
The Establishment and Spread of a Newly Introduced Begomovirus in a Dry Tropical Environment Using Tomato Yellow Leaf Curl Virus as a Case Study


Abstract
:1. Introduction
2. Results
2.1. TYLCV Genetic Diversity
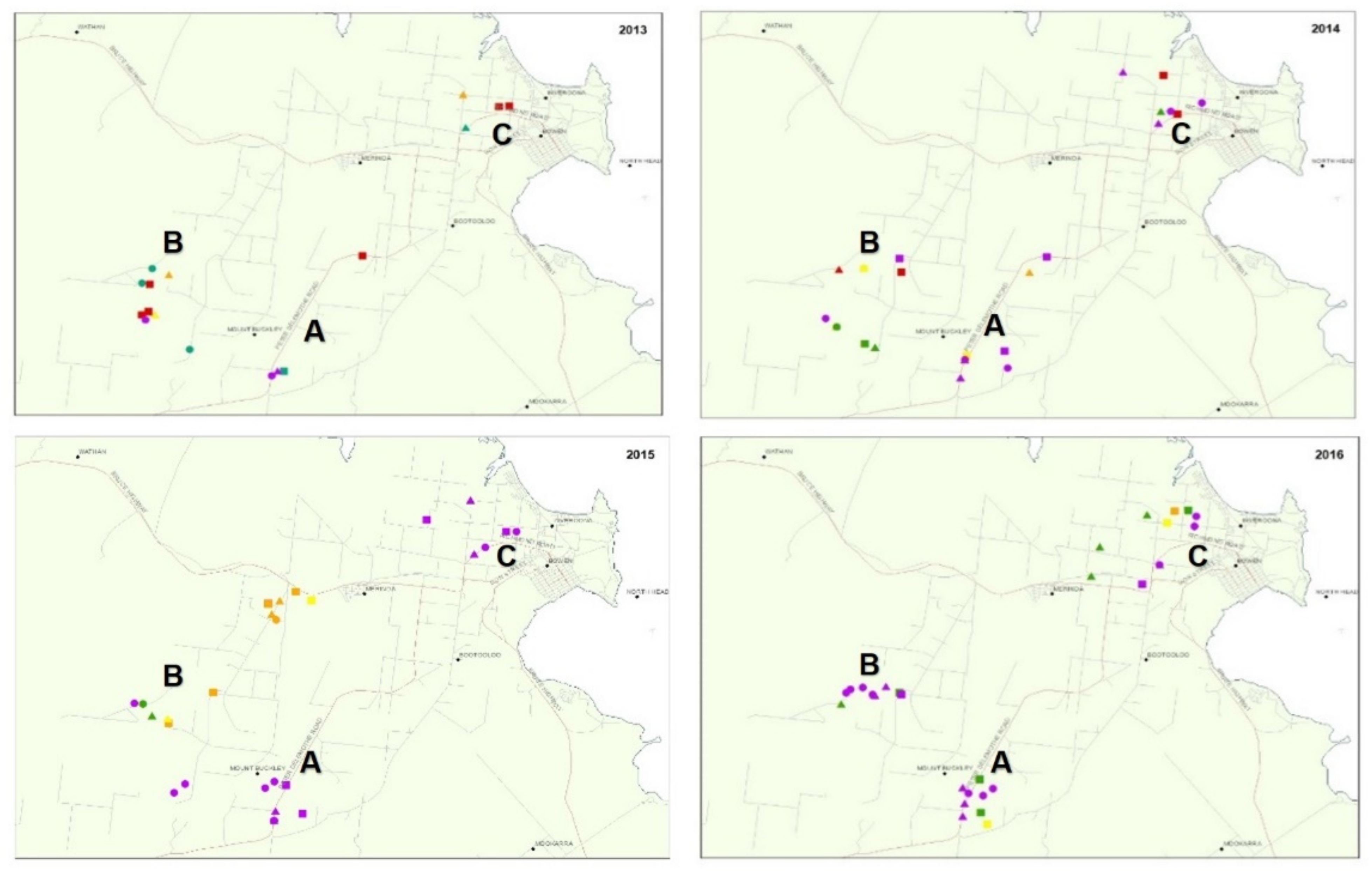
