Tolerance, and Disease Management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [Green Version]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Dutot, M.; Fagon, R.; Hemon, M.; Rat, P. Antioxidant, Anti-inflammatory, and Anti-senescence Activities of a Phlorotannin-Rich Natural Extract from Brown Seaweed Ascophyllum nodosum. Appl. Biochem. Biotechnol. 2012, 167, 2234–2240. [Google Scholar] [CrossRef]
- Gill, P.K.; Sharma, A.D.; Singh, P.; Bhullar, S.S. Osmotic stress-induced changes in germination, growth and soluble sugar content of Sorghum bicolor (L.) Moench seeds. Bulg. J. Plant Physiol. 2002, 28, 12–25. [Google Scholar]
- Fike, J.H.; Allen, V.G.; Schmidt, R.E.; Zhang, X.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Coelho, R.W.; Wester, D.B. Tasco-Forage: I. Influence of a seaweed extract on antioxidant activity in tall fescue and in ruminants. J. Anim. Sci. 2001, 79, 1011–1021. [Google Scholar] [CrossRef]
- Papenfus, H.B.; Kulkarni, M.G.; Stirk, W.A.; Finnie, J.F.; Van Staden, J. Effect of a commercial seaweed extract (Kelpak®) and polyamines on nutrient-deprived (N, P and K) okra seedlings. Sci. Hortic. 2013, 151, 142–146. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Hussain, I.; Rasheed, R.; Iqbal, M.; Riaz, M.; Arif, M.S. Advances in microbe-assisted reclamation of heavy metal contaminated soils over the last decade: A review. J. Environ. Manag. 2017, 198, 132–143. [Google Scholar] [CrossRef]
- Lebrun, M.; De Zio, E.; Miard, F.; Scippa, G.S.; Renzone, G.; Scaloni, A.; Trupiano, D. Amending an As/Pb contaminated soil with biochar, compost and iron grit: Effect on Salix viminalis growth, root proteome profiles and metal (loid) accumulation indexes. Chemosphere 2020, 244, 125397. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.-C.; Zhang, H.-B.; Wang, R.; Wang, G.-X.; Shao, Y.; Li, C.-X.; Ma, S.-T. Effects of mine wastewater irrigation on activities of soil enzymes and physiological properties, heavy metal uptake and grain yield in winter wheat. Ecotoxicol. Environ. Saf. 2015, 113, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Fu, D.; Yang, F. Effect of Saponin on the Phytoextraction of Pb, Cd and Zn from Soil Using Italian Ryegrass. Bull. Environ. Contam. Toxicol. 2015, 94, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Guo, D.; Mahar, A.; Ma, F.; Li, R.; Shen, F.; Wang, P.; Zhang, Z. Streptomyces pactum assisted phytoremediation in Zn/Pb smelter contaminated soil of Feng County and its impact on enzymatic activities. Sci. Rep. 2017, 7, 46087. [Google Scholar] [CrossRef] [Green Version]
- Kabata-Pendias, A. Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Agnello, A.C.; Potysz, A.; Fourdrin, C.; Huguenot, D.; Chauhan, P.S. Impact of pyrometallurgical slags on sunflower growth, metal accumulation and rhizosphere microbial communities. Chemosphere 2018, 208, 626–639. [Google Scholar] [CrossRef]
- Lal, S.; Ratna, S.; Ben Said, O.; Kumar, R. Biosurfactant and exopolysaccharide-assisted rhizobacterial technique for the remediation of heavy metal contaminated soil: An advancement in metal phytoremediation technology. Environ. Technol. Innov. 2018, 10, 243–263. [Google Scholar] [CrossRef]
- Prapagdee, B.; Chanprasert, M.; Mongkolsuk, S. Bioaugmentation with cadmium-resistant plant growth-promoting rhizobacteria to assist cadmium phytoextraction by Helianthus annuus. Chemosphere 2013, 92, 659–666. [Google Scholar] [CrossRef]
- Yang, C.M.; Chang, K.W.; Yin, M.H.; Huang, H.M. Methods for the determination of the chlorophylls and their derivatives. Taiwania 1998, 43, 116–122. [Google Scholar]
