
Salinity Modulates Juncus acutus L. Tolerance to Diesel Fuel Pollution

 , ,
, ,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
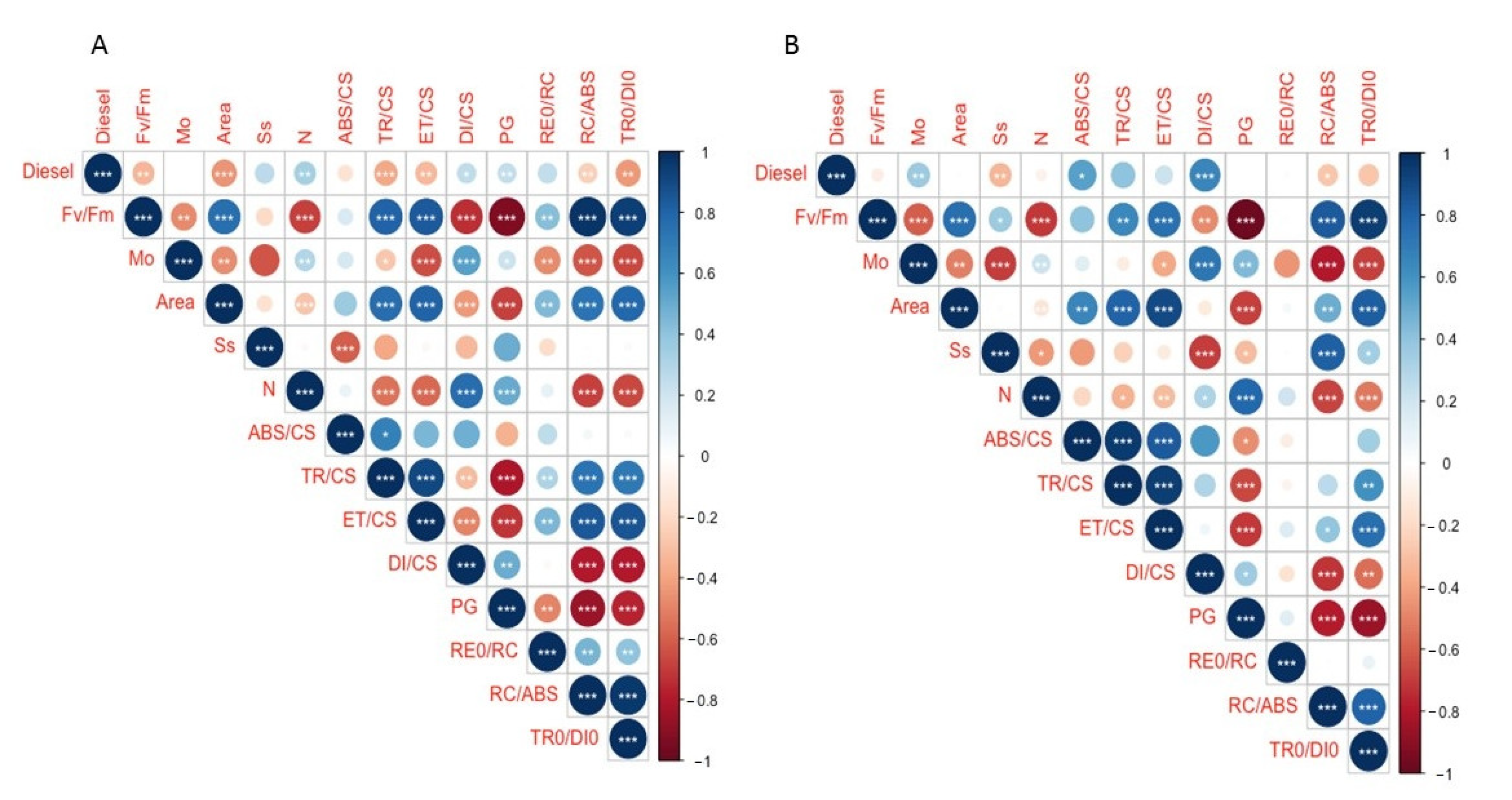
2. Results
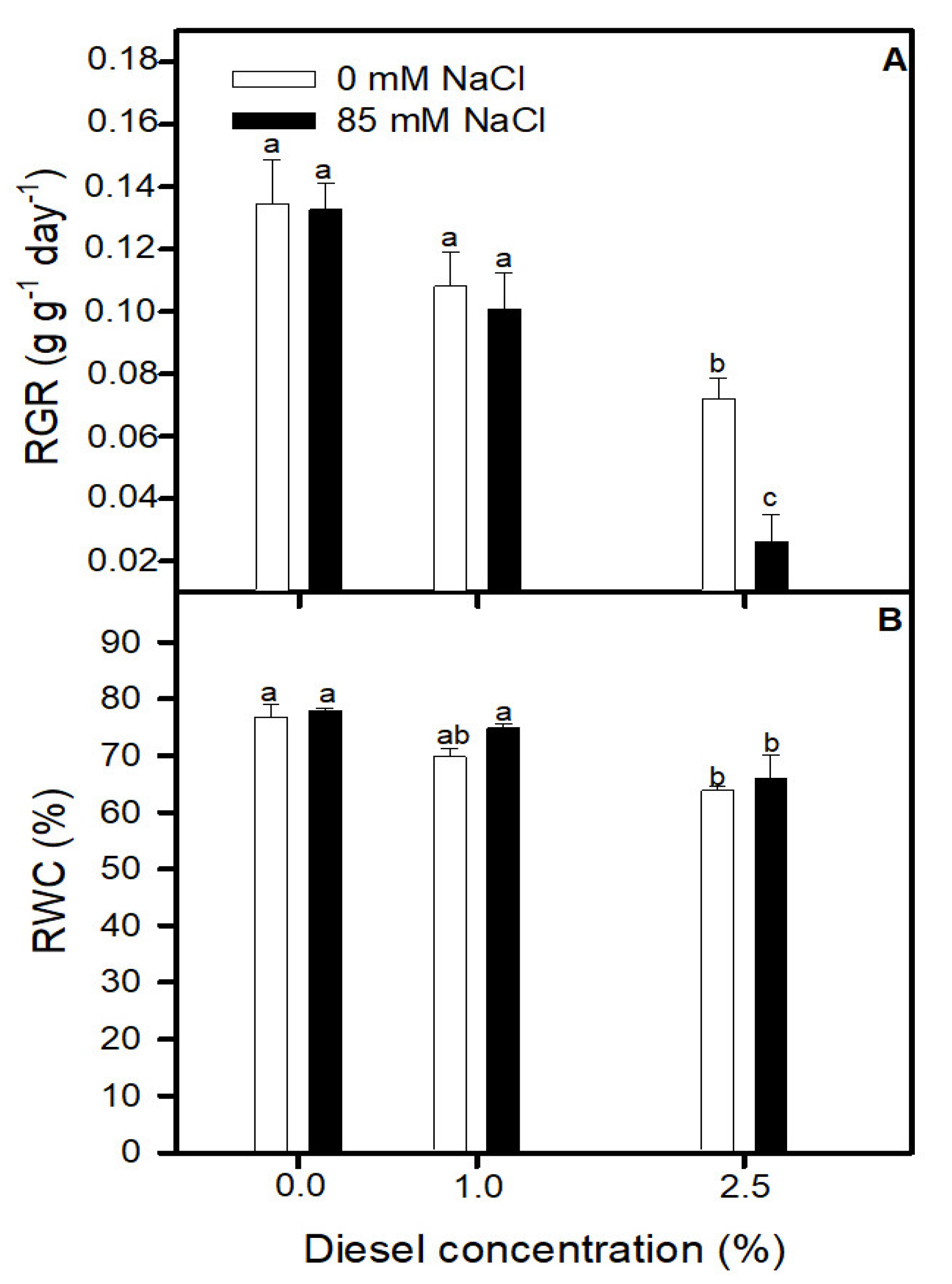
2.1. Effects of Diesel and NaCl on Relative Growth Rate and Relative Water Content
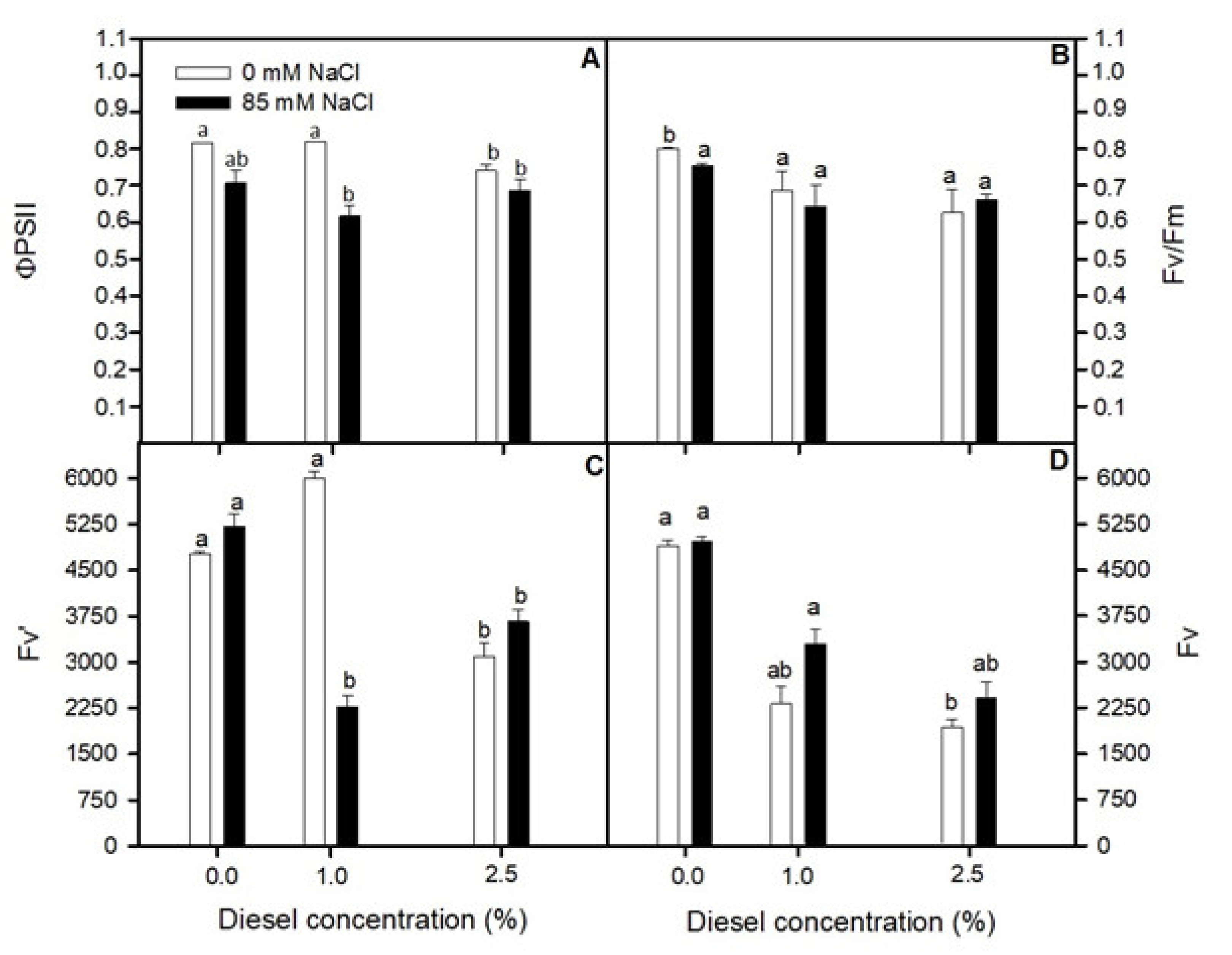
2.2. Effects of Diesel and NaCl on Chlorophyll a Fluorescence
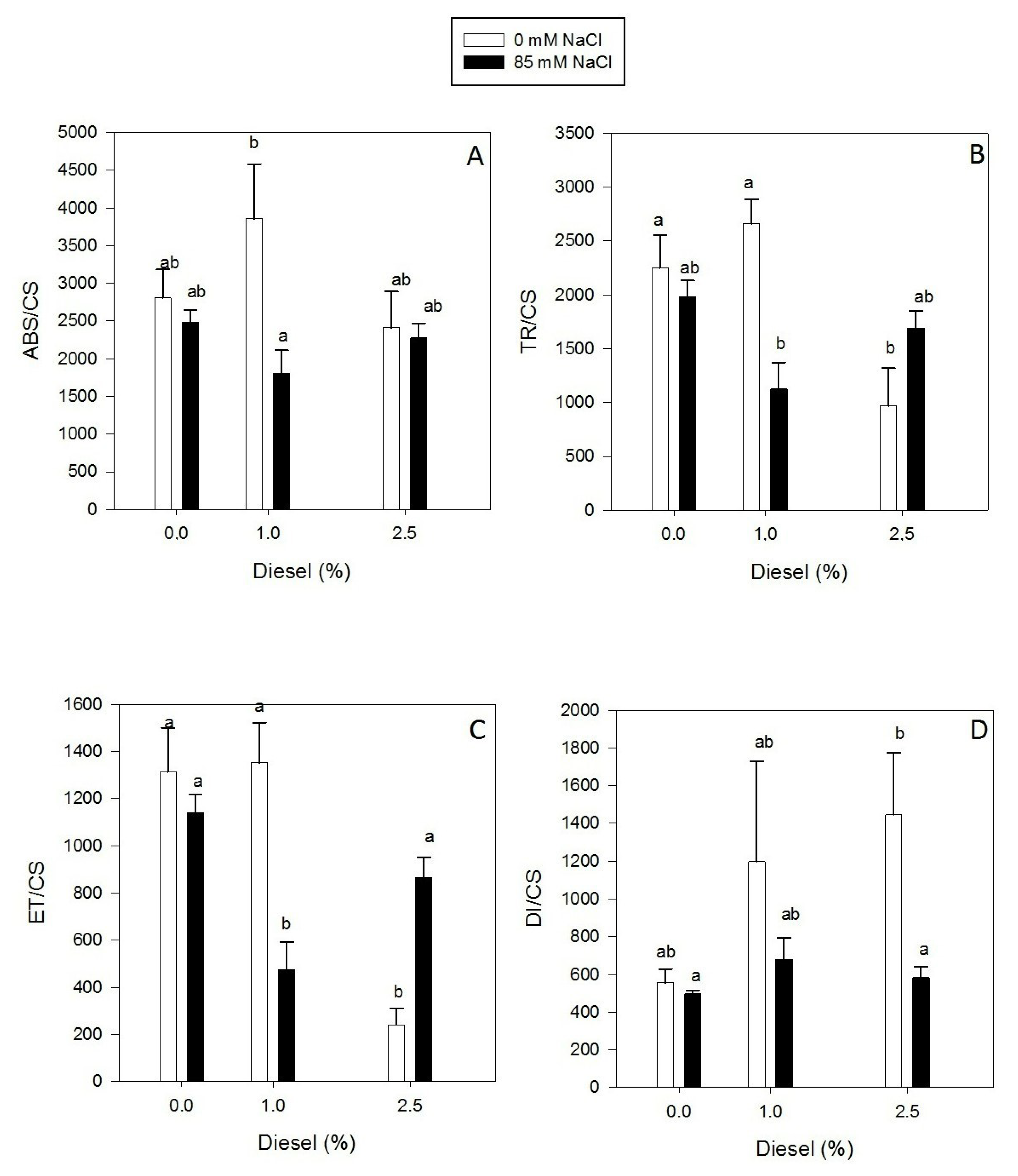
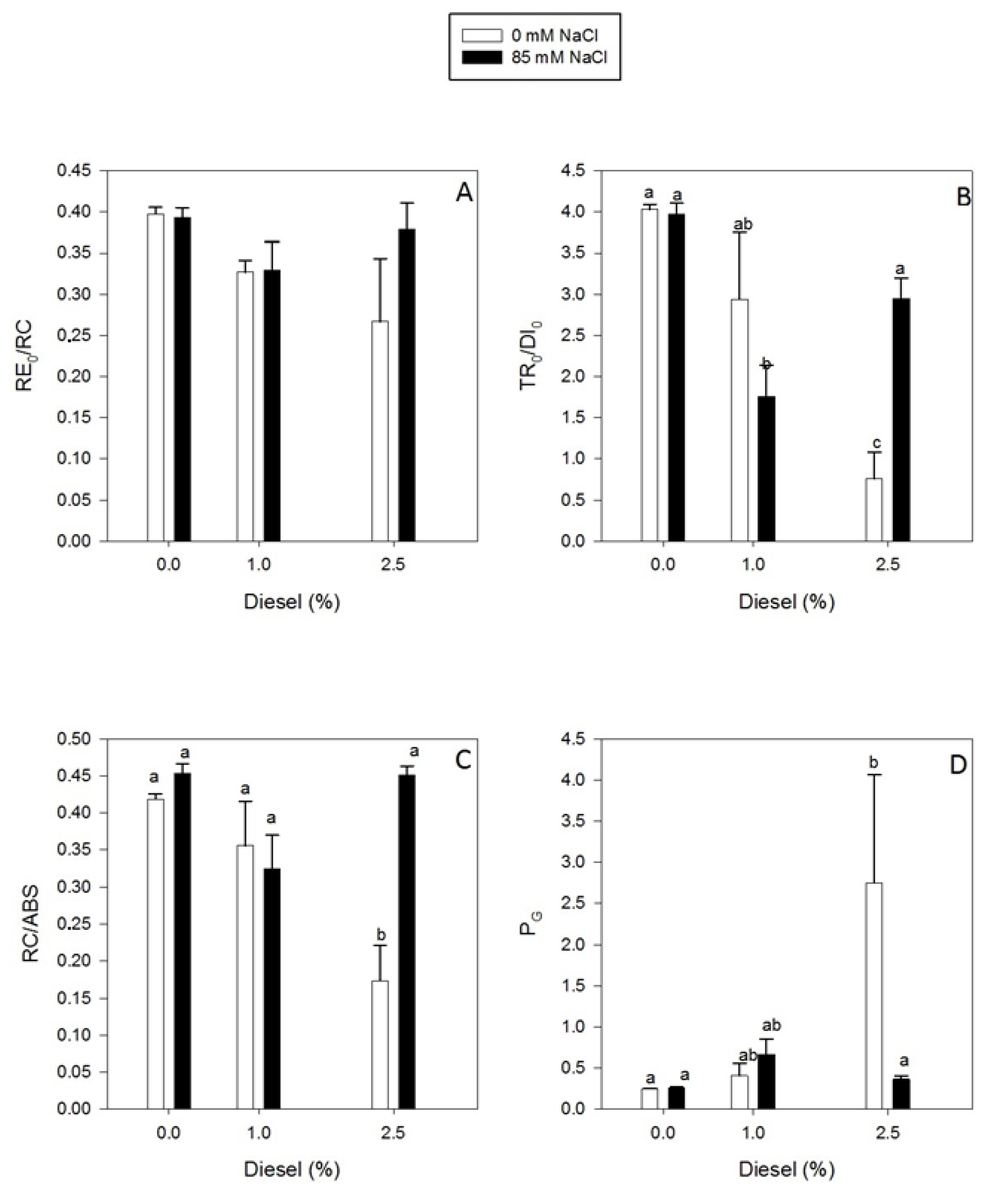
2.3. Effects of Diesel and NaCl on Kautsky Curves Parameters
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. Diesel and NaCl Experimental Stress Treatments
4.3. Growth Measurements and Water Content
4.4. Chlorophyll Fluorescence Measurement
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rusin, M.; Gospodarek, J.; Nadgórska-Socha, A. Soil Pollution by Petroleum-Derived Substances and its Bioremediation: The Effect on Aphis fabae Scop. Infestation and Antioxidant Response in Vicia faba L. Agronomy 2020, 10, 147. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, S.; Adams, W.A. The fate of diesel hydrocarbons in soils and their effect on the germination of perennial ryegrass. Environ. Toxicol. Int. J. 2002, 17, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Dib, D.; Sadoudi Ali Ahmed, D. Influence of diesel fuel contamination on Xanthium strumarium L. germination and growth. Int. J. Phytoremediation 2020, 22, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Gómez, S.; Petenello, M.C.; Feldman, S.R. Growth, nutrient status, and photosynthetic response to diesel-contaminated soil of a cordgrass, Spartina argentinensis. Mar. Pollut. Bull. 2014, 79, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Hawrot-Paw, M.; Wijatkowski, A.; Mikiciuk, M. Influence of diesel and biodiesel fuel-contaminated soil on microorganisms, growth and development of plants. Plant Soil Environ. 2015, 61, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Bona, C.; Rezende, I.M.D.; Santos, G.D.O.; Souza, L.A.D. Effect of soil contaminated by diesel oil on the germination of seeds and the growth of Schinus terebinthifolius Raddi (Anacardiaceae) seedlings. Braz. Arch. Biol. Technol. 2011, 54, 1379–1387. [Google Scholar] [CrossRef] [Green Version]
- Cruz, J.M.; Tamada, I.S.; Lopes, P.R.M.; Montagnolli, R.N.; Bidoia, E.D. Biodegradation and phytotoxicity of biodiesel, diesel, and petroleum in soil. Water Air Soil Pollut. 2014, 225, 1962. [Google Scholar] [CrossRef]
- Ogbo, E.M. Effects of diesel fuel contamination on seed germination of four crop plants-Arachis hypogaea, Vigna unguiculata, Sorghum bicolor and Zea mays. Afr. J. Biotechnol. 2009, 8, 250–253. [Google Scholar]
- Luhach, J.; Chaudhry, S. Effect of Diesel Fuel Contamination on Seed Germination and Growth of Four Agricultural Crops. Univers. J. Environ. Res. Technol. 2012, 2, 311–317. [Google Scholar]
