
The Comparison of Soil Agrochemical and Biological Properties in the Multi-Cropping Farming Systems

Abstract
:1. Introduction
2. Results
2.1. Total Nitrogen Content in the Soil
2.2. Organic Carbon Content in the Soil
2.3. Available Phosphorus Content in the Soil
2.4. Available Potassium Content in the Soil
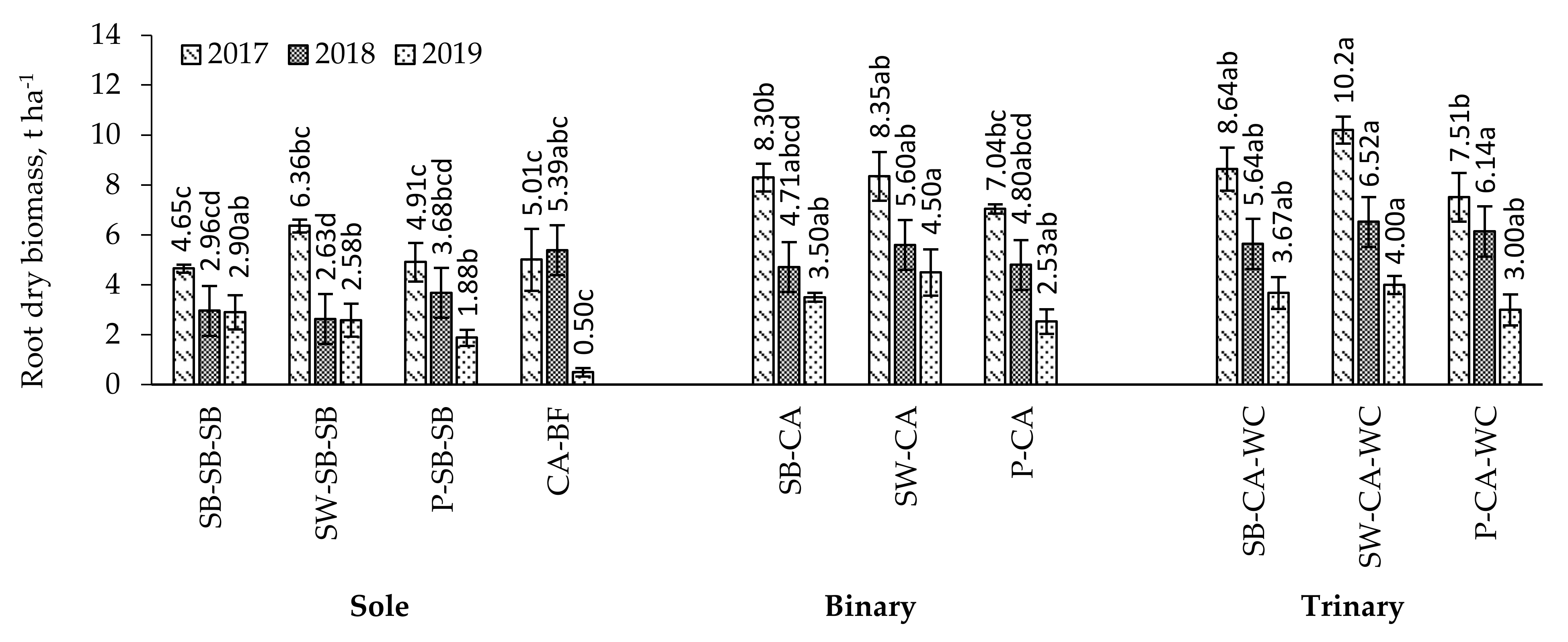
2.5. Root Dry Biomass
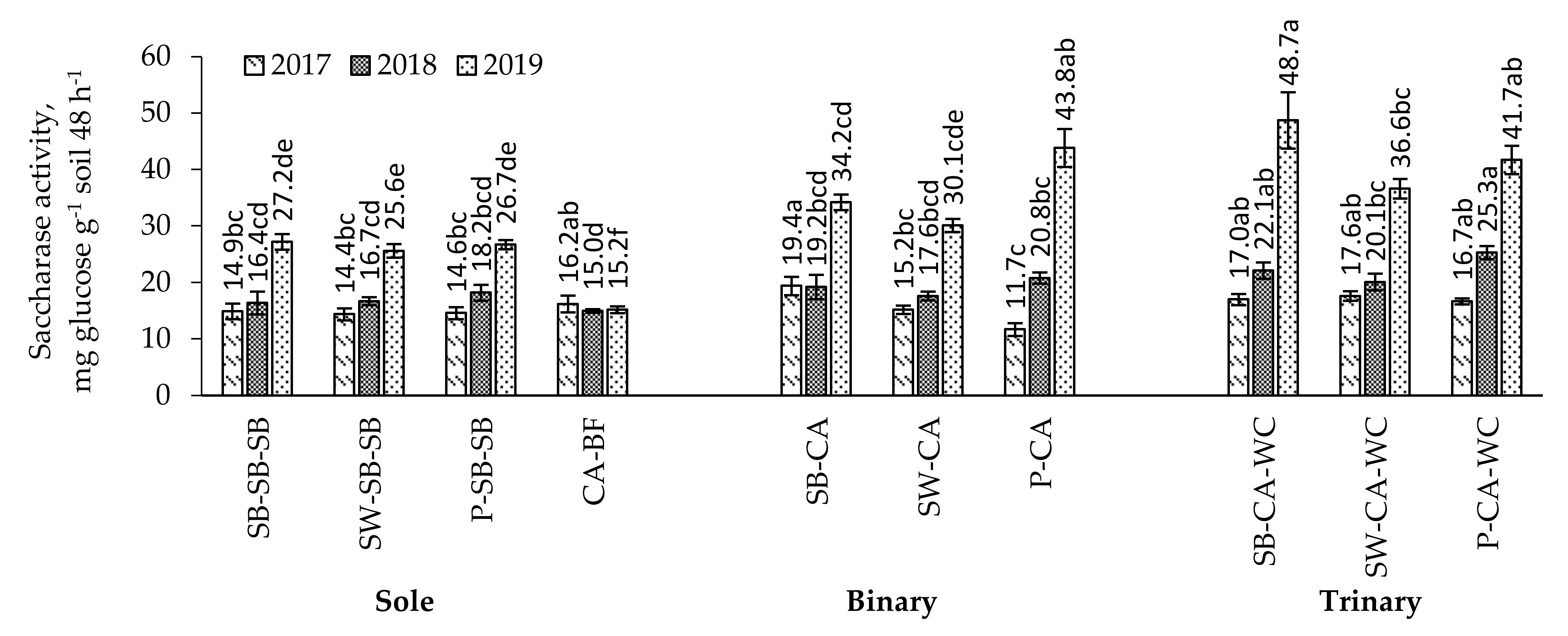
2.6. Saccharase Activity in the Soil
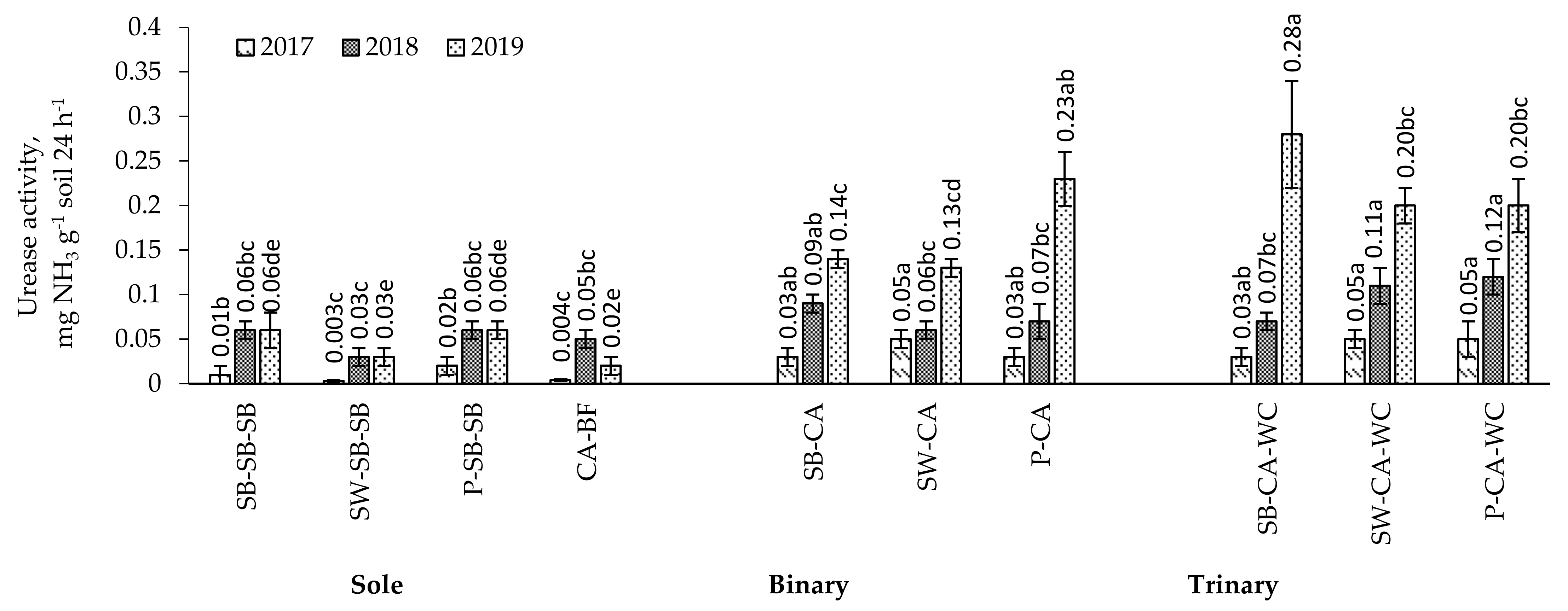
2.7. Urease Activity in the Soil
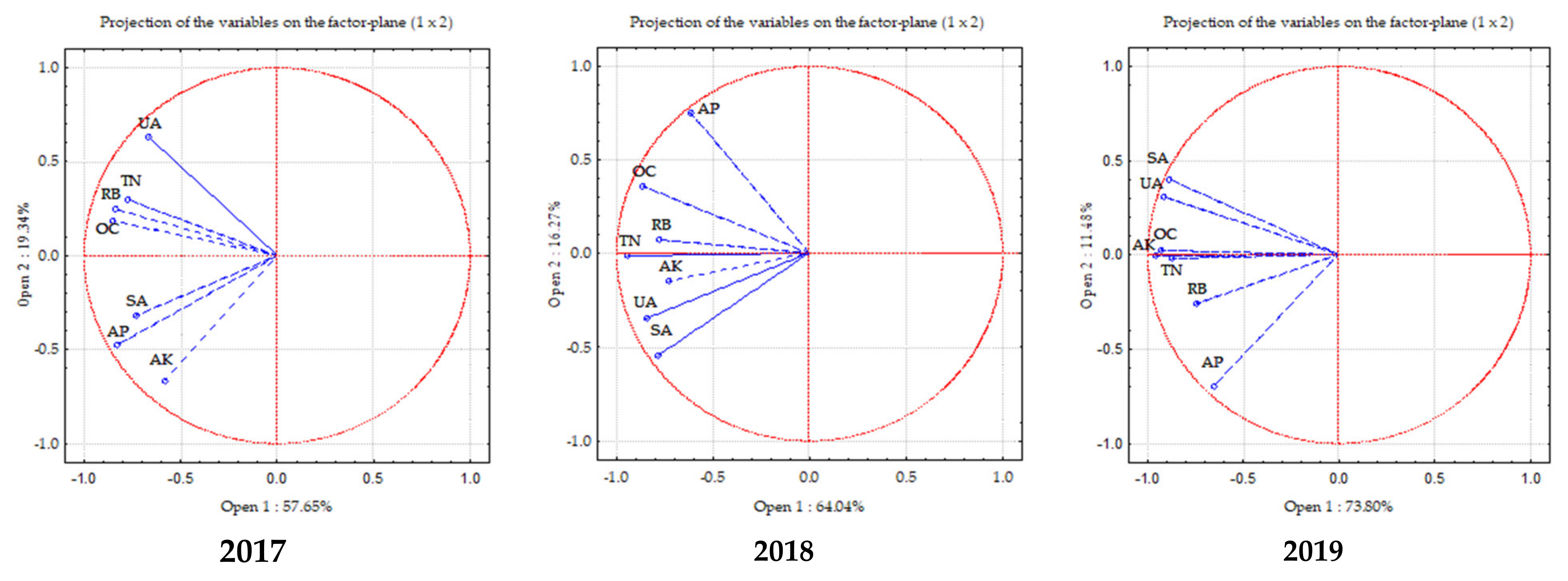
2.8. Principal Component Analysis (PCA)
3. Discussion
3.1. N, OC, P, K Contents in the Multi-Cropping System
3.2. Root Biomass in the Multi-Cropping System
3.3. Soil Enzyme Activity in the Multi-Cropping System
4. Materials and Methods
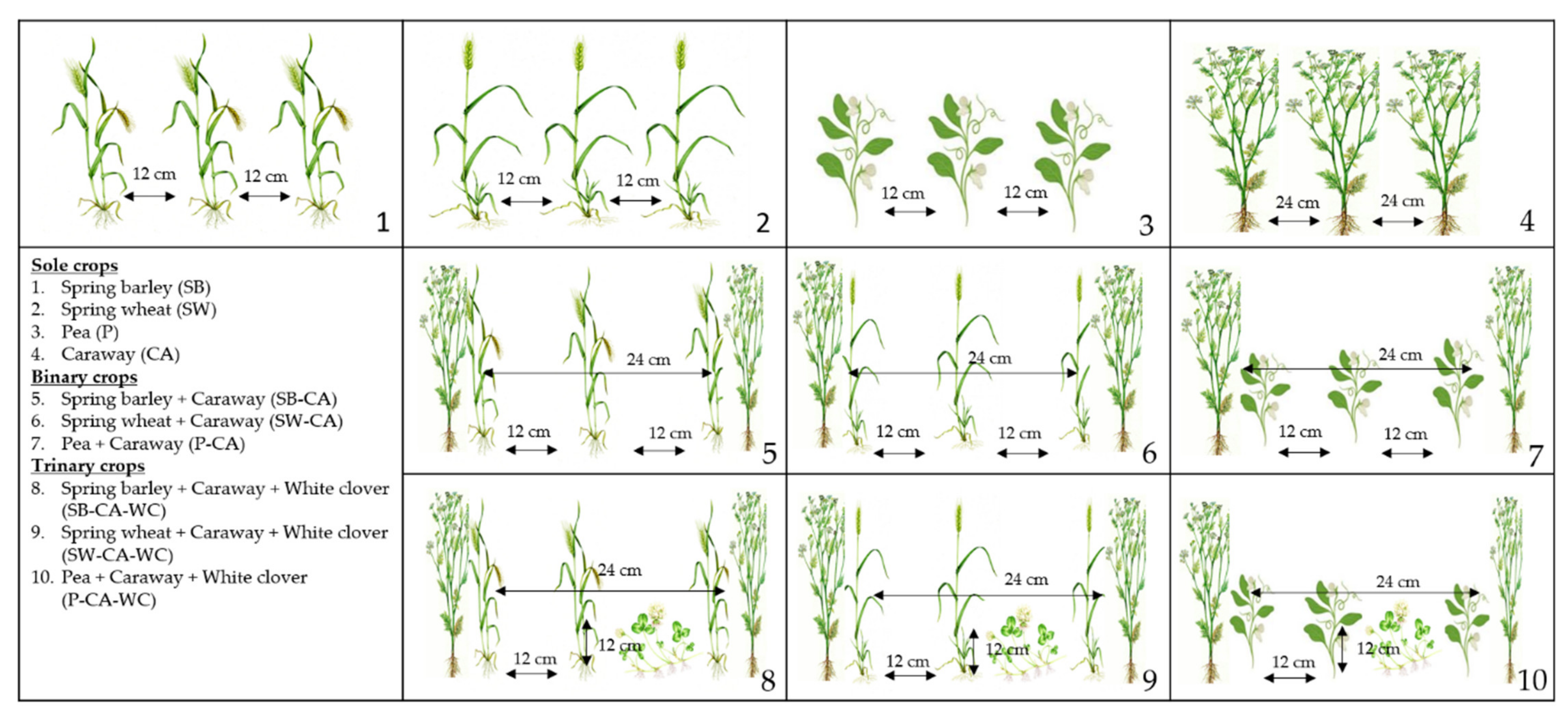
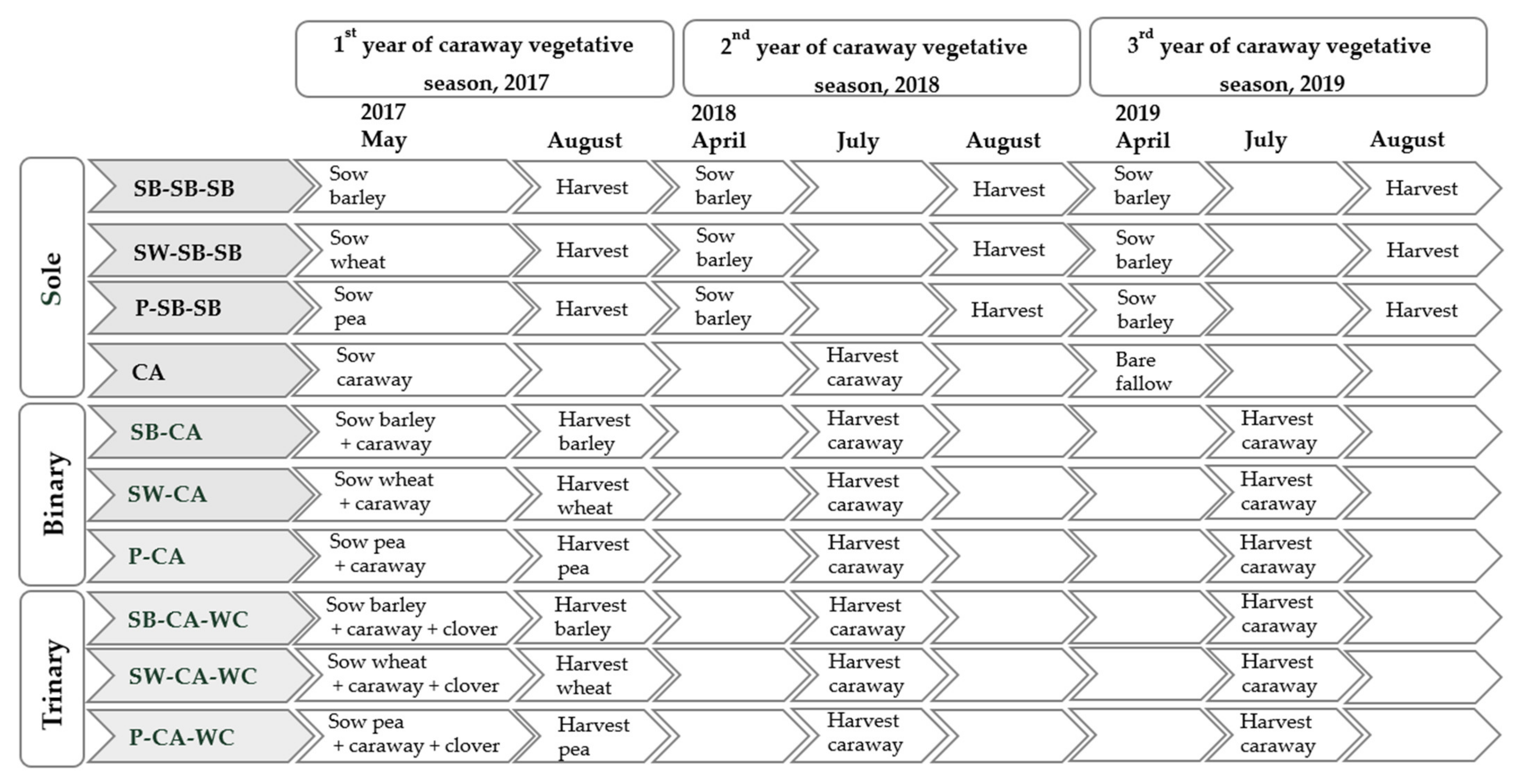
4.1. General Experimental Conditions
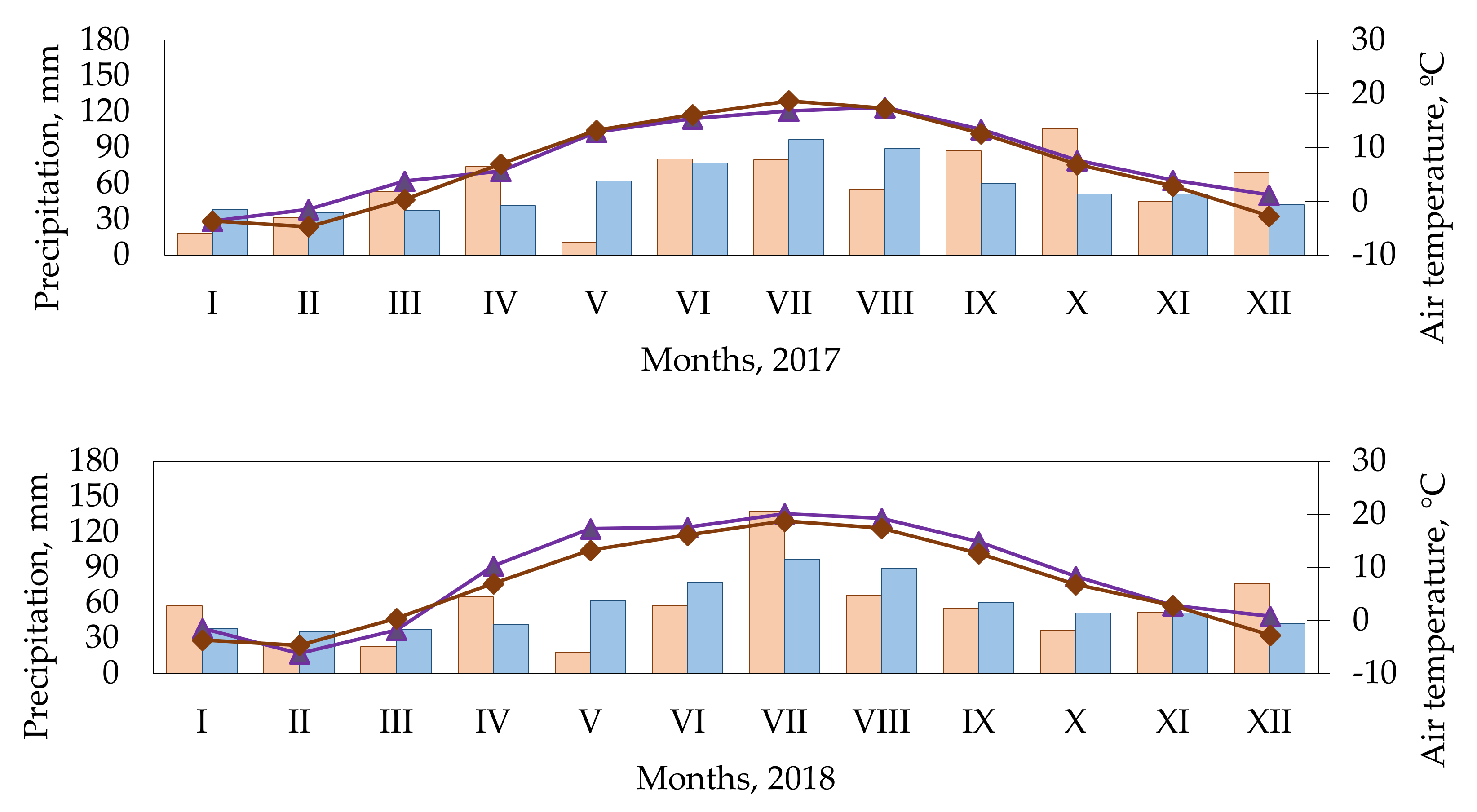
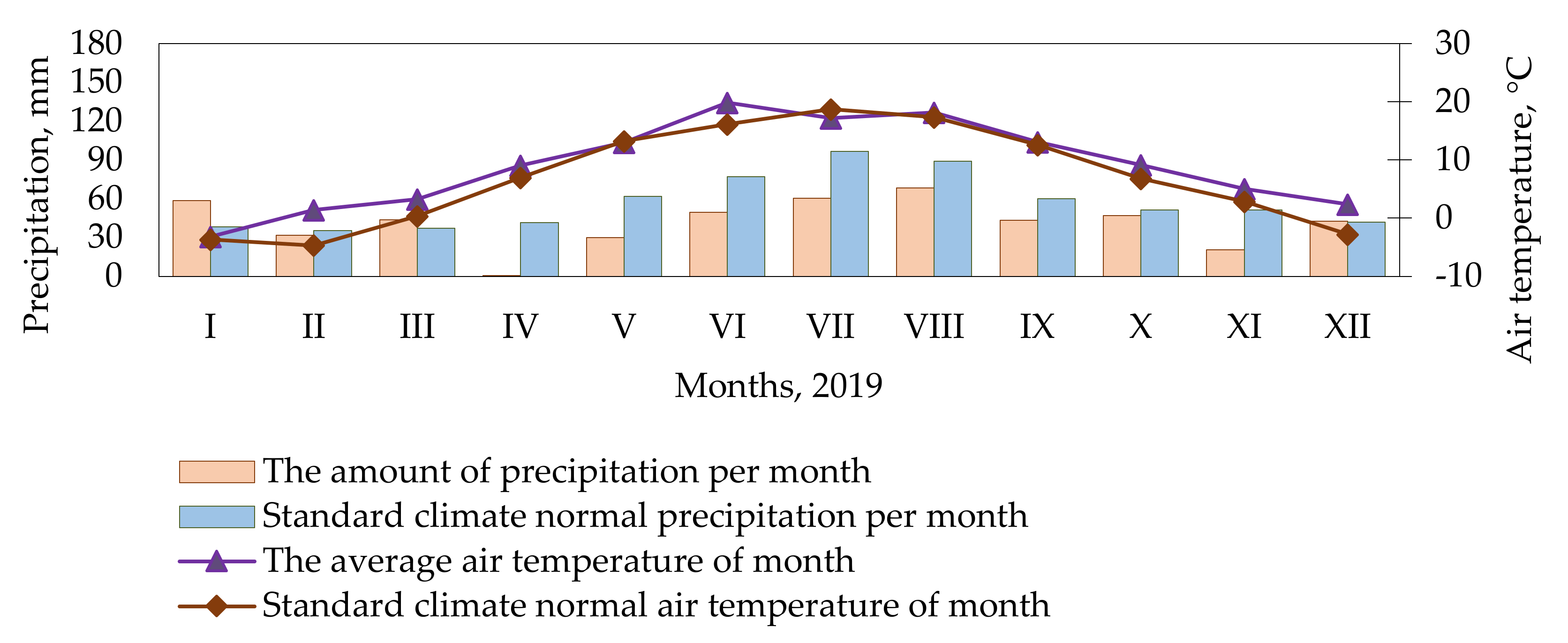
4.2. Meteorological Conditions
4.3. Analytical Methods
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hertel, T.W. The global supply and demand for agricultural land in 2050: A perfect storm in the making? Am. J. Agric. Econ. 2011, 93, 259–275. [Google Scholar] [CrossRef]
- Arora, N.K.; Fatima, T.; Mishra, I.; Verma, M.; Mishra, J.; Mishra, V. Environmental sustainability: Challenges and viable solutions. Environ. Sustain. 2018, 1, 309–340. [Google Scholar] [CrossRef]
- Naulleau, A.; Gary, C.; Prévot, L.; Hossard, L. Evaluating strategies for adaptation to climate change in grapevine production–A systematic review. Front. Plant Sci. 2021, 11, 2154. [Google Scholar] [CrossRef] [PubMed]
- Keating, B.A.; Carberry, P.S.; Bindraban, P.S.; Asseng, S.; Meinke, H.; Dixon, J. Eco-efficient agriculture: Concepts, challenges, and opportunities. Crop Sci. 2010, 50, 109–119. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, Y.; Qi, L.; Mei, X.; Liao, J.; Ding, X.; Deng, W.; Fan, L.; He, X.; Vivanco, J.M.; et al. Plant-Plant-Microbe Mechanisms Involved in Soil-Borne Disease Suppression on a Maize and Pepper Intercropping System. PLoS ONE 2014, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Crusciol, C.A.C.; Gustavo, M.P.; Momesso, L.; Pariz, C.M.; Castilhos, A.M.; Calonego, J.C.; Borghi, E.; Costa, C.; Franzluebbers, A.J.; Cantarella, H. Nitrogen-fertilized systems of maize intercropped with tropical grasses for enhanced yields and estimated land use and meat production. Front. Sustain. Food Syst. 2020, 4, 211–224. [Google Scholar]
- Wang, Z.; Zhao, X.; Wu, P.; He, J.; Chen, X.; Gao, Y.; Cao, X. Radiation interception and utilization by wheat/maize strip intercropping systems. Agric. For Meteorol. 2015, 204, 58–66. [Google Scholar] [CrossRef]