- Holm, G. Chlorophyll mutations in barley. Acta Agric. Scand. 1954, 4, 457–471. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta BBA Bioenergy 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ahmad, H.; Hayat, S.; Ali, M.; Liu, T.; Cheng, Z. The combination of arbuscular mycorrhizal fungi inoculation (Glomus versiforme) and 28-homobrassinolide spraying intervals improves growth by enhancing photosynthesis, nutrient absorption, and antioxidant system in cucumber (Cucumis sativus L.) under salinity. Ecol. Evol. 2018, 8, 5724–5740. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and scavenging of active oxygen in photosynthesis. Photoinhibition 1987, 227–287. [Google Scholar]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Zhang, D.; Quantick, P.C. Effects of chitosan coating on enzymatic browning and decay during postharvest storage of litchi (Litchi chinensis Sonn.) fruit. Postharvest Biol. Technol. 1997, 12, 195–202. [Google Scholar] [CrossRef]
- Steel, R.G.; Torrie, J.H. Principles and Procedures of Statistics; McGraw-Hill Book Co., Inc.: New York, NY, USA, 1980; p. 481. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Chlorophyll-a (µg mL−1) | Chlorophyll-b (µg mL−1) | Total Chlorophyll (µg mL−1) | Carotenoid (µg mL−1) | Anthocyanin (mg g−1) |
|---|---|---|---|---|---|
| ANE Levels (%) | |||||
| 0 | 24.56 d | 12.55 d | 37.11 d | 8.67 d | 0.6479 d |
| 1 | 26.32 c | 13.23 c | 39.55 c | 9.50 c | 0.6929 c |
| 2 | 28.05 b | 14.35 b | 42.40 b | 10.60 b | 0.7121 b |
| 3 | 30.61 a | 16.35 a | 46.96 a | 12.40 a | 0.7635 a |
| Tukey HSD | 1.3196 * | 0.6821 * | 1.7369 * | 0.4310 * | 0.0216 * |
| Drought Stress (DS) | |||||
| Control | 34.01 a | 16.77 a | 50.78 a | 12.85 a | 0.5235 c |
| Mild stress | 25.64 b | 13.67 b | 39.31 b | 9.85 b | 0.7160 b |
| Severe stress | 22.50 c | 11.92 c | 34.32 c | 8.17 c | 0.8728 a |
| Tukey HSD | 1.0346 * | 0.5348 * | 1.3617 * | 0.3379 * | 0.0169 * |
| ANE × DS | 2.9863 * | 1.5435 * | 3.9304 * | 0.9752 * | 0.0488 * |
| Treatments | Proline (µmole g−1) | APX (µ mg−1 Protein) | POD (µ mg−1 Protein) | CAT (µ mg−1 Protein) |
|---|---|---|---|---|
| ANE Levels (%) | ||||
| 0 | 20.99 d | 16.87 d | 24.29 d | 19.52 d |
| 1 | 22.81 c | 17.81 c | 25.51 c | 20.45 c |
| 2 | 23.58 b | 18.83 b | 26.95 b | 21.77 b |
| 3 | 25.65 a | 19.79 a | 28.34 a | 22.89 a |
| Tukey HSD | 0.8686 * | 0.4857 * | 0.6849 * | 0.6565 * |
| Drought Stress(DS) | ||||
| Control | 15.98 c | 17.236 c | 24.67 c | 19.93 c |
| Mild stress | 23.74 b | 18.25 b | 26.15 b | 20.96 b |
| Severe stress | 30.06 a | 19.94 a | 28.01 a | 22.58 a |
| Tukey HSD | 0.6810 * | 0.3808 * | 0.5370 * | 0.5147 * |
| ANE × DS | 1.9656 * | 1.0990 * | 1.5499 * | 1.4857 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, J.; Jan, I.; Ullah, H.; Ahmed, N.; Alam, M.; Ullah, R.; El-Sharnouby, M.; Kesba, H.; Shukry, M.; Sayed, S.; et al. Influence of Ascophyllum nodosum Extract Foliar Spray on the Physiological and Biochemical Attributes of Okra under Drought Stress. Plants 2022, 11, 790. https://doi.org/10.3390/plants11060790
Ali J, Jan I, Ullah H, Ahmed N, Alam M, Ullah R, El-Sharnouby M, Kesba H, Shukry M, Sayed S, et al. Influence of Ascophyllum nodosum Extract Foliar Spray on the Physiological and Biochemical Attributes of Okra under Drought Stress. Plants. 2022; 11(6):790. https://doi.org/10.3390/plants11060790
Chicago/Turabian StyleAli, Jawad, Ibadullah Jan, Hidayat Ullah, Nazeer Ahmed, Mukhtar Alam, Rafi Ullah, Mohamed El-Sharnouby, Hosny Kesba, Mustafa Shukry, Samy Sayed, and et al. 2022. "Influence of Ascophyllum nodosum Extract Foliar Spray on the Physiological and Biochemical Attributes of Okra under Drought Stress" Plants 11, no. 6: 790. https://doi.org/10.3390/plants11060790