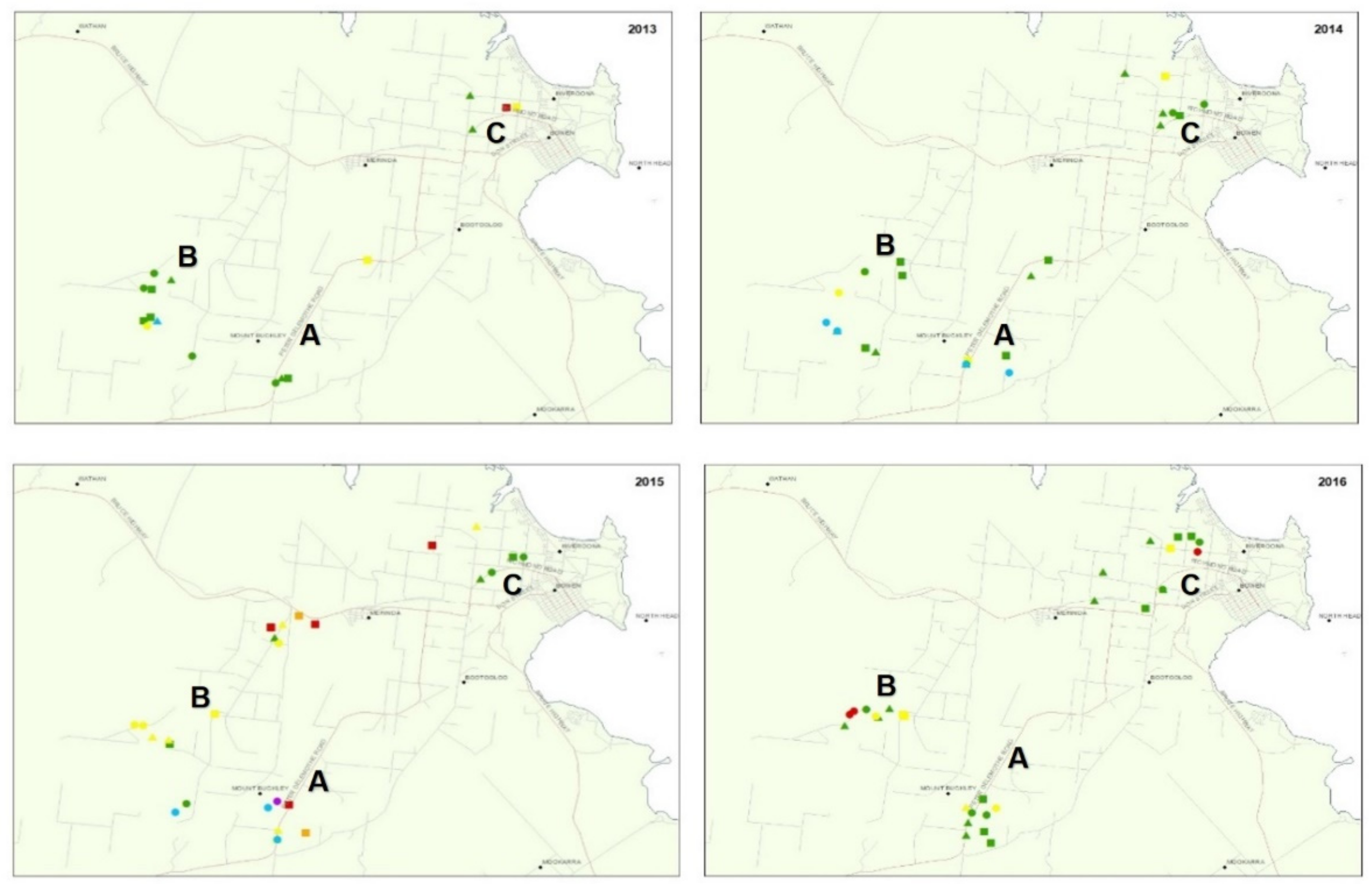
2.2. TYLCV Monitoring in Crops and Whitefly Populations
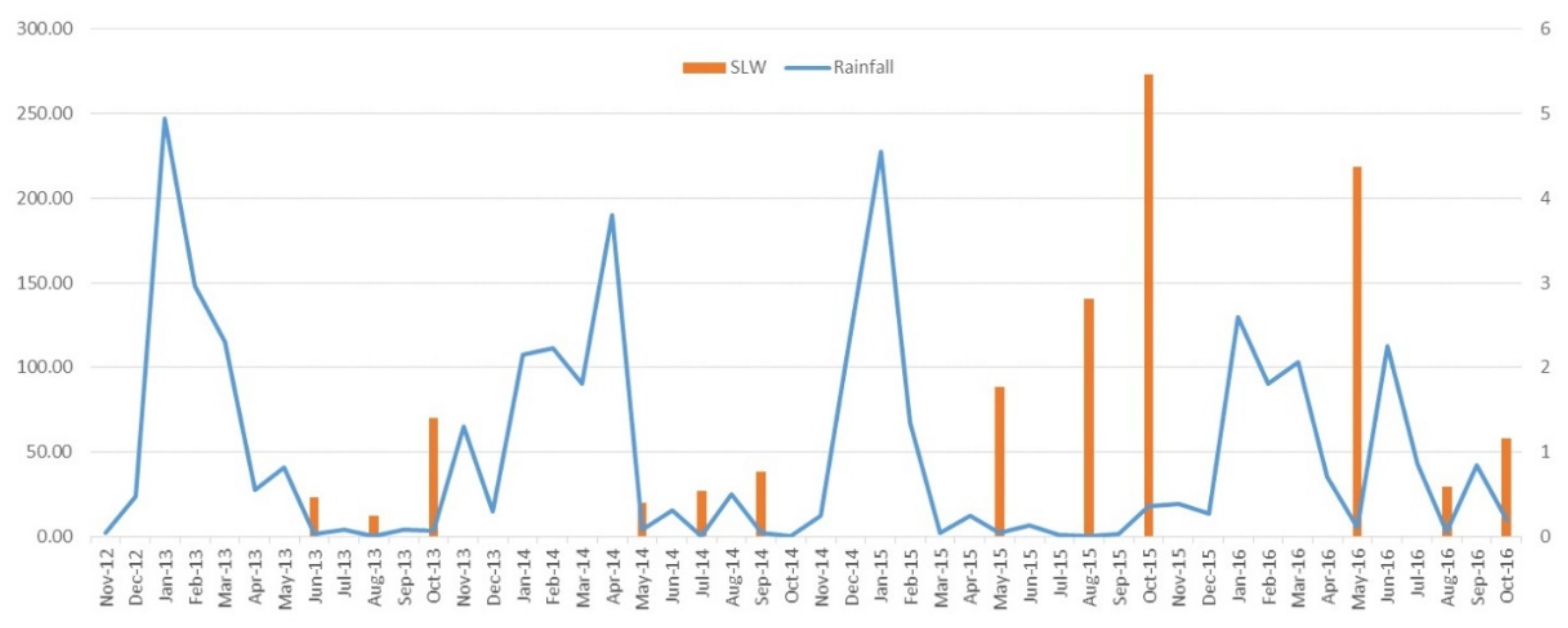
2.3. Whitefly Numbers and Rainfall
3. Discussion
4. Materials and Methods
4.1. Genetic Diversity of TYLCV
4.2. TYLCV and Vector Monitoring
4.3. Sample Indexing
4.4. Weather Monitoring
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICTV Genus: Begomovirus-Geminiviridae-ssDNA Viruses. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/ssdna-viruses/w/geminiviridae/392/genus-begomovirus (accessed on 21 February 2020).