- Petrie, C.A.; Bates, J. ‘Multi-cropping’, intercropping and adaptation to variable environments in Indus South Asia. J. World Prehist. 2017, 30, 81–130. [Google Scholar] [CrossRef]
- Zanetti, F.; Alberghini, B.; Jeromela, A.M.; Grahovac, N.; Rajković, D.; Kiprovski, B.; Monti, A. Camelina, an ancient oilseed crop actively contributing to the rural renaissance in Europe. Agron Sustain. Dev. 2021, 41, 1–18. [Google Scholar] [CrossRef]
- Maitra, S.; Hossain, A.; Brestic, M.; Skalicky, M.; Ondrisik, P.; Gitari, H.; Brahmachari, K.; Shankar, T.; Bhadra, P.; Palai, J.B.; et al. Intercropping—A Low Input Agricultural Strategy for Food and Environmental Security. Agronomy 2021, 11, 343. [Google Scholar] [CrossRef]
- Xiao, X.; Cheng, Z.; Meng, H.; Khan, M.A.; Li, H. Intercropping with garlic alleviated continuous cropping obstacle of cucumber in plastic tunnel. Acta Agric. Scand B Soil Plant Sci. 2012, 62, 696–705. [Google Scholar] [CrossRef]
- Lizarazo, C.I.; Tuulos, A.; Jokela, V.; Mäkelä, P.S. Sustainable mixed cropping systems for the boreal-nemoral region. Front. Sustain. Food Syst. 2020, 4, 103. [Google Scholar] [CrossRef]
- Stöckle, C.O.; Donatelli, M.; Nelson, R. CropSyst, a cropping systems simulation model. Eur. J. Agron. 2003, 18, 289–307. [Google Scholar] [CrossRef]
- Hiddink, G.A.; Termorshuizen, A.J.; Bruggen, A.H. Mixed cropping and suppression of soilborne diseases. Genetic Engineering, biofertilisation, soil quality and organic farming. Sustain. Agric. 2010, 5, 119–146. [Google Scholar]
- Hiltbrunner, J.; Liedgens, M. Performance of winter wheat varieties in white clover living mulch. Biol. Agric. Hortic. 2008, 26, 85–101. [Google Scholar] [CrossRef] [Green Version]
- Dinnes, D.L.; Karlen, D.L.; Jaynes, D.B.; Kaspar, T.C.; Hatfield, J.L.; Colvin, T.S.; Cambardella, C.A. Nitrogen management strategies to reduce nitrate leaching in tile-drained midwestern soils. Agron. J. 2002, 94, 153–171. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Jensen, E. Facilitative root interactions in intercrops. Plant Soil. 2005, 274, 237–250. [Google Scholar] [CrossRef]
- Rao, C.S.; Grover, M.; Kundu, S.; Desai, S. Soil Enzymes. In Encyclopedia of Soil Science; Taylor and Francis: Abingdon, UK, 2017; pp. 2100–2107. [Google Scholar]
- Wahbi, S.; Prin, Y.; Thioulouse, J.; Sanguin, H.; Baudoin, E.; Maghraoui, T.; Oufdou, K.; Roux, C.L.; Galiana, A.; Hafidi, M.; et al. Impact of wheat/faba bean mixed cropping or rotation systems on soil microbial functionalities. Front. Plant Sci. 2016, 7, 1364. [Google Scholar] [CrossRef] [Green Version]