- Adam, G.; Duncan, H. Influence of diesel fuel on seed germination. Environ. Pollut. 2020, 120, 363–370. [Google Scholar] [CrossRef]
- Truskewycz, A.; Gundry, T.D.; Khudur, L.S.; Kolobaric, A.; Taha, M.; Aburto-Medina, A.; Shahsavari, E. Petroleum hydrocarbon contamination in terrestrial ecosystems—Fate and microbial responses. Molecules 2019, 24, 3400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyszkowski, M.; Sivitskaya, V. Effect of sorbents on the content of trace elements in maize cultivated on soil contaminated with heating oil. Int. Agrophys. 2019, 33, 437–444. [Google Scholar] [CrossRef]
- Pérez-Romero, J.A.; Redondo-Gómez, S.; Mateos-Naranjo, E. Growth and photosynthetic limitation analysis of the Cd-accumulator Salicornia ramosissima under excessive cadmium concentrations and optimum salinity conditions. Plant Physiol. Biochem. 2016, 109, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Naranjo, E.; Pérez-Romero, J.A.; Mesa-Marín, J.; López-Jurado, J.; Redondo-Gómez, S. Inter-population differences tolerance to Cu excess during the initials phases of Juncus acutus life cycle: Implications for the design of metal restoration strategies. Int. J. Phytoremediation 2019, 21, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, S.D.; Shann, J.R.; Crowley, D.E.; Anderson, T.A. Phytoremediation of Contaminated Water and Soil; ACS Publications: Washington, DC, USA, 1997; pp. 2–17. [Google Scholar]