- Varma, A.; Malathi, V.G. Emerging Geminivirus Problems: A Serious Threat to Crop Production. Ann. Appl. Biol. 2003, 142, 145–164. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olive, E.; Sánchez-Campos, S. Emerging Virus Diseases Transmitted by Whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Gunning, R.V.; Byrne, F.J.; Condé, B.D.; Connelly, M.I.; Hergstrom, K.; Devonshire, A.L. First Report of B-Biotype Bemisia Tabaci (Gennadius) (Hemiptera: Aleyrodidae) in Australia. Aust. J. Entomol. 1995, 34, 116. [Google Scholar] [CrossRef]
- Wongnikong, W.; Hereward, J.P.; van Brunschot, S.L.; Cappadonna, J.K.; Walter, G.H. Assessment of Relative Host Plant Quality for Three Cryptic Species of the Bemisia Tabaci Species Complex in Australia. Arthropod-Plant Interact. 2021, 15, 845–859. [Google Scholar] [CrossRef]
- Barro, P.J.; Hart, P.J. Mating Interactions between Two Biotypes of the Whitefly, Bemisia Tabaci (Hemiptera: Aleyrodidae) in Australia. Bull. Entomol. Res. 2000, 90, 103–112. [Google Scholar] [CrossRef]
- Van Brunschot, S.L.; Persley, D.M.; Geering, A.D.W.; Campbell, P.R.; Thomas, J.E. Tomato Yellow Leaf Curl Virus in Australia: Distribution, Detection and Discovery of Naturally Occurring Defective DNA Molecules. Australas. Plant Pathol. 2010, 39, 412–423. [Google Scholar] [CrossRef]
- Hohnle, M.; Hofer, P.; Bedford, I.D.; Briddon, R.W.; Markham, P.G.; Frischmuth, T. Exchange of Three Amino Acids in the Coat Protein Results in Efficient Whitefly Transmission of a Nontransmissible Abutilon Mosaic Virus Isolate. Virology 2001, 290, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, K.; Ogawa, T.; Sharma, P.; Ikegami, M. First Report of Honeysuckle Yellow Vein Mosaic Virus on Tomato Affected by Yellow Dwarf Disease in Japan. Plant Pathol. 2008, 57, 391. [Google Scholar] [CrossRef]
- Davis, R.I.; Jones, L.M.; Pease, B.; Perkins, S.L.; Vala, H.R.; Kokoa, P.; Apa, M.; Dale, C.J. Plant Virus and Virus-like Disease Threats to Australia’s North Targeted by the Northern Australia Quarantine Strategy. Plants 2021, 10, 2175. [Google Scholar] [CrossRef]
- Pacheco, I. Detection and Distribution of Geminiviruses in Mexico and the Southern United States. Phytopathology 1996, 86, 1186. [Google Scholar] [CrossRef]
- Polston, J.E.; Anderson, P.K. The Emergence Of Whitefly-Transmitted Geminiviruses in Tomato in the Western Hemisphere. Plant Dis. 1997, 81, 1358–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polston, J.E.; McGovern, R.J.; Brown, L.G. Introduction of Tomato Yellow Leaf Curl Virus in Florida and Implications for the Spread of This and Other Geminiviruses of Tomato. Plant Dis. 1999, 83, 984–988. [Google Scholar] [CrossRef] [Green Version]
- Seal, S.E.; Jeger, M.J.; Van den Bosch, F.; Karl Maramorosch, A.J.S.; Thresh, J.M. Begomovirus Evolution and Disease Management. Adv. Virus Res. 2006, 67, 297–316. [Google Scholar]