- Hirte, J.; Leifeld, J.; Abiven, S.; Oberholzer, H.R.; Hammelehle, A.; Mayer, J. Overestimation of crop root biomass in field experiments due to extraneous organic matter. Front. Plant Sci. 2017, 8, 284. [Google Scholar] [CrossRef] [Green Version]
- Mu-Chun, Y.; Ting-Ting, X.; Peng-Hui, S.; Jian-Jun, D. Effects of different cropping patterns of soybean and maize seedlings on soil enzyme activities and MBC and MBN. J. Northeast. Agric. Univ. 2012, 19, 42–47. [Google Scholar] [CrossRef]
- Ehrmann, J.; Ritz, K. Plant: Soil interactions in temperate multi-cropping production systems. Plant Soil. 2014, 376, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Mu, Y.; Cheng, Y.; Liu, X.; Nian, H. Effects of intercropping sugarcane and soybean on growth, rhizosphere soil microbes, nitrogen and phosphorus availability. Acta Physiol. Plant. 2013, 35, 1113–1119. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, M.S.; Kim, J.G.; Kim, S.O. Use of soil enzymes as indicators for contaminated soil monitoring and sustainable management. Sustainability 2020, 12, 8209. [Google Scholar] [CrossRef]
- Utobo, E.B.; Tewari, L. Soil enzymes as bioindicators of soil ecosystem status. Appl. Ecol. Environ. Res. 2015, 13, 147–169. [Google Scholar]
- Attademo, A.M.; Sanchez-Hernandez, J.C.; Lajmanovich, R.C.; Repetti, M.R.; Peltzer, P.M. Enzyme activities as indicators of soil quality: Response to intensive soybean and rice crops. Water Air Soil Pollut. 2021, 232, 1–12. [Google Scholar] [CrossRef]
- Trap, J.; Riah, W.; Akpa-Vinceslas, M.; Bailleul, C.; Laval, K.; Trinsoutrot-Gattin, I. Improved effectiveness and efficiency in measuring soil enzymes as universal soil quality indicators using microplate fluorimetry. Soil Biol. Biochem. 2012, 45, 98–101. [Google Scholar] [CrossRef]
- Datt, N.; Singh, D. Enzymes in Relation to Soil Biological Properties and Sustainability. In Sustainable Management of Soil and Environment; Meena, R., Kumar, S., Bohra, J., Jat, M., Eds.; Springer: Singapore, 2019; pp. 383–406. [Google Scholar] [CrossRef]
- Samuel, A.D.; Domuţa, C.; Ciobanu, C.; Şandor, M. Field management effects on soil enzyme activities. Rom. Agric. Res. 2008, 28, 61–68. [Google Scholar]
- Nath, C.P.; Kumar, N.; Das, K.; Hazra, K.K.; Praharaj, C.S.; Singh, N.P. Impact of variable tillage based residue management and legume based cropping for seven years on enzymes activity, soil quality index and crop productivity in rice ecology. Ecological Indicators. 2021, 10, 1–11. [Google Scholar] [CrossRef]
- Lizarazo, C.I.; Lampi, A.M.; Mäkelä, P.S. Can foliar-applied nutrients improve caraway (Carum carvi L.) seed oil composition? Ind. Crops Prod. 2021, 170, 113793. [Google Scholar] [CrossRef]