- Palmroth, M.R.; Pichtel, J.; Puhakka, J.A. Phytoremediation of subarctic soil contaminated with diesel fuel. Bioresour. Technol. 2002, 84, 221–228. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Andrades-Moreno, L.; Parra, R.; Valera-Burgos, J.; Real, M.; Mateos-Naranjo, E.; Cornejo, J. Spartina densiflora demonstrates high tolerance to phenanthrene in soil and reduces it concentration. Mar. Pollut. Bull. 2011, 62, 1800–1808. [Google Scholar] [CrossRef] [PubMed]
- Adam, G.; Duncan, H.J. Effect of diesel fuel on growth of selected plant species. Environ. Geochem. Health 1999, 21, 353–357. [Google Scholar] [CrossRef] [Green Version]
- Pawluśkiewicz, B.; Gnatowski, T.; Janicka, M. The Influence of Soil Contamination with Diesel Oil on Germination Dynamics and Seedling Development of Selected Species of the Fabaceae family. J. Ecol. Eng. 2020, 21, 210–218. [Google Scholar] [CrossRef]
- Sparks, E.L.; Cebrian, J.; Biber, P.D.; Sheehan, K.L.; Tobias, C.R. Cost-effectiveness of two small-scale salt marsh restoration designs. Ecol. Eng. 2013, 53, 250–256. [Google Scholar] [CrossRef]
- Marques, B.; Lillebø, A.I.; Pereira, E.; Duarte, A.C. Mercury cycling and sequestration in salt marshes sediments: An ecosystem service provided by Juncus maritimus and Scirpus maritimus. Environ. Pollut. 2011, 159, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Naranjo, E.; Castellanos, E.M.; Perez-Martin, A. Zinc tolerance and accumulation in the halophytic species Juncus acutus. Environ. Exp. Bot. 2014, 100, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Santos, D.; Duarte, B.; Caçador, I. Unveiling Zn hyperaccumulation in Juncus acutus: Implications on the electronic energy fluxes and on oxidative stress with emphasis on non-functional Zn-chlorophylls. J. Photochem. Photobiol. B Biol. 2014, 140, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Christofilopoulos, S.; Syranidou, E.; Gkavrou, G.; Manousaki, E.; Kalogerakis, N. The role of halophyte Juncus acutus L. in the remediation of mixed contamination in a hydroponic greenhouse experiment. J. Chem. Technol. Biotechnol. 2016, 91, 1665–1674. [Google Scholar] [CrossRef]
- Lin, Q.; Mendelssohn, I.A. Potential of restoration and phytoremediation with Juncus roemerianus for diesel-contaminated coastal wetlands. Ecol. Eng. 2009, 35, 85–91. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Pérez-Romero, J.A.; Redondo-Gómez, S.; Mesa-Marín, J.; Castellanos, E.M.; Davy, A.J. Salinity alleviates zinc toxicity in the saltmarsh zinc-accumulator Juncus acutus. Ecotoxicol. Environ. Saf. 2018, 163, 478–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gbadebo, A.M.; Adenuga, M.D. Effect of crude oil on the emergence and growth of cowpea in two contrasting soil types from Abeokuta, Southwestern Nigeria. Asian J. Appl. Sci. 2012, 5, 232–239. [Google Scholar]
- Rusin, M.; Gospodarek, J.; Nadgórska-Socha, A. The effect of petroleum-derived substances on the growth and chemical composition of Vicia faba L. Pol. J. Environ. Stud. 2015, 24, 2157–2166. [Google Scholar] [CrossRef]