- Brown, J.K. The Bemisia Tabaci Complex: Genetic and Phenotypic Variability Drives Begomovirus Spread and Virus Diversification. APSnet Featur. 2007, 1, 25–56. [Google Scholar] [CrossRef]
- Lefeuvre, P.; Martin, D.P.; Harkins, G.; Lemey, P.; Gray, A.J.A.; Meredith, S.; Lakay, F.; Monjane, A.; Lett, J.-M.; Varsani, A.; et al. The Spread of Tomato Yellow Leaf Curl Virus from the Middle East to the World. PLoS Pathog. 2010, 6, e1001164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenyon, L.; Tsai, W.-S.; Shih, S.-L.; Lee, L.-M. Emergence and Diversity of Begomoviruses Infecting Solanaceous Crops in East and Southeast Asia. Virus Res. 2014, 186, 104–113. [Google Scholar] [CrossRef]
- Pan, L.-L.; Cui, X.-Y.; Chen, Q.-F.; Wang, X.-W.; Liu, S.-S. Cotton Leaf Curl Disease: Which Whitefly Is the Vector? Phytopathology 2018, 108, 1172–1183. [Google Scholar] [CrossRef] [Green Version]
- Péréfarres, F.; Thierry, M.; Becker, N.; Lefeuvre, P.; Reynaud, B.; Delatte, H.; Lett, J.-M. Biological Invasions of Geminiviruses: Case Study of TYLCV and Bemisia Tabaci in Reunion Island. Viruses 2012, 4, 3665–3688. [Google Scholar] [CrossRef] [Green Version]
- Czosnek, H.; Ghanim, M. Bemisia Tabaci–Tomato Yellow Leaf Curl Virus Interaction Causing Worldwide Epidemics. In The Whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) Interaction with Geminivirus-Infected Host Plants; Springer: Dordrecht, The Netherlands, 2011; pp. 51–67. ISBN 978-94-007-1523-3. [Google Scholar]
- Campbell, P.R.; Cremer, J.E.; Roach, R.L.; Steele, V.; Subramaniam, S.; Sivasubramaniam, V.; Monsour, C.; Mullins, T.; Persley, D.M.; Gambley, C.F. Towards Area Wide Management of Insect Vectored Viruses of Tomatoes in the Bowen District. Virus Res. 2017, 241, 228–235. [Google Scholar] [CrossRef]
- Lapidot, M.; Friedmann, M.; Lachman, O.; Yehezkel, A.; Nahon, S.; Cohen, S.; Pilowsky, M. Comparison of Resistance Level to Tomato Yellow Leaf Curl Virus Among Commercial Cultivars and Breeding Lines. Plant Dis. 1997, 81, 1425–1428. [Google Scholar] [CrossRef] [Green Version]
- Lapidot, M.; Friedmann, M.; Pilowsky, M.; Ben-Joseph, R.; Cohen, S. Effect of Host Plant Resistance to Tomato Yellow Leaf Curl Virus (TYLCV) on Virus Acquisition and Transmission by Its Whitefly Vector. Phytopathology 2001, 91, 1209–1213. [Google Scholar] [CrossRef] [Green Version]
- Legarrea, S.; Barman, A.; Marchant, W.; Diffie, S.; Srinivasan, R. Temporal Effects of a Begomovirus Infection and Host Plant Resistance on the Preference and Development of an Insect Vector, Bemisia Tabaci, and Implications for Epidemics. PLoS ONE 2015, 10, e0142114. [Google Scholar]