- Raal, A.; Arak, E.; Orav, A. The content and composition of the essential oil found in Carum carvi L. commercial fruits obtained from different countries. J. Essent. Oil Res. 2012, 24, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Sammama, H.; Kaoua, E.M.; Hsissou, D.; Latique, S.; Selmaoui, K.; Alfeddy, M.N. The impact of wheat and faba bean intercrop on the competitive interactions, grain yield, biochemical parameters and mineral content of leaves. Zemdirbyste 2021, 108, 233–240. [Google Scholar] [CrossRef]
- Andersen, M.K.; Hauggaard-Nielsen, H.; Ambus, P.; Jensen, E.S. Biomass production, symbiotic nitrogen fixation and inorganic N use in dual and tri-component annual intercrops. Plant Soil. 2005, 266, 273–287. [Google Scholar] [CrossRef] [Green Version]
- Hauggaard-Nielsen, H.; Andersen, M.K.; Joernsgaard, B.; Jensen, E.S. Density and relative frequency effects on competitive interactions and resource use in pea–barley intercrops. Field Crops Res. 2006, 95, 256–267. [Google Scholar] [CrossRef]
- Adamu, G.K.; Yusuf, M.A. A comparative study of changes in soil fertility under two farming practices in the Kano close-settled zone. Eur. Sci. J. 2014, 10, 313–323. [Google Scholar]
- Nasar, J.; Alam, A.; Nasar, A.; Khan, M.Z. Intercropping induce changes in above and below ground plant compartments in mixed cropping system. Biomed. J. Sci. Tech. Res. 2019, 17, 13043–13050. [Google Scholar] [CrossRef]
- Adamu, G.K.; Dawaki, M.U. Fertility Status of Soils under Irrigation along the Jakara Stream in Metropolitan Kano. Bayero. J. Pure Appl. Sci. 2008, 1, 67–70. [Google Scholar]
- Eichler-Löbermann, B.; Busch, S.; Jablonowski, N.D.; Kavka, M.; Brandt, C. Mixed cropping as affected by phosphorus and water supply. Agronomy 2020, 10, 1506. [Google Scholar] [CrossRef]
- Muofhe, M.L.; Dakora, F.D. Modification of rhizosphere pH by the symbiotic legume Aspalathus linearis growing in a sandy acidic soil. Funct. Plant Biol. 2000, 27, 1169–1173. [Google Scholar] [CrossRef]
- Li, Z.; Gao, Q.; Liu, Y.; He, C.; Zhang, X.; Zhang, J. Overexpression of transcription factor ZmPTF1 improves low phosphate tolerance of maize by regulating carbon metabolism and root growth. Planta 2011, 233, 1129–1143. [Google Scholar] [CrossRef]
- Li, L.; Tilman, D.; Lambers, H.; Zhang, F.S. Plant diversity and overyielding: Insights from belowground facilitation of intercropping in agriculture. New Phytol. 2014, 203, 63–69. [Google Scholar] [CrossRef]
- Nasar, J.; Shao, Z.; Gao, Q.; Zhou, X.; Fahad, S.; Liu, S.; Li, C.; Banda, J.S.K.; Kgorutla, L.E.; Dawar, K.M. Maize-alfalfa intercropping induced changes in plant and soil nutrient status under nitrogen application. Arch. Agron. Soil Sci. 2020, 68, 1–15. [Google Scholar] [CrossRef]