- Al Hassan, M.; López-Gresa, M.; Boscaiu, M.; Vicente, O. Stress tolerance mechanisms in Juncus: Responses to salinity and drought in three Juncus species adapted to different natural environments. Funct. Plant Biol. 2016, 43, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Castro-Mancilla, Y.V.; de la Rosa-Manzano, E.; Castro-Nava, S.; Martínez-Avalos, J.G. Physiological responses of Quercus oleoides (Schltdl & Cham) to soils contaminated by diesel. Plant Prod. Sci. 2019, 22, 519–529. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Duarte, B.; Cabrita, M.T.; Gameiro, C.; Matos, A.R.; Godinho, R.; Marques, J.C.; Caçador, I. Disentangling the photochemical salinity tolerance in Aster tripolium L.: Connecting biophysical traits with changes in fatty acid composition. Plant Biol. 2017, 19, 239–248. [Google Scholar] [CrossRef]
- Duarte, B.; Durante, L.; Marques, J.C.; Reis-Santos, P.; Fonseca, V.F.; Caçador, I. Development of a toxicophenomic index for trace element ecotoxicity tests using the halophyte Juncus acutus: Juncus-TOX. Ecol. Indic. 2021, 121, 107097. [Google Scholar] [CrossRef]
- Martina, G.; Irene, R.; Paolo, A.; Gianniantonio, P.; Beatrice, P.; Grifoni, M. A Preliminary Study on Lupinus albus and Raphanus sativus Grown in Soil Affected by Oil Spillage. Bull. Environ. Contam. Toxicol. 