- Srinivasan, R.; Riley, D.; Diffie, S.; Sparks, A.; Adkins, S. Whitefly Population Dynamics and Evaluation of Whitefly-Transmitted Tomato Yellow Leaf Curl Virus (TYLCV)-Resistant Tomato Genotypes as Whitefly and TYLCV Reservoirs. J. Econ. Enotomol. 2012, 105, 1447–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macedo, M.A.; Costa, T.M.; Barbosa, J.C.; Pereira, J.L.; Michereff-Filho, M.; Gilbertson, R.L.; Inoue-Nagata, A.K.; Bergamin Filho, A. Temporal and Spatial Dynamics of Begomovirus Disease in Tomatoes in Central Brazil. Plant Pathol. 2017, 66, 529–538. [Google Scholar] [CrossRef]
- De Barro, P.J.; Driver, F. Use of RAPD PCR to Distinguish the B Biotype from Other Biotypes of Bemisia Tabaci (Gennadius) (Hemiptera: Aleyrodidae). Aust. J. Entomol. 1997, 36, 149–152. [Google Scholar] [CrossRef]
- Moran, J.; Garrett, R.G.; Fairweather, J.V. Strategy for Detecting Low Levels of Potato Viruses X and S in Crops and Its Application to the Victorian Certified Potato Scheme. Plant Dis. 1983, 67, 1325–1328. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.J.; Sokal, R.R. Statistical Tables; W. H. Freeman and Company Ltd.: Folkestone, UK, 1969. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Survey Month and Year | Survey Area | Survey Site | Varietal Susceptibility (S) or Resistance (R) 1 to TYLCV | TYLCV Incidence 2 | SLW Population Size per Leaflet 4 | |
|---|---|---|---|---|---|---|
| Crop (%) 3a | SLW (%) 3a | |||||
| Jun-13 | A | 272 | nd | 0.3 | 0.0 | 0.2 |
| B | 266 | nd | 1.4 | 1.0 | 0.4 | |
| 267 | nd | 2.6 | 0.0 | 1.1 | ||
| 268 | nd | 0.3 | 0.6 | 0.3 | ||
| 273 | S | 1.4 | 1.5 | 0.3 | ||
| Aug-13 | A | 283 | nd | 0.0 | 0.0 | 0.2 |
| B | 277 | R | 2.6 | 6.0 | 0.4 | |
| 278 | R | 1.4 | 57.5 | 0.1 | ||
| 280 | R | 5.5 | 13.0 | 0.3 | ||
| C | 275 | S | 0.3 | 1.0 | 0.2 | |
| 284 | nd | 2.2 | 20.0 | 0.2 | ||
| Oct-13 | A | 297 | S | 100.0 | 40.0 | 1.5 |
| 298 | R | 3.5 | 4.0 | 0.6 | ||
| B | 294 | S | 100.0 | 99.0 | 0.4 | |
| 295 | R | 100.0 | 49.0 | 0.6 | ||
| 296 | nd | 100.0 | 95.0 | 0.2 | ||
| C | 290 | R | 12.4 | 67.0 | 1.3 | |
| 291 | nd | 20.6 | 97.8 | 5.4 | ||
| May-14 | A | 300 | R | 0.4 (0.01–1.9) | 0.0 | 0.1 |
| 301 | R | 0.0 | 0.0 | 0.1 | ||
| B | 302 | S | 2.2 (0.8–4.7) | 2.2 | 0.1 | |
| 303 | S | 1.4 (0.4–3.6) | 0.0 | 0.1 | ||
| 305 | S | 11.3 (6.8–17.4) | 32.0 | 1.2 | ||
| 306 | S | 0.4 (0.01–1.9) | 8.0 | 0.9 | ||
| C | 307 | nd | 0.0 | 0.0 | 0.4 | |
| 308 | nd | 0.7 (0.1–2.5) | 0.0 | 0.3 | ||
| July-14 | A | 359 | R | 0.0 | 0.0 | 0.01 |
| 360 | R | 0.4 (0.01–1.9) 3b | 35.0 | 0.7 | ||
| 361 | S | 0.0 | 16.0 | 0.6 | ||
| B | 355 | R | 0.0 | 1.3 | 0.1 | |
| 356 | S | 14.9 (9.1–22.6) | 28.0 | 0.4 | ||
| 357 | R | 28.8 (16.1–51.0) | 38.3 | 0.1 | ||
| 358 | S | 0.4 (0.01–1.9) | 6.0 | 2.0 | ||
| C | 352 | S | 0.0 | 0.0 | 0.2 | |
| 353 | S | 0.4 (0.01–1.9) | 0.0 | 0.1 | ||
| 354 | S | 2.2 (0.8–4.7) | 2.7 | 0.1 | ||
| Sept-14 | A | 382 | R | 0.0 | 6.0 | 3.0 |
| 383 | R | 0.0 | 0.0 | 0.4 | ||
| 384 | R | 0.4 (0.1–1.9) | 0.0 | 0.1 | ||