- Li, L.; Li, S.M.; Sun, J.H.; Zhou, L.L.; Bao, X.G.; Zhang, H.G.; Zhang, F.S. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc. Natl. Acad. Sci. USA 2007, 104, 11192–11196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Cheng, Z.; Meng, H.; Liu, L.; Li, H.; Dong, Y. Intercropping of green garlic (Allium sativum L.) induces nutrient concentration changes in the soil and plants in continuously cropped cucumber (Cucumis sativus L.) in a plastic tunnel. PLoS ONE 2013, 8, e62173. [Google Scholar] [CrossRef] [Green Version]
- Dotaniya, M.L.; Meena, V.D.; Basak, B.B.; Meena, R.S. Potassium uptake by crops as well as microorganisms. In Potassium for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 267–280. [Google Scholar]
- Pappa, V.A.; Rees, R.M.; Walker, R.L.; Baddeley, J.A.; Watson, C.A. Legumes intercropped with spring barley contribute to increased biomass production and carry-over effects. J. Agric. Sci. 2012, 150, 584–594. [Google Scholar] [CrossRef]
- Dusa, E.M.; Stan, V. The effect of intercropping on crop productivity and yield quality of oat (Avena sativa L.)/grain leguminous species (pea–Pissum sativum L., lentil–Lens culinaris L.) cultivated in pure stand and mixtures, in the organic agriculture system. Eur. Sci. J. 2013, 9, 69–78. [Google Scholar]
- Bellostas, N.M.; Hauggaard-Nielsen, H.; Andersen, M.K.; Jensen, E.S. Early interference dynamics in intercrops of pea, barley and oilseed rape. Biol. Agria Hortic. 2004, 21, 337–348. [Google Scholar] [CrossRef]
- Cui, T.; Fang, L.; Wang, M.; Jiang, M.; Shen, G. Intercropping of gramineous pasture ryegrass (Lolium perenne L.) and leguminous forage alfalfa (Medicago sativa L.) increases the resistance of plants to heavy metals. J. Chem. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Prakash, V.; Kundu, S.; Kumar, N.; Mina, B.L. Soil enzymatic activity as affected by long term application of farm yard manure and mineral fertilizer under a rainfed soybean–wheat system in NW Himalaya. Eur. J. Soil Biol. 2008, 44, 309–315. [Google Scholar] [CrossRef]
- Hazra, K.K.; Nath, C.P.; Singh, U.; Praharaj, C.S.; Kumar, N.; Singh, S.S.; Singh, N.P. Diversification of maize-wheat cropping system with legumes and integrated nutrient management increases soil aggregation and carbon sequestration. Geoderma 2019, 353, 308–319. [Google Scholar] [CrossRef]
- Alkorta, I.; Aizpurua, A.; Riga, P.; Albizu, I.; Amézaga, I.; Garbisu, C. Soil enzyme activities as biological indicators of soil health. Rev. Environ. Health 2003, 18, 65–73. [Google Scholar] [CrossRef]