2021, 107, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Erickson, T.E.; Ball, A.S.; Ryan, M.H. A review of germination and early growth as a proxy for plant fitness under petrogenic contamination—knowledge gaps and recommendations. Sci. Total Environ. 2017, 603, 728–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassalle, G.; Fabre, S.; Credoz, A.; Dubucq, D.; Elger, A. Monitoring oil contamination in vegetated areas with optical remote sensing: A comprehensive review. J. Hazard. Mater. 2020, 393, 122427. [Google Scholar] [CrossRef] [PubMed]
- Pullagurala, V.L.R.; Rawat, S.; Adisa, I.O.; Hernandez-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Plant uptake and translocation of contaminants of emerging concern in soil. Sci. Total Environ. 2018, 636, 1585–1596. [Google Scholar] [CrossRef] [PubMed]
- Belkheiri, O.; Mulas, M. The effects of salt stress on growth, water relations and ion accumulation in two halophyte Atriplex species. Environ. Exp. Bot. 2013, 86, 17–28. [Google Scholar] [CrossRef]
- Pérez-Romero, J.A.; Idaszkin, Y.L.; Barcia-Piedras, J.M.; Duarte, B.; Redondo-Gómez, S.; Caçador, I.; Mateos-Naranjo, E. Disentangling the effect of atmospheric CO2 enrichment on the halophyte Salicornia ramosissima J. Woods physiological performance under optimal and suboptimal saline conditions. Plant Physiol. Biochem. 