| B | 378 | R | 0.0 | 3.1 | 0.2 | |
| 381 | R | 6.1 (3.3–10.1) | 31.0 | 0.6 | ||
| 385 | R | 23.7 (14.0–38.1) | 0.0 | 0.5 | ||
| C | 376 | R | 0.5 (0.01–2.8) 3b | 38.0 | 1.7 | |
| 377 | R | 0.0 | 34.0 | 0.5 | ||
| May-15 | A | 542 | R | 0.0 | 0.0 | 0.01 |
| 543 | R | 0.0 | 0.0 | 0.0 | ||
| 544 | R | 0.0 | 0.0 | 0.01 | ||
| B | 537 | R | 0.0 | 0.0 | 0.3 | |
| 538 | R | 0.0 | 20.0 | 0.01 | ||
| 539 | S | 0.0 | 0.0 | 3.0 | ||
| 540 | S | 3.0 (1.3–6.0) | 8.3 (3.3–16.0) 3c | 1.5 | ||
| 541 | R | 0.0 | 0.0 3c | 2.0 | ||
| 545 | S | 12.4 (7.5–18.9) | 16.0 3d | 2.0 | ||
| C | 535 | S | 0.0 | 1.5 | 0.1 | |
| 536 | R | 0.0 | 0.0 3c | 0.5 | ||
| Aug-15 | A | 552 | R | 0.4 (0.01–1.9) | 1.0 (0.3–5.6) 3c | 3.0 |
| 553 | R | 0.0 | 0.0 3c | 1.5 | ||
| B | 550 | R | 4.0 (1.9–7.2) | 0.0 3c | 3.0 | |
| 551 | R | 8.8 (5.1–13.8) | 2.1 (0.3–7.3) 3c | 2.0 | ||
| 554 | S | 27.5 (14.7–49.9) | 2.1 (0.3–7.3) 3c | 2.0 | ||
| 555 | S | 13.5 (8.3–20.6) | 36.0 3d | 0.7 | ||
| C | 556 | R | 0.0 | 1.0 (0.3–5.6) 3c | 1.5 | |
| 557 | R | 0.0 | 0.0 | 0.1 | ||
| Oct-15 | A | 564 | R | 0.0 | 0.0 | 0.2 |
| 565 | R | 0.0 | 1.0 (0.3–5.6) 3c | 2.3 | ||
| B | 558 | R | 28.8 (16.1–51.0) | 5.6 (1.8–12.6) 3c | 9.4 | |
| 559 | R | 13.5 (8.3–20.6) | 0.0 3c | 9.4 | ||
| 560 | R | 14.9 (9.1–22.6) | 0.0 3c | 4.7 | ||
| 561 | R | 100 (19.4–100.0) | 8.3 (3.3–16.0) 3c | 7.8 | ||
| 562 | R | 10.4 (6.2–16.1) | 2.1 (0.3–7.3) | 4.7 | ||
| 563 | R | 5.5 (2.9–9.4) | 2.1 (0.3–7.3) 3c | 7.8 | ||
| C | 566 | R | 0.0 | 2.1 (0.3–7.3) 3c | 7.8 | |
| 567 | R | 0.0 | 1.0 (0.3–5.6) 3c | 0.5 | ||
| May-16 | A | 577 | R | 0.0 | 0.0 | 0.5 |
| 578 | R | 0.0 | 0.0 | 1.5 | ||
| 580 | R | 0.3 (0.01–1.9) | 0.0 | 0.8 | ||
| B | 572 | R | 0.3 (0.01–1.9) | 0.0 | 1.9 | |
| 573 | R | 0.0 | 0.0 | 0.2 | ||
| 574 | S | 6.1 (3.3–10.1) | 0.0 | 12.1 | ||
| 576 | S | 0.7 (0.1–2.5) | 0.0 | 11.5 | ||
| C | 571 | S | 0.0 | 0.0 | 11.5 | |
| 579 | S | 0.0 | 0.0 | 0.1 | ||
| 582 | S | 0.0 | 0.0 | 1.0 | ||
| Aug-16 | A | 583 | R | 1.05 (0.2–3.0) | 0.0 | 3.0 |
| 584 | R | 0.0 | 0.0 | 0.4 | ||
| 585 | R | 3.05 (1.3–6.0) | 0.0 | 0.5 | ||
| B | 586 | R | 0.0 | 00.0 | 0.2 | |
| 587 | R | 0.0 | 1.02 (0.03–5.5) | 0.6 | ||
| 588 | R | 1.8 (0.6–4.2) | 0.0 | 0.7 | ||
| C | 589 | R | 7.3 (4.1–11.8) | 0.0 5 | 0.5 | |
| 590 | R | 9.5 (5.6–14.9) | 0.0 5 | 0.9 | ||
| 591 | S | 14.9 (9.1–22.6) | 0.0 5 | 0.9 | ||
| 592 | nd | 18.2 (11.1–27.9) | 0.0 5 | 0.4 | ||
| Oct-16 | A | 599 | R | 5.0 (2.5–8.6) | 3.2 (0.6–9.1) | 0.8 |
| 600 | R | 5.5 (2.9–9.4) | 8.2 (3.3–16.4) | 0.7 | ||
| 601 | R | 2.6 (1.0–5.3) | 4.4 (1.2–10.8) | 0.4 | ||
| B | 606 | R | 23.7 (14.0–38.1) | 0.0 5 | 1.5 | |
| 607 | R | 23.7 (14.0–38.1) | 0.0 5 | 1.8 | ||
| 608 | R | 0.34 (0.01–1.87) | 0.0 | 3.0 | ||
| C | 602 | R | 7.34 (4.12–11.84) | 5.6 (1.8–12.6) | 1.4 | |
| 603 | R | 0.34 (0.01–1.87) | 11.3 (5.1–26.6) | 0.6 | ||