- Liu, E.; Teclemariam, S.G.; Yan, C.; Yu, J.; Gu, R.; Liu, S.; He, W.; Liu, Q. Long-term effects of no-tillage management practice on soil organic carbon and its fractions in the northern China. Geoderma 2014, 213, 379–384. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Update 2015. World Soil Resources Report 106; FAO: Rome, Italy, 2015; p. 188. [Google Scholar]
- Staugaitis, G.; Vaišvila, Z.J. Soil Agrochemical Research. In Scientific-Methodical Material; The Lithuanian Research Centre for Agriculture and Forestry Agrochemical Research Laboratory: Kėdainiai District, Lithuanian, 2019; p. 112. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- McCormack, M.L.; Guo, D.; Iversen, C.M.; Chen, W.; Eissenstat, D.M.; Fernandez, C.W.; Li, L.; Ma, C.; Ma, Z.; Poorter, H.; et al. Building a better foundation: Improving root-trait measurements to understand and model plant and ecosystem processes. New Phytol. 2017, 215, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chunderova, A.I. The Enzymatic Activity of Sod-Podzolic Soils of the North-Western Region. Doctoral Dissertation, Tallinn University, Tallinn, Estonia, 1973; p. 46. (In Russian). [Google Scholar]
- Raudonius, S. Application of statistics in plant and crop research: Important issues. Zemdirb. Agric. 2017, 104, 377–382. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Multi-Cropping System Crops | Total Nitrogen, mg kg−1 | ||
|---|---|---|---|
| First Year of Caraway Vegetative Season 2017 | Second Year of Caraway Vegetative Season 2018 | Third Year of Caraway Vegetative Season 2019 | |
| Sole | |||
| Spring barley (SB-SB-SB) | 0.114 ± 0.005 bcd | 0.116 ± 0.012 b | 0.108 ± 0.004 c |
| Spring wheat (SW-SB-SB) | 0.111 ± 0.008 cd | 0.115 ± 0.009 b | 0.110 ± 0.005 c |
| Pea (P-SB-SB) | 0.109 ± 0.003 d | 0.131 ± 0.004 a | 0.128 ± 0.004 ab |
| Caraway (CA-BF) | 0.123 ± 0.005 abcd | 0.117 ± 0.003 b | 0.109 ± 0.003 c |
| Binary | |||
| S. barley + Caraway (SB-CA) | 0.132 ± 0.006 a | 0.132 ± 0.002 a | 0.129 ± 0.008 a |
| S. wheat + Caraway (SW-CA) | 0.124 ± 0.011 abc | 0.127 ± 0.011 ab | 0.126 ± 0.006 ab |
| Pea + Caraway (P-CA) | 0.128 ± 0.010 ab | 0.130 ± 0.018 a | 0.120 ± 0.005 b |
| Trinary | |||
| S. barley + Caraway + W. clover (SB-CA-WC) | 0.123 ± 0.012 abcd | 0.130 ± 0.012 a | 0.131 ± 0.005 a |
| S. wheat + Caraway + W. clover (SW-CA-WC) | 0.122 ± 0.013 abcd | 0.130 ± 0.010 a | 0.130 ± 0.006 a |
| Pea + Caraway + W. clover (P-CA-WC) | 0.130 ± 0.009 a | 0.133 ± 0.011 a | 0.132 ± 0.002 a |
| Multi-Cropping System Crops | Organic Carbon, mg kg−1 | ||
|---|---|---|---|
| First Year of Caraway Vegetative Season 2017 | Second Year of Caraway Vegetative Season 2018 | Third Year of Caraway Vegetative Season 2019 | |
| Sole | |||
| Spring barley (SB-SB-SB) | 0.92 ± 0.03 cd | 1.00 ± 0.05 bc | 1.40 ± 0.09 b |