2018, 127, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.; Arnon, D.I. The water culture method for growing plants without soil. Calif. Agric. Exp. Stn. Bull. 1938, 347, 1–39. [Google Scholar]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Duarte, B.; Santos, D.; Marques, J.C.; Caçador, I. Ecophysiological constraints of two invasive plant species under a saline gradient: Halophytes versus glycophytes. Estuar. Coast. Shelf Sci. 2015, 167, 154–165. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer. Primer-e Plymouth 2006, 866, 154–165. [Google Scholar]
- Duarte, B.; Matos, A.R.; Caçador, I. Photobiological and lipidic responses reveal the drought tolerance of Aster tripolium cultivated under severe and moderate drought: Perspectives for arid agriculture in the mediterranean. Plant Physiol. Biochem. 2020, 154, 304–315. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Romero, J.A.; Barcia-Piedras, J.-M.; Redondo-Gómez, S.; Caçador, I.; Duarte, B.; Mateos-Naranjo, E. Salinity Modulates Juncus acutus L. Tolerance to Diesel Fuel Pollution. Plants 2022, 11, 758. https://doi.org/10.3390/plants11060758
Pérez-Romero JA, Barcia-Piedras J-M, Redondo-Gómez S, Caçador I, Duarte B, Mateos-Naranjo E. Salinity Modulates Juncus acutus L. Tolerance to Diesel Fuel Pollution. Plants. 2022; 11(6):758. https://doi.org/10.3390/plants11060758
Chicago/Turabian StylePérez-Romero, Jesús Alberto, José-María Barcia-Piedras, Susana Redondo-Gómez, Isabel Caçador, Bernardo Duarte, and Enrique Mateos-Naranjo. 2022. "Salinity Modulates Juncus acutus L. Tolerance to Diesel Fuel Pollution" Plants 11, no. 6: 758. https://doi.org/10.3390/plants11060758