| 604 | R | 1.42 (0.38–3.61) | 1.0 (0.03–5.5) | 0.4 | ||
| 605 | R | 28.8 (16.1–51.0) | 0.0 | 0.2 | ||
| Year | Season | Total Survey Sites | Prevalence of TYLCV 1 | TYLCV Incidence in Crops (%) at Each Study Location within the District | Average Viruliferous SLW (%) | TYLCV Incidence in SLW (%) across the District | Average Number of SLW per Leaflet 2 | Number of SLW per Leaflet 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | A | B | C | A | B | C | ||||||
| 2013 | Early-June | 5 | 5 | 0.34 | 0.3–2.6 | Nd 1 | 0.6 | 0.0 | 0.0–1.5 | nd 1 | 0.5 | 0.2 | 0.3–1.1 | nd 1 |
| Mid-August | 6 | 5 | 0.0 | 1.4–5.5 | 0.3–2.2 | 16.2 | 0.0 | 6.0–57.5 | 1.0–20.0 | 0.2 | 0.2 | 0.1–0.4 | 0.2–0.2 | |
| Late-October | 7 | 7 | 3.5–100.0 | 100.0 | 12.4–20.6 | 64.5 | 4.0–40.0 | 49.0–99.0 | 67.0–97.8 | 1.4 | 0.6–1.5 | 0.2–0.6 | 1.3–5.4 | |
| 2014 | Early-May | 8 | 6 | 0.0–0.4 | 0.4–11.3 | 0.0–0.7 | 5.1 | 0.0 | 0.0–32.0 | 0.0 | 0.4 | 0.1 | 0.1–1.2 | 0.3–0.4 |
| Mid-July | 10 | 6 | 0.0–0.4 | 0.0–28.8 | 0.0–2.2 | 15.9 | 0.0–35.0 | 1.3–38.3 | 0.0–2.7 | 0.5 | 0.01–0.7 | 0.1–2.0 | 0.1–0.2 | |
| Late-September | 8 | 4 | 0.0–0.4 | 0.0–23.7 | 0.0–0.5 | 16.0 | 0.0–6.0 | 0.0–31.0 | 34.0–38.0 | 0.8 | 0.1–3.0 | 0.2–0.6 | 0.5–1.7 | |
| 2015 | Early-May | 11 | 2 | 0.0 | 0.0–12.4 | 0.0 | 3.7 | 0.0 | 0.0–20.0 | 0.0–1.5 | 1.8 | 0.0–0.01 | 0.01–3.0 | 0.1–0.5 |
| Mid-August | 8 | 5 | 0.0–0.4 | 4.0–27.5 | 0.0 | 6.0 | 0.0–1.0 | 0.0–36.0 | 0.0–1.0 | 2.8 | 1.5–3.0 | 0.7–3.0 | 0.1–1.5 | |
| Late-October | 10 | 6 | 0.0 | 5.5–100.0 | 0.0 | 2.2 | 0.0–1.0 | 0.0–8.3 | 1.0–2.1 | 5.4 | 0.2–2.3 | 4.7–9.4 | 0.5–7.8 | |
| 2016 | Early-May | 10 | 4 | 0.0–0.3 | 0.0–6.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.37 | 0.8–1.5 | 0.2–12.1 | 0.1–11.5 |
| Mid-August | 10 | 7 | 0.0–3.0 | 0.0–1.8 | 7.3–18.2 | 0.1 | 0.0 | 0.0–1.0 | 0.0 | 0.59 | 0.4–3.0 | 0.2–0.7 | 0.4–0.9 | |
| Late-October | 10 | 10 | 2.6–5.5 | 0.3–23.7 | 0.3–28.8 | 9.9 | 3.2–8.2 | 0.0 | 0.0–11.3 | 1.16 | 0.4–0.8 | 1.5–3.0 | 0.2–1.4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambley, C.; Nimmo, P.; McDonald, J.; Campbell, P. The Establishment and Spread of a Newly Introduced Begomovirus in a Dry Tropical Environment Using Tomato Yellow Leaf Curl Virus as a Case Study. Plants 2022, 11, 776. https://doi.org/10.3390/plants11060776
Gambley C, Nimmo P, McDonald J, Campbell P. The Establishment and Spread of a Newly Introduced Begomovirus in a Dry Tropical Environment Using Tomato Yellow Leaf Curl Virus as a Case Study. Plants. 2022; 11(6):776. https://doi.org/10.3390/plants11060776
Chicago/Turabian StyleGambley, Cherie, Peter Nimmo, Janet McDonald, and Paul Campbell. 2022. "The Establishment and Spread of a Newly Introduced Begomovirus in a Dry Tropical Environment Using Tomato Yellow Leaf Curl Virus as a Case Study" Plants 11, no. 6: 776. https://doi.org/10.3390/plants11060776