| Spring wheat (SW-SB-SB) | 0.90 ± 0.06 d | 0.98 ± 0.04 c | 1.34 ± 0.07 b |
| Pea (P-SB-SB) | 0.89 ± 0.03 d | 1.06 ± 0.14 abc | 1.54 ± 0.07 a |
| Caraway (CA-BF) | 1.05 ± 0.02 a | 1.03 ± 0.02 abc | 1.30 ± 0.04 b |
| Binary | |||
| S. barley + Caraway (SB-CA) | 1.07 ± 0.04 a | 1.16 ± 0.04 a | 1.58 ± 0.04 a |
| S. wheat + Caraway (SW-CA) | 1.05 ± 0.02 a | 1.13 ± 0.08 a | 1.56 ± 0.12 a |
| Pea + Caraway (P-CA) | 0.97 ± 0.05 bc | 1.10 ± 0.06 ab | 1.54 ± 0.03 a |
| Trinary | |||
| S. barley + Caraway + W. clover (SB-CA-WC) | 0.96 ± 0.04 bc | 1.06 ± 0.03 abc | 1.59 ± 0.11 a |
| S. wheat + Caraway + W. clover (SW-CA-WC) | 1.06 ± 0.05 a | 1.11 ± 0.08 ab | 1.55 ± 0.11 a |
| Pea + Caraway + W. clover (P-CA-WC) | 1.02 ± 0.04 ab | 1.10 ± 0.03 b | 1.60 ± 0.06 a |
| Multi-Cropping System Crops | Available Phosphorus, mg kg−1 | ||
|---|---|---|---|
| First Year of Caraway Vegetative Season 2017 | Second Year of Caraway Vegetative Season 2018 | Third Year of Caraway Vegetative Season 2019 | |
| Sole | |||
| Spring barley (SB-SB-SB) | 229 ± 37.5 e | 193 ± 17.6 c | 254 ± 17.9 d |
| Spring wheat (SW-SB-SB) | 257 ± 6.08 c | 221 ± 26.1 bc | 226 ± 28.6 e |
| Pea (P-SB-SB) | 229 ± 33.4 e | 254 ± 25.3 abc | 252 ± 27.9 de |
| Caraway (CA-BF) | 260 ± 26.7 cd | 251 ± 44.0 abc | 258 ± 43.8 de |
| Binary | |||
| S. barley + Caraway (SB-CA) | 309 ± 14.8 a | 311 ± 16.3 a | 338 ± 41.0 a |
| S. wheat + Caraway (SW-CA) | 278 ± 18.5 bc | 282 ± 26.5 ab | 294 ± 28.3 bc |
| Pea + Caraway (P-CA) | 235 ± 27.0 de | 247 ± 29.3 abc | 260 ± 14.3 cd |
| Trinary | |||
| S. barley + Caraway + W. clover (SB-CA-WC) | 291 ± 12.2 ab | 279 ± 23.9 ab | 304 ± 26.4 b |
| S. wheat + Caraway + W. clover (SW-CA-WC) | 282 ± 13.9 bc | 269 ± 19.1 ab | 282 ± 36.0 bc |
| Pea + Caraway + W. clover (P-CA-WC) | 230 ± 27.1 e | 222 ± 28.5 abc | 244 ± 33.0 e |
| Multi-Cropping System Crops | Available Potassium, mg kg−1 | ||
|---|---|---|---|
| First Year of Caraway Vegetative Season 2017 | Second Year of Caraway Vegetative Season 2018 | Third Year of Caraway Vegetative Season 2019 | |
| Sole | |||
| Spring barley (SB-SB-SB) | 121 ± 16.3 bc | 124 ± 8.83 a | 145 ± 4.24 c |
| Spring wheat (SW-SB-SB) | 120 ± 10.0 bc | 119 ± 23.3 a | 140 ± 18.1 c |
| Pea (P-SB-SB) | 120 ± 20.9 bc | 130 ± 5.56 a | 149 ± 2.87 bc |
| Caraway (CA-BF) | 126 ± 5.32 abc | 118 ± 26.1 a | 143 ± 24.5 a |
| Binary | |||
| S. barley + Caraway (SB-CA) | 135 ± 5.12 ab | 132 ± 17.7 a | 212 ± 24.6 a |
| S. wheat + Caraway (SW-CA) | 132 ± 12.6 abc | 125 ± 13.8 a | 186 ± 25.8 a |
| Pea + Caraway (P-CA) | 120 ± 7.12 bc | 120 ± 18.1 a | 194 ± 19.8 a |
| Trinary | |||
| S. barley + Caraway + W. clover (SB-CA-WC) | 139 ± 2.75 a | 135 ± 26.6 a | 236 ± 39.2 a |
| S. wheat + Caraway + W. clover (SW-CA-WC) | 118 ± 16.9 c | 126 ± 34.9 a | 197 ± 54.9 a |
| Pea + Caraway + W. clover (P-CA-WC) | 117 ± 9.78 c | 131 ± 8.62 a | 203 ± 45.0 a |
| Name of Pesticide | Type | Active Substance | Amount | Abbreviation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fenix | herbicide | aclonifen 600 g L−1 | 3.00 L ha−1 | F | ||||||
| Signum | fungicide | boscalid 267 g kg−1 + pyraclostrobin 67 g kg−1 | 0.50 L ha−1 | S | ||||||
| Cyperkill 500 EC | insecticide | cypermethrin 500 g L−1 | 0.05 L ha−1 | C | ||||||
| Elegant 2 FD | herbicide | florasulam 6.25 g L−1 + 2.4-D 300 g L−1 | 0.40 L ha−1 | E | ||||||
| Karate Zeon 5 CS | insecticide | lambda-cyhalothrin 50 g L−1 | 0.14 L ha−1 | KZ | ||||||
| Bumper 25 EC | fungicide | propiconazole 250 g L−1 | 0.50 L ha−1 | B | ||||||
| Bulldock 025 EC | insecticide | beta-cyfluthrin 25 g L−1 | 0.30 L ha−1 | Bu | ||||||
| Miradol 250 SC | fungicide | azoxystrobin 250 g L−1 | 0.60 L ha−1 | M | ||||||
| Trimmer | herbicide | tribenuron-methyl 500 g kg−1 | 0.10 kg ha−1 | T | ||||||
| 2017 | 2018 | 2019 | ||||||||
| T1 | T2 | T3 | T1 | T2 | T3 | T1 | T2 | T3 | ||
| Sole | SB-SB-SB | – | E + KZ ** | B ** | – | E + KZ ** | M + Bu ** | – | E + T ** | B ** |
| SW-SB-SB | – | E + KZ ** | B ** | – | E + KZ ** | M + Bu ** | – | E + T ** | B ** | |
| P-SB-SB | F * | – | S + C ** | – | E + KZ ** | M + Bu ** | – | – | B ** | |
| CA-BF | F * | – | – | – | – | – | – | – | ||
| Binary | SB-CA | – | – | B ** | – | – | – | – | – | – |
| SW-CA | – | – | B ** | – | – | – | – | – | – | |
| P-CA | F * | – | S + C ** | – | – | – | – | – | – | |
| Trinary | SB-CA-WC | – | – | B ** | – | – | – | – | – | – |
| SW-CA-WC | – | – | B ** | – | – | – | – | – | – | |
| P-CA-WC | – | – | S + C ** | – | – | – | – | – | – | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudinskienė, A.; Marcinkevičienė, A.; Velička, R.; Kosteckas, R.; Kriaučiūnienė, Z.; Vaisvalavičius, R. The Comparison of Soil Agrochemical and Biological Properties in the Multi-Cropping Farming Systems. Plants 2022, 11, 774. https://doi.org/10.3390/plants11060774
Rudinskienė A, Marcinkevičienė A, Velička R, Kosteckas R, Kriaučiūnienė Z, Vaisvalavičius R. The Comparison of Soil Agrochemical and Biological Properties in the Multi-Cropping Farming Systems. Plants. 2022; 11(6):774. https://doi.org/10.3390/plants11060774
Chicago/Turabian StyleRudinskienė, Aušra, Aušra Marcinkevičienė, Rimantas Velička, Robertas Kosteckas, Zita Kriaučiūnienė, and Rimantas Vaisvalavičius. 2022. "The Comparison of Soil Agrochemical and Biological Properties in the Multi-Cropping Farming Systems" Plants 11, no. 6: 774. https://doi.org/10.3390/plants11060774